Naraoiidae
Die Naraoiidae sind eine ausgestorbene, nur fossil erhaltene trilobitenähnliche Arthropodenfamilie, die hauptsächlich im unteren und mittleren Kambrium auftrat.
| Naraoiidae | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Naraoia spinosa | ||||||||||
| Zeitliches Auftreten | ||||||||||
| Kambrium (2. Serie) bis Silur (Pridolium) | ||||||||||
| 520 bis 410 Mio. Jahre | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Naraoiidae | ||||||||||
| Walcott, 1912 |
Beschreibung
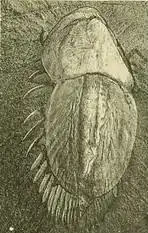
Die Naraoiidae umfassen Tiere, deren Körper im Leben von zwei großen, gelenkig miteinander verbundenen Platten (Tergiten) verdeckt war, so dass bei dorsaler Ansicht nur diese zu sehen waren. Sie ähneln dadurch in der Erscheinung aberranten Trilobiten, die nur aus Kopfschild und Schwanzschild ohne freie Segmente dazwischen aufgebaut wären. Der anteriore (vordere) oder Kopfschild war meist etwas breiter als der posteriore (hintere) oder Schwanzschild, dieser dafür merklich länger. Die im Leben gewölbten Schilder besaßen außen wenig differenzierte Strukturen, bei einigen Arten sitzt am Seitenrand eine seitlich gerichtete Dornenreihe, oder am Hinterende des anterioren Schilds sitzen seitlich der Gelenknaht zwei sog. Wangendornen. Im Unterschied zu den Trilobiten war allerdings die Dorsalhülle des Exoskeletts nicht durch Kalkeinlagerung verstärkt. Die Körpergliederung (Tagmosis) der Naraoiidae war auf der dorsalen (oberen) und der ventralen (unteren) Seite markant verschieden.
Bei zahlreichen fossilen Exemplaren ist auf dem Kopfschild eine zweiteilig zur Mittellinie symmetrische, baumförmig verzweigte Struktur erkennbar, die entweder ein Relief bildet oder durch taphonomische mineralische Ausfällungen anders, meist dunkler, gefärbt ist. Nach übereinstimmender Interpretation handelt es sich dabei um das Verdauungssystem der Tiere, das aus einem zentralen Darm besteht, von dem seitlich zahlreiche, mehrfach verzweigte, blind endende Divertikel oder Caeca abzweigten, wobei die vorderste Abzweigung die weitaus größte war und fast den ganzen anterioren Schild ausfüllte. Im Hinterende ist ein gerades, fast glattes Darmrohr mit nur wenigen, schwach ausgeprägten Seitenästen erkennbar. Nach der Anatomie und dem fossil erhaltenen Darminhalt gilt es als wahrscheinlich, dass die Naraoiidae Substratfresser waren, die nährstoffreichen, organisch angereicherten Schlamm fraßen und den organischen Anteil verdauten. Einige Forscher nehmen aber auch eine Lebensweise als Räuber oder Aasfresser an.
Während solche meist relativ merkmalsarmen Fossilien von Naraoiidae, die nur die zwei dorsalen Platten erkennen lassen, in entsprechenden Strata häufig und weit verbreitet sein können, liegen von deutlich weniger Exemplaren auch Befunde von der Körperunterseite (Ventralseite) vor. Hier zeigten die Tiere ein radikal anderes Erscheinungsbild. Den zwei Tergiten steht eine lange Reihe (bis zu 30) deutlich schmalere Sternite gegenüber. Dabei saßen die meisten, zwischen 15 und 26, unter dem posterioren Schild. Der Körper beginnt auf der Unterseite am Vorderende mit einem Hypostom genannten Kopflappen, der bei den Naraoiidae durch drei Vorwölbungen dreiteilig erscheint. Die seitlichen Vorwölbungen werden teilweise als Komplexaugen interpretiert.[1] Am Hypostom saßen lange, gegliederte Antennen an, die weit über den Kopfschild nach vorn vorstanden. Die Antennenportion unterhalb des Kopfschilds ist nur sehr selten erhalten, bei einem einzigen Exemplar von Misszhouia longicaudata ist erkennbar, dass der Basis der Antennengeißel eine winzige, dreigliedrige Nebengeißel ansaß; möglicherweise gehen die Antennen also auf Spaltbeine zurück. Bei den Segmenten hinter dem antennentragenden Abschnitt (dessen Segmentnatur interpretationsahängig und daher umstritten ist) entsprach jeweils ein Extremitätenpaar einem Sternit, ähnlich den rezenten Euarthropoden. Die Beine waren über die Körperlänge jeweils sehr ähnlich aufgebaut, bereits das erste Beinpaar entsprach dem Grundbauplan. Echte Mundwerkzeuge sind also nicht erkennbar, obwohl das vorderste Beinpaar seitlich des Hypostoms und der Mundöffnung saß. Der vordere Körperabschnitt mit dem Kopfschild trug immer, neben den Antennen, vier Extremitätenpaare.
Die Beingliederung der Naraoiidae ist an fast allen Arten erkennbar. Es handelt sich um typische Spaltbeine. Proximal zum Körper saß ein Basis genanntes, plattenartiges Glied. An dessen Innenseite war ein mit Dornen besetzter Endit (oder Gnathobasis) abgesetzt, die Endite beider Beinreihen umgriffen eine zentrale Nahrungsrinne. Es ist also anzunehmen, dass wie zum Beispiel bei den rezenten Kiemenfußkrebsen, alle Beine neben der Fortbewegung auch an der Nahrungsaufnahme beteiligt waren. An der Basis saß die eigentliche Extremität an. Der Innenast (Endopodit) des Spaltbeins bestand aus bei allen Arten sieben Gliedern und einer abgesetzten Kralle am Ende. Der Bau des Exopoditen differierte zwischen den Arten. Meist war er kammförmig gefiedert, wobei ein Basisschaft eine Reihe flacher, bandartiger Lamellen trug, am Ende der Reihe saß ein lappenartiger Anhang mit zahlreichen Borsten. Bei Naraoia spinosa war der Exopodit geschlossen zungenförmig, der nach außen hin breiter wurde. Das Bein gleicht in seinem Aufbau vollkommen demjenigen ursprünglicher Trilobiten wie Eoredlichia oder Olenoides, aber auch anderen Stammgruppen-Arthopoden, etwa Xandarella. Dass bei Arten einer Gattung sowohl kammartige wie zungenförmige Exopodite vorhanden waren, zeigt, dass das Merkmal für eine phylogenetische Analyse nur mit Vorsicht verwendbar ist.
Verbreitung
Naraoiidae sind im Fossilbericht aus dem Kambrium, Ordovizium und Silur gemeldet. Die meisten und am besten erhaltenen kambrischen Funde stammen aus dem Burgess-Schiefer Britisch Kolumbiens und der Chengjiang-Faunengemeinschaft aus dem Maotianshan-Schiefer der Volksrepublik China. Weitere Funde stammen aus der mittelkambrischen Marjum-Formation in Utah, USA, und Gibson Jack Formation im östlichen Idaho, USA. Die ordovizische Pseudonaraoia hammanni wurde in der mittelordovizischen Šárka Formation, am Roten Berg (Červený vrch) nahe der Stadt Prag gefunden. Der einzige silurische Vertreter, Naraoia bertiensis entstammt der obersilurischen (Pridolium) Bertie-Formation in Südost-Ontario.
Systematik und Phylogenie
Der Familie Naraoiidae werden acht Arten aus drei Gattungen zugeschrieben:
- Naraoia
- Naraoia compacta Walcott, 1912. Typusart der Gattung.
- Naraoia spinifer Walcott, 1931
- Naraoia halia Simonetta & Delle Cave, 1975 (die Art wird von manchen Taxonomen nur als Synonym von Naraoia compacta aufgefasst)
- Naraoia spinosa Zhang and Hou, 1985
- Naraoia bertiensis Caron et al., 2004[2]
- Naraoia taijiangensis Peng et al., 2012[3]
- Misszhouia Chen et al., 1997
- Misszhouia longicaudata (Zhang & Hou, 1985).
- Pseudonaraoia Budil et al., 2003
- Pseudonaraoia hammanni Budil et al., 2003[4]
Die Naraoiiden gehören in einen fossilen Formenkreis, der manche Merkmale mit den bekannteren Trilobiten gemeinsam hat, diesen gegenüber aber auch markante Unterschiede aufweist, am auffallendsten dabei die nicht verkalkte Dorsalhülle und das völlige Fehlen freier Rumpfsegmente. Sie werden traditionell mit zahlreichen weiteren fossilen Arthropoden ähnlicher Merkmalskombinationen, meist aus dem Kambrium, in einen weiten, schlecht definierten Verwandtschaftskreis gestellt, der als Lamellipedia, Trilobitomorpha oder auch Arachnomorpha bezeichnet wird. Bearbeiter der Vergangenheit ordneten die Naraoiidae sogar oft als Ordnung den Trilobiten selbst zu, dies zu einer Zeit, als die meisten verwandten Gruppen noch unbekannt waren. Ähnliche fossile Formen wurden in den letzten Jahren mehrfach neu gefunden, so zum Beispiel 2013 Arthroaspis bergstroemi aus der Sirius-Passet-Faunengemeinschaft Grönlands[5] oder 2010 die Gattungen Emucaris und Kangacaris aus dem unterkambrischen Emu Bay-Schiefer von Kangaroo Island, Süd-Australien.[6]
Neueren phylogenetischen Analysen auf kladistischer Basis zufolge ist die wahrscheinlichste Schwestergruppe der Naroiidae die Familie Liwiidae, eine artenarme und schlecht erforschte, erst 1988 neu beschriebene Gruppe, die den Naroiidae in den meisten Merkmalen stark ähneln, aber, anders als diese, freie Rumpfsegmente besitzen. Nächstverwandt zu dieser Klade wären die Emucarididae Australiens (inzwischen auch aus China angegeben). Diese Gruppen zusammen werden als Ordnung Nectaspida Raymond, 1920 aufgefasst (auch Nectaspia, Nectaspidida oder Nektaspida geschrieben). Ihre Zusammengehörigkeit gilt inzwischen als gut abgesichert.[7][6][5]
Während die Verwandtschaft bis hierhin noch recht plausibel ist, ist die weitere Einordnung noch recht spekulativ und variiert zwischen verschiedenen wissenschaftlichen Studien. Die Gruppe könnte mit einigen weiteren, darunter auch den Trilobiten, eine oft Artiopoda genannte Klade bilden, bzw., unter Einbeziehung auch von Marrella und Verwandten (Marrellomorpha), ein Taxon Lamellipedia. Diese Gruppe ist, vielen, aber nicht allen[8] Analysen zufolge, vermutlich näher mit der Stammgruppe der Mandibeltiere (Mandibulata) verwandt als zu derjenigen der Kieferklauenträger (Chelicerata). Die früher favorisierte Zusammenfassung als „Arachnomorpha“ erscheint danach heute nur noch eine Möglichkeit unter Vielen.
Quellen
- X.-L. Zhang, D.-G. Shu, D. H. Erwin (2007): Cambrian naraoiids (Arthropoda): morphology, ontogeny, systematics, and evolutionary relationships. Journal of Paleontology 81: 1–52. doi:10.1666/06-082.1
Einzelnachweise
- Brigitte Schoenemann, Euan N. K. Clarkson (2012): Compound Eyes in the Chengjiang Biota. Scientific Papers University of Latvia, Earth and Environmental Sciences 783: 57-71.
- Jean-Bernard Caron, David M. Rudkin, Stuart Milliken (2004): A New Late Silurian (Pridolian) Naraoiid (Euarthropoda: Nektaspida) from the Bertie Formation of Southern Ontario Canada: Delayed Fallout from the Cambrian Explosion. Journal of Paleontology 78 (6): 1138-1145.
- Jin Peng, Yuanlong Zhao, Hijing Sun (2012): Discovery and significance of Naraoia from the Qiandongian (lowe Cambrian) Balang Formation, Eastern Guizhou, South China. Bulletin of Geosciences 87(1):143-150.
- P. Budil, O. Fatka, J. Bruthansová (2003). Trilobite fauna of the Šárka Formation at Praha – Červený vrch Hill (Ordovician, Barrandian area, Czech Republic). Bulletin of Geosciences 78(2): 113–117.
- Martin Stein, Graham E Budd, John S Peel, David AT Harper (2013): Arthroaspis n. gen., a common element of the Sirius Passet Lagerstätte (Cambrian, North Greenland), sheds light on trilobite ancestry. BMC Evolutionary Biology 2013, 13:99. online
- J.R. Paterson, G.D. Edgcombe, D.C. Garcia-Bellido, J.B. Jago J.G. Gehling (2010): Nektaspid arthropods from the Lower Cambrian Emu Bay Shale Lagerstätte, South Australia, with a reassessment of lamellipedian relationships. Palaeontology 53: 377–402. doi:10.1111/j.1475-4983.2010.00932.x
- Martin Stein & Paul A. Selden (2011): A restudy of the Burgess Shale (Cambrian) arthropod Emeraldella brocki and reassessment of its affinities. Journal of Systematic Palaeontology 10 (2): 361-383. doi:10.1080/14772019.2011.566634
- David A. Legg, Mark D. Sutton, Gregory D. Edgecombe (2013): Arthropod fossil data increase congruence of morphological and molecular phylogenies. Nature Communications 4: 2485. doi:10.1038/ncomms3485
- Tafel aus Charles D. Walcott. Cambrian Geology and Paleontology/Volume 2/Middle Cambrian Branchiopoda, Malacostraca, Trilobita, and Merostomata, 1912 Volltext in Wikisource
Weblinks
- Are Naraoids trilobites? Abbildungen verschiedener Arten und Diskussion der kontroversen Klassifizierungen