Ostamerikanischer Maulwurf
Der Ostamerikanische Maulwurf (Scalopus aquaticus) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Er bewohnt weite Teile des östlichen Nordamerikas vom südlichsten Kanada über den zentralen und östlichen Teil der USA bis in das nördlichste Mexiko. In diesem ausgedehnten Verbreitungsgebiet nutzt die Art zahlreiche unterschiedliche Lebensräume von offenen Landschaften bis hin zu Wäldern sowohl auf Meeresspiegelniveau bis in mittlere Gebirgslagen. Hierbei bevorzugen die Tiere Standorte mit feuchten lehmigen bis sandigen Böden. Es handelt sich um einen mittelgroßen Vertreter der Neuweltmaulwürfe. Wie alle Angehörigen der Gruppe ist auch der Ostamerikanische Maulwurf an eine grabende Lebensweise angepasst. Dadurch besitzt er einen walzenförmigen Körper, einen kurzen Hals und grabschaufelartige Vorderbeine. Die Fellfärbung variiert von silbergrau bis schwarz, teilweise kommt eine auffallende Fleckenbildung vor. Typisch sind die lange Schnauze, schwimmhautartige Hautlappen zwischen den Fingern und das reduzierte Gebiss. Bezüglich der Zahnanzahl bestehen allerdings Abweichungen, da gelegentlich Zähne fehlen oder überzählige ausgebildet sind.
| Ostamerikanischer Maulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Ostamerikanischer Maulwurf (Scalopus aquaticus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Scalopus | ||||||||||||
| É. Geoffroy Saint-Hilaire, 1803 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Scalopus aquaticus | ||||||||||||
| (Linnaeus, 1758) |
Die Lebensweise des Ostamerikanischen Maulwurfs ist partiell gut erforscht. Die Tiere legen Gangsysteme im Untergrund an, die in mehreren Ebenen verlaufen: oberflächennahe Tunnel zur Nahrungssuche und tiefere Gänge als Rückzugsgebiete. In letzteren befinden sich auch mit Pflanzen ausgepolsterte Nestkammern. Oberirdisch markieren charakteristische Maulwurfshügel die Gangsysteme. Generell lebt der Ostamerikanische Maulwurf einzelgängerisch und beansprucht ein eigenes Gangsystem als Territorium. Männliche und weibliche Individuen kommen nur zur Paarungszeit zusammen. Diese findet im ersten Jahresdrittel statt. Ein Wurf umfasst zwei bis fünf Jungen. Der Nachwuchs wird nur für einen kurzen Zeitraum von rund vier Wochen aufgezogen, danach verlassen die Jungtiere das mütterliche Nest. Die Nahrung besteht aus Regenwürmern und Insekten. Zusätzlich beinhaltet sie einen gewissen Anteil an pflanzlichen Bestandteilen. Die Nahrungssuche erfolgt unterirdisch mit Hilfe des Geruchssinns.
Bereits kurz nach der Besiedlung Nordamerikas durch die Europäer wurde der Ostamerikanische Maulwurf bekannt. Die wissenschaftliche Erstbenennung der Art stammt aus dem Jahr 1758. Eine eigene Gattung erhielt sie knapp ein halbes Jahrhundert später. Vor allem im 19. Jahrhundert war die Gattung eher unter der Bezeichnung Scalops bekannt. Es werden insgesamt 16 Unterarten unterschieden, von denen einzelne aber nur wenig dokumentiert sind. Fossil traten noch weitere Arten auf, die vom heutigen Ostamerikanischen Maulwurf durch eine eigene Untergattung abgetrennt werden. Der stammesgeschichtlich früheste Nachweis datiert in das Obere Miozän vor gut 7 bis 6 Millionen Jahren. Weitere Reste sind aus dem Pliozän belegt. Der heutige Ostamerikanische Maulwurf tritt erstmals im Übergang vom Pleistozän zum Holozän in Erscheinung. Das Fundmaterial insgesamt ist überwiegend spärlich erhalten. Der heutige Bestand der Art gilt als nicht gefährdet.
Merkmale
Habitus
Der Ostamerikanische Maulwurf ist ein mittelgroßer Vertreter der Neuweltmaulwürfe. Die Kopf-Rumpf-Länge variiert von 12,9 bis 15,4 cm, die Schwanzlänge von 1,5 bis 3,0 cm. Der Schwanz nimmt dadurch weniger als ein Viertel der Gesamtlänge ein. Das Körpergewicht beträgt etwa 40 bis 64 g. Männliche Individuen werden etwa 28 % schwerer als weibliche, so dass ein deutlicher Sexualdimorphismus besteht.[1] Außerdem gibt es geographische Variationen. Die größten Tiere finden sich im zentral-nördlichen Bereich des Verbreitungsgebietes, nach Ost und West hin nimmt die Körpergröße ab, die kleinsten Formen leben im südöstlichen Abschnitt des Vorkommens. Im Körperbau ähnelt der Ostamerikanische Maulwurf anderen Vertretern der Neuweltmaulwürfe und zeigt wie diese Anpassungen an eine grabende Lebensweise. Der Körper ist dadurch walzenförmig, der Kopf sitzt auf einem kurzen Hals und die Vorderbeine sind zu Grabschaufeln umgestaltet. Als auffälliges Kennzeichen bestehen an diesen zwischen den Zehen schwimmhautartige Hautlappen, die das Graben unterstützen. Die Handflächen selbst sind breiter als lang. Die Hinterfüße wirken dem gegenüber eher schmal, ihre Länge reicht von 1,5 bis 2,2 cm. Allgemein sind die Vorder- und Hinterfüße fleischig und nackt, ebenso der Schwanz. Das Körperfell ist dicht und weich, es kann von silbergrau über bräunlich bis schwarz gefärbt sein. Rund ein Drittel aller Tiere unabhängig vom Alter weisen Farbflecken auf, die sich häufig im Bauch- oder Brustbereich, seltener am Kinn, der Schnauze oder auf dem Kopf befinden. Es dominieren hierbei orangefarbene Töne. Die Größe der Flecken liegt im Durchschnitt bei 2,81 cm², kann aber zwischen 0,04 und 25,0 cm² schwanken. Sie treten häufiger bei Männchen als bei Weibchen auf und sind nicht mit Farbveränderungen zu verwechseln, die durch Sekrete aus Drüsen verursacht werden und fast nur bei ausgewachsenen Individuen vorkommen.[2] Die Schnauze ist relativ lang ausgezogen und nackt. Wie bei vielen Maulwürfen sind die Augen und Ohren äußerlich nicht sichtbar. Die Augen erscheinen als dunkle Flecken unter dem Fell, zumeist sind sie kleiner als 1,0 mm im Durchmesser und stärker zurückgebildet als beim Europäischen Maulwurf (Talpa europaea). Da strukturell alle Elemente der Säugetieraugen vorkommen, vermögen sie wahrscheinlich noch zwischen hell und dunkel zu unterscheiden.[3] Die Ohröffnungen haben einen Durchmesser von 0,5 mm. Weibchen weisen sechs Zitzen auf, jeweils ein Paar befindet sich im Brust-, Bauch- und im Lendenbereich.[4][5][6]
Schädel- und Gebissmerkmale
Messungen an Tieren aus Texas ergaben eine Schädellänge zwischen 30,1 und 38,3 mm sowie eine Breite an den Warzenfortsätzen von 16,0 bis 19,3 mm. Die Weite im Bereich der Augen beträgt 6,3 bis 8,2 mm, die Höhe am Hirnschädel 8,8 bis 11,3 mm. Der Schädel ist konisch und flach geformt mit einem vergleichsweise breiten Hirnschädel und einem schmalen Zwischenaugenabschnitt. Die Stirnregion verläuft flach und nicht sehr stark aufsteigend, die Stirnhöhlen sind aber teils geschwollen. Das Rostrum ist kurz und am Mittelkieferknochen etwas verdickt. Seine Weite auf Höhe der Molaren liegt bei 8,3 bis 11,2 mm, auf Höhe der Eckzähne bei 3,4 bis 5,4 mm. Der Mittelkieferknochen überragt das Nasenbein deutlich. Die Nasenöffnungen richten sich nach vorn. Die Jochbögen schließen sich vollständig, sind aber relativ dünn ausgebildet und verlaufen leicht gekurvt. An der Schädelbasis heben sich die Warzenfortsätze kräftig und prominent hervor. Die Paukenblasen sind vollständig und gepresst, aber nicht so deutlich wie bei den Westamerikanischen Maulwürfen (Scapanus). Der Gaumen ist leicht verlängert und endet hinter dem letzten Mahlzahn. Am Unterkiefer weist der horizontale Knochenkörper einen kräftigen Bau auf. Sowohl am vorderen als auch am hinteren Ende steigt er leicht auf. Der Kronenfortsatz erhebt sich moderat hoch, er ist steil aufgerichtet. Der Winkelfortsatz am hinteren Ende des Unterkiefers ist eher kurz.[7][8][4][5][6]
Gegenüber den Westamerikanischen Maulwürfen ist das Gebiss des Ostamerikanischen Maulwurfs reduziert und besteht aus 36 Zähnen. Dadurch ergibt sich folgende Zahnformel: .[6] Bezüglich der Zahnanzahl bestehen aber einzelne Unterschiede und Variationen. Dies wird einerseits durch das Auftreten rudimentärer Zähne verursacht, die bei rund einem Viertel aller Tiere beobachtet werden können. Die zusätzlichen Zähne füllen zumeist die Lücke zwischen dem letzten Schneidezahn und dem ersten Prämolaren im Unterkiefer. Ihre Größe variiert erheblich von mikroskopisch klein bis sichtbar. Die Ausprägung nimmt mit steigendem Alter eines Individuums ab. Teilweise wurde in der Vergangenheit die Zahnanzahl daher mit 40 und mit einer entsprechenden Zahnformel angegeben, bei der ein zusätzlicher unterer (dritter) Schneidezahn und ein Eckzahn berücksichtigt waren.[7] Untersuchungen haben aber gezeigt, dass die rudimentären Zähne eher dem Eckzahn und dem ersten Prämolaren entsprechen.[9][10] Andererseits tritt auch eine gewisse Oligodontie auf, was wiederum bei etwa 14,5 % aller untersuchten Tiere nachgewiesen ist. Häufig fehlt ein- oder beidseitig der vordere Prämolar im Oberkiefer, mitunter auch ein Schneidezahn. Überzählige Zähne hingegen sind seltener, betreffen aber auch die Prämolaren und Schneidezähne der oberen Gebissreihe.[11] Wie bei den anderen Neuweltmaulwürfen ist der vorderste obere Schneidezahn vergrößert, die beiden nachfolgenden sind deutlich kleiner. Der obere Eckzahn erreicht etwa zwei Drittel der Höhe des ersten Schneidezahns. Im Unterkiefer hingegen überragt der zweite Schneidezahn den ersten deutlich, er ist stark hypertrophiert und eckzahnartig (caniniform). Ein dauerhafter Eckzahn besteht nicht. Der zweite obere Prämolar ist größer als der erste und dritte, in der unteren Zahnreihe nehmen die Vormahlzähne von vorn nach hinten an Größe zu. Die ersten beiden Molaren sind sowohl oben wie auch unten relativ gleich groß, der jeweils letzte ist deutlich kleiner. Sie weisen mehrere Haupthöcker auf der Kauoberfläche auf, die bei den oberen ein W-förmiges (dilambdodontes) Muster bilden. Außerdem sind die oberen Mahlzähne im Querschnitt W-förmig, die unteren M-förmig gestaltet. Die obere Zahnreihe wird zwischen 9,2 und 11,3 mm lang, die untere Prämolaren-Molaren-Reihe zwischen 9,8 und 10,8 mm.[7][8][4][5][6]

Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 34. Er entspricht dadurch weitgehend dem der meisten anderen Neuweltmaulwürfe. Alle Chromosomen, ausgenommen das X- und das Y-Chromosom, sind zweiarmig. Es kommen somit 8 metazentrische, 6 submetazentrische und 2 subtelozentrische Autosomenpaare vor. Die fundamentale Anzahl, also die Anzahl der Autosomenarme, beträgt 64. Das Y-Chromosom ist klein.[12][4][6]
Verbreitung und Lebensraum
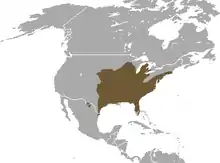
Das Verbreitungsgebiet des Ostamerikanischen Maulwurfs ist das größte aller Neuweltmaulwürfe in Nordamerika. Es erstreckt sich vom südlichen Kanada über weite Teile der östlichen und zentralen USA bis in das nordöstliche Mexiko. In Kanada ist die Art auf die Südspitze von Ontario beschränkt, wo sie auf einer Fläche von rund 10 km² im Essex County zwischen den Ortschaften Essex im Westen und Leamington im Osten einschließlich des westlichen Teils des Point-Pelee-Nationalparks vorkommt.[13] In den USA verteilt sie sich auf einen Raum von Florida im Süden entlang der Ostküste nordwärts bis nach Massachusetts und westwärts bis nach Texas, Kansas und den Südwesten von Wyoming. Die Nordgrenze verläuft durch das südliche South Dakota und Minnesota sowie Michigan. Für Mexiko wurden lediglich zwei Reliktpopulationen in den nördlichen Bundesstaaten Tamaulipas und Coahuila beschrieben, letztere ist auf die Sierra del Carmen beschränkt, einem nördlichen Ausläufer der Sierra Madre Oriental.[14][4][6]
Der Lebensraum umfasst zahlreiche unterschiedliche Landschaftstypen. Die Tiere sind in nahezu allen bewaldeten oder offenen Habitaten zu finden, letztere schließen Felder, Wiesen, Weiden, Parks und bewohnte Gebiete ein. Sie bevorzugen aber weitgehend feuchte und gut entwässerte sandige oder lehmige Böden. Stark gewässerten Untergrund beziehungsweise sehr trockene Areale meiden sie. Gleiches gilt für schwere tonige Böden, tiefgründig locker-sandige Bereiche oder steinige bis geröllige Gebiete. Derartige Substrate können als Ausbreitungsbarrieren für den Ostamerikanischen Maulwurf wirken, ebenso wie saure Böden, die das Nahrungsangebot einschränken. Flüsse hingegen beeinträchtigen die Migration der Art nicht. Der Ostamerikanische Maulwurf fehlt in den Everglades, besiedelt aber einzelne andere größere Feuchtlandschaften im Südosten der USA. Optimale Voraussetzungen für die Art bilden ein Mosaik aus Wald- und Offenlandschaften. Für die mexikanischen Bestände werden Vegetationslandschaften durchsetzt mit Mexikanischer Nusskiefer und Alligator-Wacholder angegeben.[14] Die Höhenverbreitung reicht vom Meeresspiegelniveau bis auf 1375 m in den USA und 1633 m in Mexiko. Die Populationsdichte schwankt sehr stark, basierend auf Untersuchungen in drei Gebieten in South Carolina während und kurz nach der Fortpflanzungsphase. Demnach variiert sie von 0,86 bis 12,5 Individuen je Hektar.[15] Die Art tritt im südlichen Teil des Verbreitungsgebietes häufiger auf als im nördlichen.[4][6]
Lebensweise
Territorialverhalten
.jpg.webp)
Wie alle Neuweltmaulwürfe führt der Ostamerikanische Maulwurf eine unterirdisch grabende Lebensweise. Er legt dabei ein komplexes Tunnel- und Gangsystem an, das mehrere Ebenen einschließt und bis zu 3540 m² bedecken kann. Oberflächennahe Gänge werden in der Regel nur temporär genutzt und dienen vornehmlich der Nahrungssuche. Sie bestehen aus Röhren von 3,2 bis 3,8 cm Durchmesser und sind häufig weniger als einen Meter lang, einzelne Tunnel erreichten aber auch bis zu 90 m Länge. Oberflächlich zeichnen sich diese Gänge durch charakteristische Rippeln ab. In lockeren Böden kann ein Tier täglich rund 31 m an Gängen konstruieren. Tiefere Gänge liegen zwischen 2,5 und 60 cm unter der Erdoberfläche, zumeist bei 35,5 cm. Es handelt sich bei diesen um permanentere Systeme, die bis zu fünf Jahre genutzt werden können und aufgrund ihrer Tiefe sowohl vor sommerlicher Trockenheit als auch vor winterlicher Kälte schützen. Innerhalb der tieferen Gänge befinden sich mehrere Nestkammern. Diese haben eine Länge von 18 bis 22 cm und eine Breite von 10 bis 12 cm. Zumeist befinden sie sich 8 bis 46 cm unter der Erdoberfläche und in der Nähe von Steinen, Stämmen oder Büschen. Die Nester werden mit Pflanzenmaterial aus Gräsern und Blättern ausgepolstert.[16] Eine Ausnahme bildet der Ostamerikanische Maulwurf in Florida, der offensichtlich in seinen Kammern keine Pflanzen zu Nestern anhäuft.[17] Ein einzelnes Tunnelsystem enthält zwei bis sieben Nestkammern, möglicherweise eine Kammer fungiert als dauerhaftes Winterquartier, während im Sommer unterschiedliche Nester genutzt werden.[18][19][4][5][6]
Die Grabungsaktivitäten des Ostamerikanischen Maulwurfs erfolgen mit alternierenden seitlichen Bewegungen der Vorderfüße. Der Körper ist dabei um rund 52° von der ausführenden Hand weggedreht, die restlichen Füße halten das Tier am Boden. Die Nase wird meist als taktiles Organ eingesetzt. Für die Konstruktion der oberflächennahen Tunnel presst das Tier das gelockerte Erdmaterial an die Seite. Der Aushub der tieferen Tunnel wird zunächst hinter dem Körper angesammelt und nach einer gewissen Zeit über senkrechte Gänge an die Oberfläche gebracht, wodurch die charakteristischen Auswurfhügel (Maulwurfshügel) entstehen. Dabei rotiert das Tier im Untergrund um seine Körperachse und drückt das gelockerte Erdmaterial mit den Vorderfüßen durch die Gänge. Teilweise werden dadurch bis zu das 32fache des eigenen Körpergewichts bewegt. Eine auffällige Eigenschaft ist, dass der Ostamerikanische Maulwurf keine Öffnungen oder Einbruchstellen in den Gängen toleriert und diese unmittelbar repariert.[18][19] Die Tiere graben ganzjährig, die meisten Grabungsaktivitäten finden jedoch im Frühjahr statt, wenn der Boden gut durchfeuchtet ist. Die Bodenkonsistenz bestimmt weitgehend die Grabgeschwindigkeit. In härteren Böden nimmt sie erheblich ab, teilweise von 29 m auf 1 m je Stunde, ebenso verringert sich die Tunnellänge. Dem gegenüber steigt die Menge des transportierten Aushubs sowohl absolut als auch relativ zum individuellen Körpergewicht und zur Arbeitsleistung je Stunde mitunter um das Doppelte. Auch die kontinuierlich absolvierte Grabtätigkeit reduziert sich.[20] In den Gangsystemen herrschen durch die teils fehlende Frischluftzufuhr teilweise extreme Bedingungen vor, die zu einer Anreicherung von Kohlenstoffdioxid und einer Verarmung an Sauerstoff führen. Der Ostamerikanische Maulwurf ist an diese speziellen Umgebungsbedingungen durch eine Modifikation seines Hämoglobins im Blut angepasst, die eine höhere CO2-Aufnahme ermöglicht und so mehr Sauerstoff zur Verfügung stellt. Der Effekt wirkt unbeeinflusst von den relativ niedrigen Temperaturen in den Tunneln.[21][22][4][5][6]
Der Ostamerikanische Maulwurf lebt einzelgängerisch und territorial. Jedes Individuum beansprucht ein eigenes Gangsystem, gelegentlich okkupiert es auch das Gangsystem eines anderen grabenden Tieres, etwa der Präriewühlmaus, und passt es seinen Gegebenheiten an. Nach Untersuchungen in Kentucky haben Männchen durchschnittlich größere Reviere als Weibchen. Bei ersteren sind sie zwischen 0,36 und 1,8 ha groß, bei letzteren zwischen 0,15 und 0,35 ha. Sie werden über eine längere Zeit gehalten, jahreszeitliche Schwankungen in der Größe bestehen nicht. Teilweise überlappen sich die Territorien am Rand. Dadurch kommt es mitunter dazu, dass ein Individuum das Gangsystem eines Nachbarn benutzt. Wie die meisten Maulwürfe hat auch der Ostamerikanische Maulwurf keinen ausgeprägten Tag-Nacht-Rhythmus, zudem hält er keinen Winterschlaf. Die Tiere aus Kentucky zeigten jedoch zwei stärkere Aktivitätsperioden, die zwischen 08.00 und 16.00 Uhr sowie zwischen 23.00 und 04.00 Uhr stattfinden. Aktive Phasen dauern im Durchschnitt 5,5 Stunden, inaktive 3,2 Stunden. Weibchen verbringen rund 39 % ihrer Zeit im Nest, bei Männchen sind es demgegenüber 36 %. Während ihrer Zeit im Nest verhalten sich die Tiere aber nicht immer passiv, andererseits können Ruhephasen auch außerhalb des Nestes absolviert werden. Die in 24 Stunden innerhalb des Gangsystems zurückgelegten Entfernungen betragen bis zu 414 m. In bestehenden tieferen Gängen gemessene Geschwindigkeiten beliefen sich auf 24 m je Minute. In der Regel hält sich ein Tier in der Umgebung der Nester auf. Die größte Distanz zu einem Nest wurde mit linear 204 m ermittelt. Auf dem Weg zu seinem Ruheplatz legte das Individuum 278 m durch die Gänge zurück. Der Ostamerikanische Maulwurf ist zudem ein guter Schwimmer, tätigt dies aber nur in Ausnahmefällen. Im wässrigen Milieu setzt er zumeist nur die Hinterbeine zur Fortbewegung ein.[23][4][5][6]
Ernährung
Die Nahrung des Ostamerikanischen Maulwurfs besteht hauptsächlich aus Regenwürmern, daneben nehmen auch Hundertfüßer, Insekten sowie deren Larven und Schnecken einen größeren Anteil ein.[23][24][19] Die tierischen Bestandteile umfassen rund 85 % der gesamten aufgenommenen Menge. Der Rest beläuft sich auf Pflanzen, hier sind vor allem Samen von Gräsern und Mykorrhiza hervorzuheben. Anhand von mehr als 100 Magenresten aus Indiana ergaben sich anteilig etwa 27 % Regenwürmer, 14 % Blatthornkäfer, 9 % unbestimmbares Pflanzenmaterial, 7 % Ameisen und knapp 6 % Laufkäfer. Untergeordnet waren auch Schmetterlinge, Hundertfüßer und Spinnen vertreten.[25] Relativ vergleichbar dazu erwiesen sich mehr als ein Dutzend Inhalte von Mägen von Cumberland Island in Georgia, wo Regenwürmer fast 30 % und Blatthornkäfer 26 % aller Nahrungsreste darstellten. Hinzu kamen Ameisen und Heuschrecken.[26] Dem gegenüber erbrachte die Untersuchung von über 370 Mageninhalten aus South Carolina eine von dieser generellen Ernährungsweise abweichende Zusammensetzung. Es dominierten hier Larven von Blatthornkäfern, die fast ein Drittel der Nahrungsmenge erreichten und in mehr als zwei Drittel aller analysierten Proben auftraten. Weiter häufig ließen sich Ameisen und Hundertfüßer belegen. Erstere konnten mit allein 29 Arten dokumentiert werden. Hingegen traten Regenwürmer nur in 8 % aller Mägen auf und wiesen einen Mengenanteil von 3 % auf. Insgesamt wurden 52 verschiedene Nahrungsbestandteile identifiziert. Unterschiede zwischen männlichen und weiblichen Individuen beziehungsweise zu Jungtieren bestanden nicht. Allerdings sind jahreszeitliche Variationen fassbar. So bilden die Larven der Blatthornkäfer die Hauptnahrung in den Winter- und Sommermonaten, während Ameisen häufiger im Frühjahr gefressen werden. Ähnlich verhält es sich mit dem Pflanzenmaterial, bei dem Pilze im Sommer und Herbst, Samen hingegen im Frühjahr überwiegen.[27] Gelegentlich plündert der Ostamerikanische Maulwurf Nester von Wespen.[28] Tiere in menschlicher Gefangenschaft verzehren zudem auch Mäuse, Vögel, Frösche, kleinere Schlangen und Tiernahrung. Wasser trinkt der Ostamerikanische Maulwurf nur selten.[4][5][6]
Die Nahrungssuche erfolgt in der Regel über den Geruchssinn. Als besondere Anpassung ist der Ostamerikanische Maulwurf in der Lage, mit beiden Nasenlöchern Gerüche getrennt wahrzunehmen, was man als Stereoriechen bezeichnen kann und wodurch eine Orientierung im Raum erleichtert ist.[29] Die Beute wird durch Pressen mit den Vorderfüßen gegen die Tunnelwände oder durch mehrere Bisse getötet, bei letzterem begräbt der Ostamerikanische Maulwurf sie teilweise auch unter lockerer Erde. Wenig wehrhafte Beute frisst er direkt. Die grabende Lebensweise bedingt einen hohen Energieverbrauch. Die täglich benötigte Nahrungsmenge beläuft sich auf 25 bis 100 % des eigenen individuellen Körpergewichtes, der Durchschnitt beträgt 32 %. Innerhalb von 18 Stunden kann er die Menge von rund 66 % seines eigenen Körpergewichts fressen.[24] Im Magen sammeln sich zwischen 0,8 und 1,9 g an Nahrungsresten.[27][4][5][6]
Fortpflanzung
Die Paarung findet im ersten Jahresdrittel statt und dauert etwa drei bis vier Wochen. Bei Männchen wird dies durch die starke Vergrößerung der Hoden angezeigt, die von durchschnittlich 170 bis 200 mg auf 600 bis 2500 mg anwachsen können. Der Prozess setzt bereits im Verlauf des Dezembers ein.[30] Im Süden des Verbreitungsgebietes wie in Texas und Louisiana beginnt die Fortpflanzungsphase möglicherweise schon im Januar, im restlichen zeichnet sich ein Höhepunkt im späten März und frühen April ab. Sehr selten bisher wurden trächtige Weibchen im Juni beobachtet. Während der Paarungszeit kommt es zu den wenigen bekannten Oberflächenaktivitäten beim Ostamerikanischen Maulwurf. Die Dauer der Tragzeit ist unbekannt, wird aber mit 28 bis 45 Tagen veranschlagt.[19][30][8][31] Weibchen bringen einmal jährlich einen Wurf zur Welt, der zwei bis fünf Jungtiere umfasst. Die meisten Geburten wurden zwischen März und Juni beobachtet. Der Nachwuchs ist blind und nackt, im Vergleich zur Körpergröße des Muttertiers aber schon recht groß. Die Gesamtlänge liegt durchschnittlich bei 5,3 cm, die Schwanzlänge bei 0,6 cm und das Gewicht bei rund 5 g. Manchmal kommt es zum Infantizid durch das Muttertier.[32][4][5][6]
Allgemein verläuft die Individualentwicklung nach der Geburt sehr schnell. Nach rund zehn Tagen bildet sich ein graues samtiges Fell. Bereits nach vier Wochen verlassen die Jungen das mütterliche Nest. Sie erreichen mit rund einem Jahr die Geschlechtsreife. Allerdings stirbt mehr als die Hälfte der Jungtiere in den ersten sechs Monaten. Die ausgewachsenen Tiere hingegen haben eine große Überlebensrate. Weibliche Tiere können im Maximum 6,2 Jahre alt werden, männliche 5,9. Der Ostamerikanische Maulwurf lebt dadurch länger, als es Berechnungen basierend auf seiner Körpergröße annehmen ließen.[33] In der Regel beträgt das durch Beobachtungen ermittelte Geschlechterverhältnis 1:1, nur während der Paarungszeit treten aufgrund verstärkter Oberflächenaktivitäten mehr Männchen in Erscheinung.[34] Eine in Kansas untersuchte Population aus 196 Individuen setzte sich aus insgesamt 66 Jungtieren, 68 Jährlingen, 50 Tieren im zweiten Lebensjahr und 12 älteren Exemplaren zusammen. Das Verhältnis zwischen männlichen und weiblichen Tieren betrug durchschnittlich 1:1,8. Es nahm aber vor allem bei den älteren Individuen zugunsten der Weibchen zu und konnte bei betagten Tieren dann 1:9 erreichen.[35] Auch Studien an 240 Tieren aus dem südlichen Illinois erbrachten ein Überwiegen weiblicher Individuen. Hier wurden aber in der ersten Jahreshälfte häufiger Weibchen festgestellt, in der zweiten dem gegenüber Männchen. Die Gründe für die auftretenden regionalen Unterschiede sind bisher noch nicht geklärt.[1][4][5][6]
Abweichend von zahlreichen Vertretern der Eurasischen Maulwürfe (Talpa) oder dem Sternmull (Condylura) und übereinstimmend mit den Westamerikanischen Maulwürfen (Scapanus) gibt es beim Ostamerikanischen Maulwurf keinen Hinweis auf einen ausgeprägten Hermaphroditismus etwa in Form von Zwitterdrüsen.[36]
Fressfeinde und Parasiten
Die unterirdische Lebensweise des Ostamerikanischen Maulwurfs führt dazu, dass er nur selten im Nahrungsspektrum von Beutegreifern auftritt. Hinzu kommt sein moschusartiger Geruch, den zahlreiche potentielle Fressfeinde meiden. Unter den Säugetieren wäre der Kojote zu nennen, der nach Studien in North Carolina im Sommer und Herbst vereinzelt ein Tier stellt.[37] Vergleichbares ist zum Rotfuchs anhand von Kotanalysen aus Iowa zu sagen.[38] Vögel sind unter anderem mit dem Rotschwanzbussard vertreten, der gelegentlich ein Individuum erlegt.[39] Andere Predatoren finden sich mit der Schleiereule und der Östlichen Fuchsnatter, letztere kann in die Tunnel eindringen. Eventuell als Kommensalen treten Amerikanische Kurzschwanzspitzmäuse innerhalb der Gangsysteme auf.[5]
Als äußere Parasiten wurden unter anderem Flöhe der Gattungen Ctenophthalmus und Corypsylla beschrieben, des Weiteren auch Milben wie Hirstionyssus, Androlaelaps, Eulaelaps und Haemogamasus aus der Gruppe der Raubmilben beziehungsweise Pygmaephorus aus der Gruppe der Prostigmata sowie Scalopacarus aus der Gruppe der Hornmilben. Mit Leptinus ist auch ein Käfer nachgewiesen, der aber möglicherweise keinen echten Parasiten darstellt, sondern in den Nestern haust und sich von den Milben ernährt.[40][25][41] Zu den inneren Parasiten zählen Saugwürmer wie Ityogonimus, Bandwürmer wie Hymenolepis, Fadenwürmer wie Physaloptera, Rictularia und Strongyloides sowie Kratzwürmer wie Macracanthorhynchus und Moniliformis.[42][43]
Systematik
Innere Systematik der Neuweltmaulwürfe nach Chen et al. 2021[44]
|
Der Ostamerikanische Maulwurf ist eine Art aus der Gattung Scalopus, er stellt deren einzigen rezenten Vertreter dar, womit Scalopus aus heutiger Sicht als monotypisch gilt. Art und Gattung gehören wiederum zur Familie der Maulwürfe (Talpidae). Innerhalb dieser bilden sie zusammen mit einigen weiteren in Asien und Nordamerika verbreiteten Formen die Tribus der Neuweltmaulwürfe (Scalopini). Die Neuweltmaulwürfe repräsentieren, ähnlich den Eigentlichen Maulwürfen (Talpini), grabende Angehörige der Familie, allerdings sind die beiden Gruppen nicht unmittelbar verwandt, sondern entwickelten ihre Grabeigenschaften unabhängig. Andere Familienmitglieder wiederum leben nur teilweise unterirdisch, bewegen sich oberirdisch fort oder sind an eine semi-aquatische Lebensweise angepasst.[45] Als besondere Kennzeichen der Neuweltmaulwürfe können der gegenüber den Eigentlichen Maulwürfen deutlich längeren Schwanz, der stark vergrößerte vordere obere Schneidezahn und der auftretende Zahnwechsel herausgestellt werden. Übereinstimmend mit den Eigentlichen Maulwürfen kommt an der Hand ein zusätzliches Sesambein vor dem Daumen vor, der sogenannte Präpollex („Vordaumen“), das die Handfläche verbreitert.[46][6] Nach molekulargenetischen Untersuchungen erfolgte die Abtrennung der Neuweltmaulwürfe von den anderen Triben der Maulwürfe im Oberen Eozän vor rund 39 bis 35 Millionen Jahren.[45] Die Tribus teilt sich in zwei Entwicklungslinien: die Parascalopina und die Scalopina. Beide sind anhand der Ausprägung des Metastylids am unteren zweiten Molar unterscheidbar, das bei den Scalopina fehlt, bei den Parascalopina hingegen vorkommt.[47][48] Die zwei Linien bestehen wenigstens seit dem Unteren Miozän vor 21,4 Millionen Jahren als eigenständig. Der Ostamerikanische Maulwurf gehört zu den Scalopina, die nächsten Verwandten bilden die Westamerikanischen Maulwürfe (Scapanus). Beide Gattungen diversifizierten sich vermutlich schon im Mittlere Miozän vor gut 14 Millionen Jahren.[49][44]
In der Regel werden 16 Unterarten des Ostamerikanischen Maulwurfs unterschieden:[4][50][6][51]
- S. a. aereus (Bangs, 1896); USA: südöstliches Colorado, südlich-zentrales Kansas, nordöstliches Arkansas, südliches Louisiana und östliches sowie nord-zentrales Texas einschließlich Texas Panhandle
- S. a. alleni R. H. Baker, 1951; USA: Teile des südlichen und süd-zentralen Texas
- S. a. anastasae (Bangs, 1898); USA: nur auf Anastasia Island in Florida
- S. a. aquaticus (Linnaeus, 1758); Nominatform; USA: östliches und südliches Massachusetts, südöstliches New York, südöstliches Pennsylvania, nordöstliches West Virginia, Virginia, westliches North Carolina und östliches Tennessee, vom Meeresspiegelniveau bis auf 1220 m Höhe in den Appalachen
- S. a. australis (Chapman, 1893); USA: südöstliches Georgia und östliches Florida südlich bis Hypoluxo im Palm Beach County
- S. a. bassi A. H. Howell, 1939; USA: nur von Engelwood im Sarasota County in Florida bekannt
- S. a. caryi Jackson, 1914; USA: südöstliches Wyoming, zentrales und westliches Nebraska, nordöstliches Colorado und nordwestliches Kansas; hauptsächlich in den trockenen und halbtrockenen Abschnitten der Great Plains
- S. a. cryptus W. B. Davis, 1942; USA: östliches Texas
- S. a. howelli Jackson, 1914; USA: North Carolina (außer den Appalachen), South Carolina, nördliches und westliches Georgia, Florida Panhandle, Alabama, Mississippi und Louisiana östlich des Mississippi
- S. a. inflatus Jackson, 1914; USA und Mexiko: südöstliche Spitze von Texas bis nach Tamaulipas
- S. a. machrinoides Jackson, 1914; USA: westlich des Mississippi, zentrales Minnesota, südöstliches South Dakota, östliches Nebraska, östliches Kansas und nördliches Arkansas
- S. a. machrinus (Rafinesque, 1832); Kanada und USA: östlich des Mississippi und westlich der Appalachen, westliches Wisconsin, nördliches Illinois, südliches Michigan, südwestliches Ontario, nördliches Ohio und zentrales bis westliches Tennessee
- S. a. montanus R. H. Baker, 1951; Mexiko: nur in der Sierra del Carmen im nördliches Coahuila
- S. a. parvus (Rhoads, 1894); USA: zentrales und westliches Florida
- S. a. porteri Schwartz, 1952; USA: nur im Miami-Dade County in Florida
- S. a. texanus (J. A. Allen, 1891); USA: im Big-Bend-Nationalpark in Texas
In einigen Publikationen wird S. a. nanus anstatt S. a. cryptus geführt,[4][50][6] beide Formen gelten als identisch. Sie wurden von William B. Davis im Jahr 1942 in der gleichen Publikation anhand von Individuen aus dem östlichen Texas vorgestellt, erstere Form begründet sich auf ein weibliches Tier aus dem Leon County, letztere auf ein männliches aus dem Brazos County.[52] Eine Revision des Ostamerikanischen Maulwurfs durch Terry L. Yates und David J. Schmidly im Jahr 1977 listete nur S. a. cryptus auf und setzte S. a. nanus mit diesem gleich.[8] Im Jahr darauf gaben beide Autoren in einem Artportrait über den Ostamerikanischen Maulwurf, erschienen im Fachblatt Mammalian Species, jedoch S. a. nanus Priorität, möglicherweise da Davis diese Form zuerst benannt hatte.[4] Eine erneute Revision aus dem Jahr 2018 durch Neal Woodman erkannte dem gegenüber S. a. cryptus als korrekt an, da diese durch die erste Revision als gültig benannt worden war.[51]
Fossil wurden mehrere weitere Arten der Gattung Scalopus zugewiesen. Alle stehen innerhalb einer eigenständigen Untergattung mit der Bezeichnung Hesperoscalops. Diese zeichnet sich durch relativ hochkronige Zähne und eine geringere Reduktion der Größe des letzten Mahlzahns gegenüber dem vorletzten in der unteren Zahnreihe aus. Hinzu kommen einzelne weitere Zahnmerkmale wie zusätzliche Höckerchen auf den Kauoberflächen der Molaren. Insgesamt gliedert sich die Gattung Scalopus dadurch folgendermaßen:[53][54]
- Untergattung Scalopus É. Geoffroy Saint-Hilaire, 1803
- Scalopus aquaticus (Linnaeus, 1758)
- Untergattung Hesperoscalops Hibbard, 1941
- Scalopus blancoensis (Dalquest, 1975)
- Scalopus mcgrewi Voorhies, 1977
- Scalopus rexroadi (Hibbard, 1941)
- Scalopus ruficervus Dalquest, 1983
- Scalopus sewardensis (K. Reed, 1962)
Hesperoscalops war im Jahr 1941 von Claude W. Hibbard anhand eines Unterkiefers und einzelner isolierter Zähne von der oberpliozänen Rexroad local fauna in Kansas als eigenständige Gattung eingeführt worden.[55] Gut acht Jahre später stellte er Hesperoscalops in eine direkte Beziehung mit Scalopus und sah ersteren als Vorgängerform von letzterer an.[56] Im Jahr 1967 stufte Leigh Van Valen in einer Überblicksarbeit zu rezenten und fossilen Insektenfressern Hesperoscalops jedoch als identisch mit Scalopus ein.[57] Nahezu zeitgleich und unabhängig von Van Valen kam J. Howard Hutchinson zu dem Schluss, dass beide Formen nicht genügend einzigartige Merkmale besitzen, um eine generische Trennung zu rechtfertigen. Er stufte daher Hesperoscalops als Untergattung von Scalopus ein.[47] Der Status hat sich in der Nachfolgezeit nicht geändert.[54]
Forschungsgeschichte



Neben dem Sternmull gehört der Ostamerikanische Maulwurf zu den ersten Vertretern der Familie, die aus Nordamerika in der westlichen Welt bekannt wurden. Die wissenschaftliche Erstbenennung stammt von Linnaeus. In der zehnten Auflage seines Werkes Systema Naturae aus dem Jahr 1758 führte er den Ostamerikanischen Maulwurf unter der Bezeichnung Sorex aquaticus und verwies ihn damit zu den Spitzmäusen. Als besondere Kennzeichen hob Linnaeus auf die mit schwimmhautartigen Hautlappen versehenen Hände und Füße hervor. Das Artepitheton stellt dabei die vermeintlich amphibische Lebensweise heraus. Die Typusregion gab er America septentrionali an, was das damals bekannte Nordamerika entlang der Ostküste meint. Erst im Jahr 1915 legte Hartley H. T. Jackson dies genauer mit Philadelphia in Pennsylvania fest.[7] Linnaeus verwies in seiner kurzen Darstellung auf Albert Seba als Referenz. Seba hatte in seinem Werk Thesaurus aus dem Jahr 1734 einen Maulwurf namens Talpa virginianus niger benannt.[58] Der beigefügten Abbildung stand nach Meinung einzelner Wissenschaftler möglicherweise eher der Europäische Maulwurf Pate, doch Linnaeus' Beschreibung verweist durch die Angabe der Schwimmhäute eindeutig auf den Ostamerikanischen Maulwurf.[59] Die Artbezeichnung Talpa virginianus wurde später noch mehrfach verwendet, gilt aber als synonym zu Scalopus aquaticus. Die postulierte semi-aquatische Lebensweise des Ostamerikanischen Maulwurfs verleitete später, im Jahr 1811, Johann Karl Wilhelm Illiger dazu, den deutschen Trivialnamen „Wasserwurf“ zu prägen.[60] Dieser findet sich unter anderem auch bei Johann Andreas Wagner im Jahr 1855[61] und in Abwandlung als „Wassermaulwurf“ bei Leopold Fitzinger im Jahr 1868.[62] Ein weiteres genutztes wissenschaftliches Synonym ist Talpa canadensis, kreiert von Anselme Gaëtan Desmarest im Jahr 1820,[63] das fünf Jahre darauf auch von Richard Harlan geführt wurde. Letzterer etablierte gleichzeitig auch Talpa pennsylvanica.[64][7][51]

Eine eigenständige Gattungsbezeichnung erhielt der Ostamerikanische Maulwurf bereits Anfang des 19. Jahrhunderts. Nach heutiger anerkannter Meinung zeichnet Étienne Geoffroy Saint-Hilaire dafür verantwortlich. Er führte im Jahr 1803 Scalopus als Gattungsnamen ein. Allerdings verwies er dabei auf Georges Cuvier, der bereits drei Jahre zuvor Scalope beziehungsweise Scalops in einer Tabellenübersicht innerhalb seines Werkes Leçons d’anatomie comparée gebraucht hatte.[65] Da er jedoch keine weiterführenden Merkmale aufzeigte, gilt die Bezeichnung als Nomen nudum. Saint Hilaire dem gegenüber gab eine recht umfassende Beschreibung ab, in der er vor allem auf die Gebissstruktur und die Fußgestaltung Bezug nahm. Innerhalb der Gattung Scalopus führte Saint Hilaire den Sternmull, der später als unabhängig ausgelagert wurde, und den Ostamerikanischen Maulwurf unter der Bezeichnung Scalopus virginianus.[66] Gut 100 Jahre später korrigierte dies Theodore Sherman Palmer in Scalopus aquaticus als Nominatform. Palmer gab auch die Herkunft des wissenschaftlichen Namens aus dem Griechischen an. Hierbei besteht sowohl die Möglichkeit einer Herleitung von σκάλλω (skallow) für „scharren“, „schürfen“ oder „graben“ und πούς (pous) für „Fuß“ als auch von σκάλοψ beziehungsweise σκάλοπος (skalops beziehungsweise skalopos), was einen „Maulwurf“ meint.[67] Nach Saint Hilairs Einführung von Scalopus wurde der Name nur selten verwendet. Vielmehr hatte Illiger im Jahr 1811 Cuviers Bezeichnung Scalops aufgegriffen und mit einer ausführlichen Beschreibung versehen.[60] Diese fand dadurch weite Verbreitung. Das heute gebräuchliche Scalopus erlangte erst Anfang des 20. Jahrhunderts wieder an Bedeutung, so durch Palmer im Jahr 1904[67] und durch Gerrit S. Miller im Jahr 1912.[68]
Gattung und Art des Ostamerikanischen Maulwurfs waren im Laufe der Forschungsgeschichte mehrfach im Fokus von Revisionen. Hierzu gehören die Arbeit von Frederick W. True aus dem Jahr 1896,[69] von Hartley H. T. Jackson aus dem Jahr 1915,[7] von Terry L. Yates und David J. Schmidly aus den Jahren 1977 und 1978[8][4] sowie von Neal Woodman aus dem Jahr 2018.[51] Vor allem Yates und Schmidly arbeiteten die heutige Gliederung mit sechzehn Unterarten heraus und hoben die teils zuvor verschiedentlich getätigte Aufteilung der Gattung Scalopus in mehrere Arten, wie etwa bei Jackson 1914 zu finden, auf.[8][4]
Stammesgeschichte
Allgemein sind die Neuweltmaulwürfe recht häufig im Fossilbericht dokumentiert, Funde liegen sowohl aus der Neuen als auch der Alten Welt vor. Die Gattung Scapalus ist hierbei in der Regel nur durch einzelne Fossilreste vertreten, die räumlich und zeitlich recht weit streuen. Die älteren Nachweise werden alle der Untergattung Hesperoscalops zugesprochen. Erstmals erschienen frühe Vorfahren des Ostamerikanischen Maulwurfs im Oberen Miozän. Hierzu gehören die Arten Scalopus ruficervus und Scalopus mcgrewi. Erstere Form wurde anhand eines Unterkiefers mit den beiden vorderen erhaltenen Mahlzähnen aus der Coffee Ranch Quarry im Hemphill County des US-Bundesstaates Texas benannt. Die dortigen Ablagerungen gehen auf einen ehemaligen See zurück, an dessen Ufern sich eine vielfältige Fauna tummelte.[70] Das Fundobjekt lag etwa 1,5 m unterhalb einer Schicht aus vulkanischer Asche, deren Alter rund 6,6 Millionen Jahre beträgt. Die Größe des Unterkiefers lässt auf ein recht kleines Individuum schließen.[53] Die letztere Art ist nur unwesentlich jünger und gehört in den Übergang vom Miozän zum Pliozän vor circa 5 Millionen Jahren. Ihr Fundmaterial umfasst einzelne obere und untere Kieferfragmente, isolierte Zähne sowie Reste des postcranialen Skelettes. Es entstammt der Fundstelle Santee im Knox County von Nebraska. Die großen Tiere wiesen schon die für den Ostamerikanischen Maulwurf typische reduzierte Zahnanzahl auf. Der Oberarmknochen, von dem allein 50 Exemplare vorliegen, entspricht in seiner Robustizität dem der heutigen Art. Auffallend an der Elle ist jedoch ein Einzug am Olecranon, dem oberen Gelenkfortsatz, der bei allen Vertretern der Neuweltmaulwürfe vorkommt, jedoch beim rezenten Ostamerikanischen Maulwurf fehlt.[71] Wiederum in das Untere Pliozän gehört ein Unterkiefer aus dem Saw Rock Canyon im südwestlichen Kansas, der Scalopus sewardensis zugesprochen wird.[72] Die weiteren Formen sind dem Oberen Pliozän zuzuweisen. Auf ein vergleichsweise recht umfangreiches Fundmaterial kann Scalopus rexroadi verweisen. Es handelt sich um die Typusform der Untergattung Hesperoscalops. Die Fossilreste sind Teil der umfassenden Rexroad local fauna und kamen in der Rexroad-Formation im Meade County von Kansas zum Vorschein. Sie setzen sich aus Einzelzähnen, Kieferresten beziehungsweise Teilen des Bewegungsapparates zusammen und verweisen auf einen großen Maulwurf, etwa in der Größe des Townsend-Maulwurfes (Scapanus townsendii) aus der Gruppe der Westamerikanischen Maulwürfe.[55][56] Zu der Art werden auch mehr als drei Dutzend Funde an Unterkiefern, isolierten Zähnen und Vorderbeinelementen der Beck Ranch local fauna im zentralen Texas gezählt.[73] Aus vergleichbaren Skelettelementen setzt sich das Belegmaterial von Scalopus blancoensis zusammen, eine ebenfalls große Form, die aus der Blanco local fauna im Crosby County in Texas beschrieben wurde.[74] Die Art kam möglicherweise auch im heutigen New Mexico vor, wie Reste in Form eines unteren Vorderbeins aus Truth or Consequences und eines Unterkiefers aus Caballo vorliegen, beide Fundstellen gehören der Palomas-Formation an, welche wiederum im Sierra County im Südwesten des Bundesstaates aufgeschlossen ist.[75] Ein weiterer Unterkiefer mit fraglicher Zuweisung wurde aus dem Valencia County im zentralen New Mexico berichtet.[76]
Der heutige Ostamerikanische Maulwurf verfügt dem gegenüber über sehr spärliches Fossilmaterial. Einzelne Zähne sowie ein Unterkiefer aus der Schulze Cave im Edwards County von Texas werden in den Übergang vom Pleistozän zum Holozän datiert.[77] Subfossil sind Reste aus dem Tal des Ohio im zentralen und westlichen Pennsylvania aufgefunden worden, etwa vom Sheeps Rock Shelter im Huntingdon County oder von der McKees Rocks Indian Site im Allegheny County, deren Alter zwischen 500 und 1600 n. Chr. einzuordnen ist. Die Art tritt heute in der Region nicht mehr auf.[78][79]
Bedrohung und Schutz
Die IUCN stuft den Bestand des Ostamerikanischen Maulwurfs als „nicht gefährdet“ (least concern) ein. Begründet wird dies mit der weiten Verbreitung der Art, ihrem häufigen Auftreten und ihrer Toleranz gegenüber einem gewissen Grad an Landschaftsveränderungen. Es sind keine größeren Bedrohungen für die Gesamtpopulation bekannt. Lokal können Konflikte mit dem Menschen vorkommen, was sich hauptsächlich in den Grabungsaktivitäten der Tiere begründet. Des Weiteren sind regional einzelne Bestandseinschränkungen möglich. In Kanada tritt die Art nur im südlichsten Ontario auf einer Fläche von 10 km² auf und ist dort von besonderem Interesse, zumal der Bestand, der im Jahr 1997 auf 2120 bis 12.760 Individuen geschätzt wurde, bis 2007 um 26 % zurückgegangen ist.[80][13] Die Unterart S. a. texanus ist nur von ihrer Erstsichtung im Jahr 1887 aus dem Presidio County in Texas bekannt, während die Unterart S. a. montanus im mexikanischen Bundesstaat Coahuila seit dem Jahr 1953 nicht mehr beobachtet wurde. Auch die zweite Unterart in Mexiko, S. a. inflatus im Bundesstaat Tamaulipas hat starke Einbußen aufgrund der Ausdehnung landwirtschaftlicher Nutzflächen hinnehmen müssen. Die Tiere treten in verschiedenen Naturschutzgebieten auf.[81][6]
Literatur
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 601) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Charles W. Schwartz und Elizabeth R. Schwartz: The wild mammals of Missouri. 2nd revised edition. University of Missouri Press, 2001, S. 48–53
- Terry L. Yates und David J. Schmidly: Scalopus aquaticus. Mammalian Species 105, 1978, S. 1–4
Einzelnachweise
- George A. Feldhamer, Jill C. Devine und Sara M. Ressing: Age-Related Morphometrics and Sex Ratios in the Eastern Mole (Scalopus aquaticus). The American Midland Naturalist 169, 2013, S. 409–415
- Ava A. Kamm, George A. Feldhamer und John D. Reeve: Pelage Spotting and Staining in Eastern Moles (Scalopus aquaticus). Northeastern Naturalist 15 (2), 2008, S. 303–308
- James Rollin Slonaker: The eye of the common mole, Scalopus aquaticus machrinus. Journal of Comparative Neurology 12 (4), 1902, S. 335–366 ()
- Terry L. Yates und David J. Schmidly: Scalopus aquaticus. Mammalian Species 105, 1978, S. 1–4
- Charles W. Schwartz und Elizabeth R. Schwartz: The wild mammals of Missouri. 2nd revised edition. University of Missouri Press, 2001, S. 48–53
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 601) ISBN 978-84-16728-08-4
- Hartley H. T. Jackson: A review of the American moles. North American Fauna 38, 1915, S. 1–100 (S. 27–54)
- Terry L. Yates und David J. Schmidly: Systematics of Scalopus aquaticus (Linnaeus) in Texas and adjacent states. Occasional Papers Museum Texas Tech University 45, 1977, S. 1–36 ()
- Clinton Conaway und Stuart O. Landry, Jr.: Rudimentary mindibular teeth of Scalops aquaticus. Journal of Mammalogy 39 (1), 1958, S. 58–60
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- George A. Feldhammer und Brenna N. Towery: Dental Anomalies in the Eastern Mole (Scalopus aquaticus). The American Midland Naturalist 165 (2), 2011, S. 421–425
- Terry L. Yates und David J. Schmidly: Karyotype of the Eastern mole (Scalopus aquaticus), with comments on the karyology of the family Talpidae. Journal of Mammalogy 56 (4), 1975, S. 902–905
- Environment Canada: Management Plan for the Eastern Mole (Scalopus aquaticus) in Canada. Species at Risk Act Management Plan Series, Environment Canada, Ottawa, 2015, S. 1–18
- Iván Castro-Arellano und Gerardo Ceballos: Scalopus aquaticus (Linnaeus, 1758) Eastern Mole. In: Gerardo Ceballos (Hrsg.): Mammals of Mexico. Johns Hopkins University Press, 2014, S. 491–492
- George D. Hartman und John D. Krenz: Estimating population density of moles Scalopus aquaticus using assessment lines. Acta Theriologica 38 (3), 1993, S. 305–314
- Michael J. Harvey: Home Range, Movements, and Diel Activity of the Eastern Mole, Scalopus aquaticus. The American Midland Naturalist 95 (2), 1976, S. 436–445
- Larry N. Brown: Unique features of tunnel systems of the Eastern mole in Florida. Journal of Mammalogy 53 (2), 1972, S. 394–395
- Frederick L. Hisaw: Observation on the burrowing habits of moles (Scalopus aquaticus machrinoides). Journal of Mammalogy 4 (2), 1923, S. 79–88
- A. V. Arlton: An ecological study of the mole. Journal of Mammalogy 17 (4), 1936, S. 349–371
- Y.‐F. Lin, A. Chappuis, S. Rice und E. R. Dumont: The effects of soil compactness on the burrowing performance of sympatric eastern and hairy‐tailed moles. Journal of Zoology 301 (4), 2017, S. 310–319, doi:10.1111/jzo.12418
- Kevin L. Campbell, Jay F. Storz, Anthony V. Signore, Hideaki Moriyama, Kenneth C. Catania, Alexander P. Payson, Joseph Bonaventura, Jörg Stetefeld und Roy E. Weber: Molecular basis of a novel adaptation to hypoxic-hypercapnia in a strictly fossorial mole. BMC Evolutionary Biology 10, 2010, S. 214, doi:10.1186/1471-2148-10-214
- Anthony V. Signore, Jörg Stetefeld, Roy E. Weber und Kevin L. Campbell: Origin and mechanism of thermal insensitivity in mole hemoglobins: a test of the ‘additional’ chloride binding site hypothesis. Journal of Experimental Biology 215 (3), 2012, S. 518–525, doi:10.1242/jeb.063669
- F. A. Hanawalt: Habits of the common mole, Scalopus aquaticus machrinus (Rafinesque). Ohio Journal of Science 22, 1922, S. 164–169
- Frederick L. Hisaw: Feeding habits of moles. Journal of Mammalogy 4 (1), 1923, S. 9–20
- John O. Whitaker, Jr. und Larry L. Schmeltz: Food and External Parasites of the Eastern Mole, Scalopus aquaticus, from Indiana. Proceedings of the Indiana Academy of Sciences 83, 1974, S. 478–481
- John O. Whitaker, Jr. und Carol Ruckdeschel: Food of Eastern Moles, Scalopus aquaticus, on Cumberland Island, Georgia. Georgia Journal of Science 71 (2), 2013, S. 167–172
- Gregory D. Hartman, John O. Whitaker, Jr. und Jack R. Munsee: Diet of the Mole Scalopus aquaticus from the Coastal Plain Region of South Carolina. The American Midland Naturalist 144, 2000, S. 342–351
- Fred E. Brooks: Moles destroy wasps' nests. Journal of Mammalogy 4 (3), 1923, S. 183
- Kenneth C. Catania: Stereo and serial sniffing guide navigation to an odour source in a mammal. Nature Communications 4, 2012, S. 1441, doi:10.1038/ncomms2444
- Clinton H. Conaway: The reproductive cycle of the Eastern mole. Journal of Mammalogy 40 /2), 1959, S. 180–194
- Gregory D. Hartman: Seasonal Effects on Sex Ratios in Moles Collected by Trapping. The American Midland Naturalist 133 (2), 1995, S. 298–303
- Gregory D. Hartman und Jack L. Gottschang: Notes on sex determination, neonates, and behavior of the Eastern mole, Scalopus aquaticus. Journal of Mammalogy 64 (3), 1983, S. 539–540
- Gregory D. Hartman: Age determination, age structure, and longevity in the mole Scalopus aquaticus (Mammalia: Insectivora). Journal of Zoology 237 (1), 1995, S. 107–122
- J. M. Bedford, O. B. Mock, S. K. Nagdas, V. P. Winfrey und G. E. Olson: Reproductive features of the eastern mole (Scalopus aquaticus) and star-nose mole (Condylura cristata). Journal of Reproduction and Fertility 117, 1999, S. 345–353
- Forrest W. Davis und Jerry R. Choate: Morphologic variation and age structure in a population of the Eastern mole, Scalopus aquaticus. Journal of Mammalogy 74 (4), 1993, S. 1014–1025
- N. M. Rubenstein, G. R. Cunha, Y. Z. Wang, K. L. Campbell, A. J. Conley, K. C. Catania, S. E. Glickman und N. J. Place: Variation in ovarian morphology in four species of New World moles with a peniform clitoris. Reproduction 126, 2003, S. 713–719
- Morgan B. Swingen, Christopher S. DePerno und Christopher E. Moorman: Seasonal Coyote Diet Composition at a Low-Productivity Site. Southeastern Naturalist 14 (2), 2015, S. 397–404
- Thos G. Scott: Comparative analysis of red fox feeding trends on two central Iowa areas. Research Bulletin (Iowa Agriculture and Home Economics Experiment Station) 28 (353), 1947, S. 427–487
- Mark Andrew Springer und John Stephen Kirkley: Inter and intraspecific interactions between Red-tailed hawks and Great horned owls in Central Ohio. Ohio Journal of Science 78 (6), 1978, S. 323–328
- A. Fain und John O. Whitaker, Jr.: Phoretic hypopi of North American mammals (Acarina: Sarcoptiformes, Glycyphagidae). Acarologia 15, 1973, S. 144–170
- Terry L. Yates, Danny B. Penc und Glenn K. Launchbaugh: Ectoparasites from seven species of North American moles (Insectivora: Talpidae). Journal of Medical Entomology 16, 1979, S. 166–168
- John R. Olive: Some parasites of the Prairie mole, Scalopus aquaticus machrinus (Rafinesque). Ohio Journal of Science 50, 1950, S. 263–266
- John M. Kinsella und Vasyl V. Tkach: Checklist of helminth parasites of Soricomorpha (= Insectivora) of North America north of Mexico. Zootaxa 1969, 2009, S. 36–58
- Zhong-Zheng Chen, Shui-Wang He, Wen-Hao Hu, Wen-Yu Song, Kenneth O Onditi, Xue-You Li und Xue-Long Jiang: Morphology and phylogeny of scalopine moles (Eulipotyphla: Talpidae: Scalopini) from the eastern Himalayas, with descriptions of a new genus and species. Zoological Journal of the Linnean Society, 2021, S. zlaa172, doi:10.1093/zoolinnean/zlaa172
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang, Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumenting the polydactyly ‚constraint‘: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- J. Howard Hutchinson: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117
- Achim Schwermann, Kai He, Benjamin J. Peters, Thorsten Plogschties und Gabrielle Sansalone: Systematics and macroevolution of extant and fossil scalopine moles (Mammalia, Talpidae). Palaeontology 62 (4), 2019, S. 661–676, doi:10.1111/pala.12422
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Neal Woodman: American Recent Eulipotyphla. Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithsonian Institution Scholary Press, 2018, S. 1–107 (S. 20–25)
- William B. Davis: The Moles (Genus Scalopus) of Texas. The American Midland Naturalist 27 (2), 194, S. 380–386
- Walter W. Dalquest: Mammals of the Coffee Ranch local fauna, Hemphilian of Texas. The Pearce-Sellards Series 38, 1983, S. 1–41
- Gregg F. Gunnell, Thomas M. Bown, J. Howard Hutchison und Jonathan I. Bloch: Lipotyphla. In: C. M. Janis, G. F. Gunnell und M. D. Uhen (Hrsg.): Evolution of Tertiary Mammals of North America, Vol. 2. Cambridge University Press, 2007, S. 89–125
- Claude W. Hibbard: New Mammals from the Rexroad Fauna, Upper Pliocene of Kansas. The American Midland Naturalist 26 (2), 1941, S. 337–368
- Claude W. Hibbard: The Insectivores of the Rexroad Fauna, Upper Pliocene of Kansas. Journal of Paleontology 27 (1), 1953, S. 21–32
- Leigh Van Valen: New Paleocene insectivores and insectivore classification. Bulletin of the American Museum of Natural History 135 (5), 1967, S. 219–284 (S. 264)
- Albert Seba: Locupletissimi rerum naturalium thesauri accurata descriptio, et iconibus artificiosissimis expressio, per universam physices historiam. Amsterdam, 1734, Taf. 32 sowie S. 51 ()
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 53 ()
- Johann Karl Wilhelm Illiger: Prodromus Systematis Mammalium et Avium. Berlin, 1811, S. 1–301 (S. 126) ()
- Johann Andreas Wagner: Die Säugthiere Abbildungen nach der Natur mit Beschreibungen von Dr. Johann Christian Daniel von Schreber. Leipzig, 1855 Supplementband 5. S. 571–574 () und Tafelband Tafel 158 ()
- Leopold Fitzinger: Die natürliche Familie der Maulwürfe (Talpae) und ihre Arten, nach kritischen Untersuchungen. Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 59, 1868, S. 353–432 (S. 375–392) ()
- Anselme Gaëtan Desmarest: Mammalogie, ou, Description des espèces de mammifères. Premiere Partie. Paris, 1820, S. 1–276 (S. 155) ()
- Richard Harlan: Fauna americana: being a description of the mammiferous animals inhabitating North America. Philadelphia, 1825, S. 1–325 (S. 32) ()
- Georges Cuvier: Leçons d’anatomie comparée, Tome Premier. Paris, 1800, S. 1–521 (Tabelle 1) ()
- Étienne Geoffroy Saint-Hilaire: Catalogue des Mammifères du Museum National d'histoire naturelle. Paris, 1803, S. 1–272 (S. 77) ()
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 620–621 ()
- Gerrit S. Miller: List of North American land mammals in the United States National Museum, 1911. Bulletin of the United States National Museum 79, 1912, S. 1–455 (S. 7–8) ()
- Frederick W. True: A revision of the American moles. Proceedings of the United States National Museum 19 (1101), 1896, S. 1–112 (S. 19–47)
- Walter W. Dalquest: Carnivores of the Coffee Ranch (Type Hemphillian) local fauna. Bulletin of the Texas Memorial Museum 15, 1969, S. 1–44
- M. R. Voorhies: Fossil moles of the Late Hemphillian age from Northwestern Nebraska. Transactions of the Nebraska Academy of Sciences 4, 1977, S. 129–138
- Katherine M. Reed: Two new species of the fossil talpid insectivores. Breviora 168, 1962, S. 1–7 ()
- Walter W. Dalquest: Early Blancan mammals of the Beck Ranch local fauna of Texas. Journal of Mammalogy 59 (2), 1978, S. 269–298
- Walter W. Dalquest: Vertebrate fossils from the Blanco local fauna of Texas. Occasional Papers from the Museum Texas Tech University 30, 1975, S. 1–52
- Gary S. Morgan, Paul L. Sealey und Spencer George Lucas: Pliocene and Early Pleistocene (Blancan) vertebrates from the Palomas Formation in the vicinity of Elephant Butte Lake and Caballo, Sierry County, Southwestern New Mexico. New Mexico Museum of Natural History and Science, Bulletin 53, 2011, S. 664–736
- Gary S. Morgan und Spencer George Lucas: Pliocene and Pleistocene vertebrate faunas from the Albuquerque basin, New Mexico. New Mexico Museum of Natural History and Science, Bulletin 16, 2000, S. 217–240
- Walter W. Dalquest, Edward Roth und Frank Judd: The mammal fauna of Schulze Cave, Edwards County, Texas. Bulletin of the Florida State Museum, Biological Sciences 13 (4), 1969, S. 205–276
- John E. Guilday: Prehistoric record of Scalopus from western Pennsylvania. Journal of Mammalogy 42 (1), 1961, S. 117–118
- John E. Guilday: Archaeological evidence of Scalopus aquaticus in the upper Ohio Valley. Journal of Mammalogy 53 (4), 1972, S. 905–907
- Louise E. Ritchie und Joseph J. Nocera: Assessing the Distribution of Eastern Moles (Scalopus aquaticus) in Canada in Relation to Loam Soils and Forest Cover. The American Midland Naturalist 164 (1), 2010, S. 61–73, doi:10.1674/0003-0031-164.1.61
- J. Matson, N. Woodman, I. Castro-Arellano und P. C. de Grammont: Scalopus aquaticus (errata version published in 2017). The IUCN Red List of Threatened Species 2016. e.T41471A115188304 (); zuletzt aufgerufen am 26. April 2021
Weblinks
- Scalopus aquaticus in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: J. Matson, N. Woodman, I. Castro-Arellano & P. C. de Grammont, 2016. Abgerufen am 26. April 2021.