Eigentliche Maulwürfe
Die Eigentlichen Maulwürfe (Talpini) bilden eine Gattungsgruppe (Tribus) innerhalb der Altweltmaulwürfe (Talpinae). Diese Gruppe umfasst über 30 Arten in sechs Gattungen, von denen der Europäische Maulwurf der wohl bekanntester Vertreter ist. Ihr Verbreitungsgebiet erstreckt sich über Europa und Asien, wo die einzelnen Vertreter Wälder und offene Landschaften in Tiefland- und Gebirgslagen besiedeln. Es handelt sich um strikt unterirdisch aktive Tiere. An diese Lebensweise sind sie mit einzelnen Körpermerkmalen angepasst. Hierzu gehören der walzen- bis zylinderförmige Körper, der kurze Hals, die äußerst kräftigen und zu Grabwerkzeugen umgestalteten Vordergliedmaßen sowie die stark zurückentwickelten Augen. Dadurch ist es ihnen möglich, sich in selbst gegrabenen Tunneln und Gängen fortzubewegen. Oberirdisch werden diese Gangsysteme häufig durch Maulwurfshügel angezeigt. Als Nahrung dienen überwiegend Regenwürmer und Insekten. Der Nachwuchs kommt ein- bis zweim,al im Jahr zur Welt. Wissenschaftlich eingeführt wurden die Eigentlichen Maulwürfe bereits zu Beginn des 19. Jahrhunderts, auch wenn sich ihr heutiger systematischer Status erst im Verlauf des 20. Jahrhunderts herausbildete. Stammesgeschichtlich sind die Eigentlichen Maulwürfe bis in das Oligozän zurückverfolgbar. Dadurch reihen sich neben den heutigen Gattungen auch einzelne fossile Formen ein.
| Eigentliche Maulwürfe | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Europäischer Maulwurf (Talpa europaea) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Talpini | ||||||||||||
| Fischer, 1817 |
Merkmale
Habitus

Die Eigentlichen Maulwürfe repräsentieren kleine bis große Vertreter der Maulwürfe. Sie erreichen eine Kopf-Rumpf-Länge von 8 bis 23 cm und ein Gewicht von 20 bis 220 g. Der Schwanz ist üblicherweise kurz und übertrifft in der Regel nicht die Länge der Hinterfüße. Die Tiere sind an eine unterirdische Lebensweise angepasst. Dies zeigt sich in dem walzen- bis zylinderförmigen Körper, dem kurzen Hals und den sehr kräftigen sowie nach außen gedrehten Vordergliedmaßen, die dadurch effektive Grabwerkzeuge darstellen. Sie enden in fünf, mit kräftigen Krallen ausgestatteten Fingern. Die Hinterbeine tragen ebenfalls fünf Zehen, sind aber weniger spezialisiert als die Vordergliedmaßen und deutlich schlanker. Des Weiteren ist die Schnauze zugespitzt, äußere Ohrmuscheln sind nicht erkennbar. Die Augen verbergen sich entweder unter einen Hautfalte oder liegen frei, sind aber in jedem Fall in ihrer Größe reduziert. Das Fell ist meist kurz, die Färbung kann von schwarz über grau bis bräunlich variieren, mitunter treten bei einigen Arten Farbanomalien auf. Der Schwanz weist häufig nur eine spärliche Fellbedeckung auf.[1]
Schädel- und Gebissmerkmale
Der Schädel ist bei den grabenden Maulwürfen relativ uniform gebaut. In der Regel erreicht er seine größte Breite am Hirnschädel und spitzt sich nach vorn zu. Das Rostrumj ist je nach Art kurz und breit oder lang und schmal. Eine auffällige Einschnürung im Bereich der Orbita kommt meist nicht vor. Jochbögen sind ausgebildet, wirken aber sehr schwach. Die Schädeloberfläche ist glatt, auffällige Rippeln oder ein Scheitelkamm sind nicht vorhanden. Das Hinterhaupt ist häufig kräftig gestaltet. Die einzelnen Schädelknochen verwachsen sehr früh miteinander. An der Schädelbasis treten vollständige Paukenblasen auf.[2] Das Mittelohr weist einzelne Variabilitäten bezüglich der Gehörknöchelchen auf. Bei allen Arten der Eigentlichen Maulwürfe sind diese nur locker mit dem Ohr verankert, was eine höhere Beweglichkeit ermöglicht und wahrscheinlich mit der Wahrnehmung von Geräuschen im niedrigen Frequenzbereich im Zusammenhang steht. Der Hammer besitzt einen Kopf aus massivem Knochenmaterial, lediglich bei Weißschwanzmaulwurf (Parascaptor) und beim Kurzgesichtmaulwurf (Scaptochirus) ist dieser stark hypertrophiert.[3][4][5]
| Gattung | Zahnformel | Zahnanzahl |
|---|---|---|
| Euroscaptor, Oreoscaptor, Talpa | 44 | |
| Mogera | 42 | |
| Parascaptor | 42 | |
| Scaptochirus | 40 |




Die einzelnen Gattungen der Eigentlichen Maulwürfe lassen sich hauptsächlich anhand des Gebissaufbaus unterscheiden. Die Anzahl der Zähne variiert zwischen 40 und 44. Allerdings ist die Zahnanzahl innerhalb der Gattungen nicht einheitlich, da bei zahlreichen Arten Oligodontie auftritt. Dadurch können Zähne fehlen, zusätzliche auftreten oder mehrere miteinander verschmolzen sein. In entsprechenden Fällen betrifft dies meist die Prämolaren, von denen der jeweils zweite besonders klein ausgebildet ist. Eine weitere Auffälligkeit der Eigentlichen Maulwürfe findet sich in dem kleinen oberen vorderen Schneidezahn, der den beiden hinteren ähnelt. Im Vergleich dazu verfügen die Neuweltmaulwürfe (Scalopini) und die Desmane (Desmanini) über einen stark vergrößerten oberen vorderen Schneidezahn. Des Weiteren haben die Eigentlichen Maulwürfe einen großen sowie spitzen oberen Eckzahn. Das Merkmal kommt auch beim Langschwanzmaulwurf (Scaptnonychini) vor, fehlt aber bei den Neuweltmaulwürfen. Im Unterkiefer zeigt sich bei den Eigentlichen Maulwürfen der Eckzahn hingegen klein und gleicht den Schneidezähnen, ist also incisiviform, während der erste Prämolar vergrößert und eckzahnähnlich, also caniniform wirkt. Bei den Neuweltmaulwürfen bleibt der untere erste Vormahlzahn klein, er ist aber beim Langschwanzmaulwurf ebenfalls vergrößert. Die übrigen vorderen Prämolaren sind bei den Eigentlichen Maulwürfen meist einspitzig. Die Molaren kennzeichnen sich durch mehrere spitze Haupthöcker, die bei Oberkiefermahlzähnen in einer dilambdodonten (W-förmigen) Anordnung stehen. Eine Eigenheit der Eigentlichen Maulwürfe ist der fehlende Zahnwechsel vom Milch- zum Dauergebiss.[2][6][1]
Skelettmerkmale
| Beckenform | Merkmal | Gattung |
|---|---|---|
| caecoidal | keine Knochenbrücke | Euroscaptor, Talpa |
| europaeoidal | Knochenbrücke über der Öffnung des vierten Kreuzbeinwirbels | Parascaptor, Talpa |
| mogerid | Knochenbrücke über der Öffnung des vierten und fünften Kreuzbeinwirbels | Mogera, Oreoscaptor, Scaptochirus |
Auffallend im Skelettbau der Eigentlichen Maulwürfe ist die variierende Gestaltung des Beckens mit drei grundsätzlichen Formen. Diese unterscheiden sich durch eine auftretende Knochenbrücke, die die Öffnungen des vierten (europaeoidal) beziehungsweise des vierten und fünften (mogerid) Kreuzbeinwirbels überdeckt oder fehlt (ceacoidal). Es handelt sich hier jeweils um Anpassungen an die unterirdische Lebensweise. Beim mogeriden Becken versteift die Knochenbrücke das Becken stärker und bietet so größere Muskelansatzstellen für die hinteren Gliedmaßen. Ähnliches kann für das europaeoidale Becken angenommen werden. Das caecoidale Becken kommt vor allem bei Arten vor, die feuchtere Landschaften besiedeln. In einzelnen Fällen treten Übergangsformen auf, bei denen die Foramen der Kreuzbeinwirbel deutlich verschmälert oder geschlossen sind, so dass wiederum eine größere Ansatzfläche entsteht.[7][8] Als zusätzliche Auffälligkeit ist die Symphyse des Schambeins verknöchert.[1]
Weitere besondere Charakteristika teilen die Eigentlichen Maulwürfe mit anderen grabenden Maulwurfsvertetern. Hierfür können der extrem kurze und an den Gelenkenden stark verbreiterte Oberarmknochen und das ebenfalls kurze und breite Schlüsselbein genannt werden. Ersterer stellt den massivsten Knochen im Bewegungsapparat dar und verfügt über kräftige Rippeln für die Armmuskulatur. Der Gelenkkopf sitzt zumeist tief und wird von den beiden Knochenvorsprüngen Tuberculum majus und Tuberculum minus deutlich überragt. Beim Schlüsselbein wiederum übertrifft die Breite mitunter die Länge um teils bis zum Doppelten. Das untere Ende artikuliert wie bei allen Maulwürfen mit den Knochenvorsprüngen des Oberarmknochens. Die einzelnen Handknochen sind kurz und breit gestaltet und ergeben so eine breite Handfläche. Diese wird seitlich zusätzlich durch ein Sesambein erweitert, das bei den Eigentlichen Maulwürfen die Länge der Handwurzel zuzüglich der Mittelhandknochen erreicht.[9][10][11][12] Das Merkmal ist wenigstens seit dem Oligozän nachweisbar.[13] Die jeweiligen Endphalangen der Fingerstrahlen weisen am vorderen Ende einen tiefen Einschnitt auf.[2][8]
Genetische Merkmale
| Gattung | Chromosomensatz | Fundamentale Anzahl | X-Chromosom | Y-Chromosom |
|---|---|---|---|---|
| Euroscaptor | 2n = 34–38 | FNa = 52–60 | metazentrisch | fleckenartig/metazentrisch |
| Mogera | 2n = 30–36 | FNa = 52–60 | metazentrisch | fleckenartig |
| Oreoscaptor | 2n = 36 | FNa = 52 | metazentrisch | fleckenartig |
| Parascaptor | 2n = 34 | FNa = 58/64 | metazentrisch | acrozentrisch |
| Scaptochirus | 2n = 48 | FNa = 54? | ? | ? |
| Talpa | 2n = 34–38 | FNa = 62–64 | metazentrisch | fleckenartig |
Der diploide Chromosomensatz der Eigentlichen Maulwürfe ist stark variabel und reicht von 2n = 30 bis 2n = 48. Der ursprünglich angedachte Ansatz einer eher konservativen Chromosomenstruktur ließ sich durch die zahlreichen Untersuchungen dadurch nicht bestätigen. Ein größerer Anteil der Eigentlichen Maulwürfe weist jedoch einen Chromosomensatz von 2n = 34 bis 36 auf. Dem gegenüber zeigt die Anzahl der Arme der Autosomen, die als fundamentale Anzahl bezeichnet wird, eine bimodale Verteilung. Häufig sind dabei 52 bis 60 Arme ausgebildet, was sich sowohl bei den variantenreicheren Gattungen wie den Ostasiatischen Maulwürfen (Mogera) wie auch bei den Südostasiatischen Maulwürfen (Euroscaptor) findet. Jedoch weichen die Eurasischen Maulwürfe (Talpa) davon ab, bei denen die fundamentale Anzahl jeweils über 60 liegt. Bei den Eurasischen Maulwürfen überwiegen demnach zweiarmige Autosomen wie meta- bis submeta- und telo- bis subtelozentrische Formen gegenüber den einarmigen wie den acrozentrischen. Bei den Ost- und Südostasiatischen Maulwürfen nimmt hingegen die Anzahl acrozentrischer Autosomen im Vergleich zu den zweiarmigen zu.[14][15][16]
Verbreitung und Lebensraum
Die Eigentlichen Maulwürfe sind ausschließlich in Eurasien verbreitet. Der westliche Teil des Kontinentalgebietes wird dabei nur von den Vertretern der Eurasischen Maulwürfe bewohnt, die von Europa bis nach Vorderasien und teils abgetrennt davon auch in Nordasien vorkommen. Im übrigen Asien, vor allem in Ostasien und Südostasien sind verschiedene Gattungen beheimatet, deren Lebensbereiche sich teilweise überschneiden. Hierzu gehören unter anderem die Südostasiatischen Maulwürfe und die Ostasiatischen Maulwürfe. Erstere sind mit einer hohen Diversität in Indochina zu finden, letzterer treten recht vielgestaltig auf den japanischen Inseln auf. Dort ist auch der Japanische Bergmaulwurf (Oreoscaptor) anzutreffen. Der Weißschwanzmaulwurf wiederum besiedelt größere Teile des südöstlichen und östlichen, der Kurzgesichtmaulwurf weitgehend des östlichen Asiens. Ein tatsächlich sympatrisches Vorkommen ist aber nur in den wenigsten Fällen gegeben, in der Regel liegt bei der Nutzung gleicher Regionen eine parapatrische Verbreitung vor.[17][18] Als Lebensräume können geschlossene und offene Landschaften in Tiefland- oder Gebirgslagen herausgestellt werden, zumeist in Verbindung mit tiefgründigeren Böden und einer hohen Biomasse an Beutetieren.[1]
Lebensweise

Die unterirdisch grabenden Aktivitäten bedingen, dass trotz des großen Formreichtums der Eigentlichen Maulwürfe die Lebensweise der Tiere bis auf wenige Arten nur ungenügend erforscht ist. Sie sind einzelgängerisch und legen Tunnel- und Gangsysteme an. Die Ein- und Ausgänge werden häufig oberirdisch durch einen Hügel an Auswurfmaterial (Maulwurfshügel) angezeigt. In den Gangsystemen finden sich Nestkammern als Rückzugsort und zur Aufzucht des Nachwuchses. Sofern untersucht haben die einzelnen Individuen häufig ein fest umrissenes Territorium. Aktivitäten finden sowohl tagsüber als auch nachts statt. Die Nahrung der Eigentlichen Maulwürfe besteht vorwiegend aus Regenwürmern und verschiedenen Insekten einschließlich deren Larven, gelegentlich vertilgen sie auch kleine Wirbeltiere. Bei manchen Arten ist Vorratsspeicherung belegt. Die Eigentlichen Maulwürfe haben eine hohe Stoffwechselrate und müssen daher große Mengen an Nahrung bis fast zum eigenen Körpergewicht zu sich nehmen. Die Fortpflanzung findet einmal, manchmal auch zweimal jährlich statt. Die Tragzeit währt nur wenige Wochen, ebenso die Aufzucht des Nachwuchses. Die maximale Lebenserwartung beträgt rund vier Jahre. Eine Besonderheit findet sich darin, dass zumindest bei einigen Arten der Eurasischen und der Ostasiatischen Maulwürfe Hermaphroditismus nachgewiesen ist, da die Weibchen Zwitterdrüsen besitzen.[19][1]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[20]
|
Die Eigentlichen Maulwürfe sind eine Tribus aus der Familie der Maulwürfe (Talpidae) und der Ordnung der Insektenfresser (Eulipotyphla). Sie bilden die größte und umfangreichste Gruppe innerhalb der Maulwürfe und vereinen die zumeist grabenden Vertreter der Familie. Die Angehörigen anderer Gattungsgruppen leben mit Ausnahmen wie den Neuweltmaulwürfen (Scalopini) oder dem Sternmull (Condylurini) dagegen nur teilweise unterirdisch, bewegen sich oberirdisch fort oder sind an eine semi-aquatische Lebensweise angepasst.[20] Äußerlich weisen die Eigentlichen Maulwürfe dadurch die generellen Kennzeichen der grabenden Maulwürfe auf, die sich durch die kurzen und kräftigen Ober- beziehungsweise Unterarme und großen, schaufelartigen sowie dauerhaft nach außen gedrehten Hände äußern. Darüber hinaus ist ihnen ein kurzer Schwanz zu eigen, bei einigen Arten findet sich zudem eine Hautfalte, welche die Augen. überdeckt. Molekulargenetische Studien zufolge reicht der Ursprung der Maulwürfe bis in das Mittlere Eozän vor rund 47 Millionen Jahren zurück. Hierbei formten sich die Spitzmausmaulwürfe (Uropsilinae) als ursprünglich Gruppe zuerst heraus. Sie stehen als eigenständige Unterfamilie den Altweltmaulwürfen (Talpinae) gegenüber und bilden so deren Schwestergruppe. Die Eigentlichen Maulwürfe, die wiederum eine Teilgruppe der Altweltmaulwürfe darstellen, trennten sich von den anderen Triben im Oberen Eozän vor rund 34 bis 36 Millionen Jahren ab. Die weitere Aufsplittung der Talpini fand nicht vor dem Mittleren Miozän vor rund 12 Millionen Jahren. Als nächstverwandte Gruppen können die Desmane (Desmanini) und der Sternmull angesehen werden.[21][18][20]
Die Eigentlichen Maulwürfe gliedern sich heute in insgesamt sechs Gattungen:[22][1]
Innere Systematik der Eigentlichen Maulwürfe nach He et al. 2016[20]
|
- Tribus: Eigentliche Maulwürfe (Talpini Fischer, 1817)
- Südostasiatische Maulwürfe (Euroscaptor Miller, 1940); östliches und südöstliches Asien (hauptsächlich Indochina, Malaiische Halbinsel, südliches China), 10 Arten
- Ostasiatische Maulwürfe (Mogera Pomel, 1848); östliches und südöstliches Asien (hauptsächlich Japan, nordöstliches und südliches China sowie Indochina), 9 Arten
- Japanischer Bergmaulwurf (Oreoscaptor Kawada, 2016); östliches Asien (Japan), 1 Art
- Weißschwanzmaulwurf (Parascaptor Gill, 1875); südliches, östliches und südöstliches Asien (nordöstliches Indien, Indochina und südliches China), 1 Art
- Kurzgesichtmaulwurf (Scaptochirus Milne-Edwards, 1867); östliches Asien (nordöstliches China), 1 Art
- Eurasische Maulwürfe (Talpa Linnaeus, 1758); Europa sowie westliches und nördliches Asien, 14 Arten
Neben diesen sechs anerkannten Gattungen wurden noch weitere benannt. Hierzu zählen etwa Asioscalops, Chiroscaptor, Eoscalops und Nesoscaptor, sie sind aber allesamt identisch mit einzelnen der bestehenden Gattungen. So bezieht sich Asioscaptor auf den Sibirischen Maulwurf der Gruppe der Eurasischen Maulwürfe und wird innerhalb dieser auch als Untergattung geführt. Chiroscaptor ist wiederum identisch mit dem Weißschwanzmaulwurf. Dagegen wurde Eoscalops teilweise für den Langnasen-Maulwurf aus der Gattung der Südostasiatischen Maulwürfe verwendet, während Nesoscaptor den Ryukyu-Maulwurf aus der Gattung der Ostasiatischen Maulwürfe meint. Der Japanische Bergmaulwurf indes stand ursprünglich innerhalb der Südostasiatischen Maulwürfe, ist aber nach genetischen Untersuchungen mit den Ostasiatischen Maulwürfe näher verwandt. Er wurde daher im Jahr 2016 in die eigene Gattung Oreoscaptor ausgelagert.[22][1]
Fossil sind noch einzelne weitere Gattungen beschrieben worden:[23][24]
- Geotrypus Pomel, 1848; Oligozän, Unteres Miozän
- Skoczenia Rzebik-Kowalska, 2014; Pliozän, Pleistozän
Forschungsgeschichte

Die wissenschaftliche Bezeichnung Talpini für die Tribus der Eigentlichen Maulwürfe wird auf Johann Fischer von Waldheim aus dem Jahr 1817 zurückgeführt. Nur drei Jahre zuvor hatte Fischer von Waldheim in einer Abhandlung bereits den Namen „Talpinorum“ genutzt. In beiden Fällen verstand Fischer von Waldheim diese als oberhalb der Tribus stehende Klassifizierungseinheit, da die Taxa neben den damals bekannten Maulwürfen zusätzlich die Goldmulle (Chrysochloridae) mit einschlossen, die allerdings nicht näher verwandt sind und in Afrika auftreten.[25][26] In der wissenschaftlichen Literatur wird Fischer von Waldheim daher sowohl mit 1814 als auch mit 1817 als Namensgeber zitiert, was gleichzeitig auch für den Namen der Unterfamilie (Talpinae) und der Familie (Talpidae) gilt. Im Jahr 1825 gab John Edward Gray erstmals die korrekte Bezeichnung Talpidae für die Familie der Maulwürfe an. Innerhalb seiner Talpidae unterschied er mehrere Unterfamilien, die wiederum die Goldmulle, die Spitzmäuse (Soricidae), die Igel (Erinaceidae), die Tenreks (Tenrecidae) und die eigentlichen Spitzhörnchen (Tupaiidae) beinhalteten. Die Maulwürfe beschränkten sich allerdings nicht auf die von Gray so benannten „Talpina“, in die er lediglich die Eurasischen Maulwürfe (Talpa) einordnete, sondern verteilten sich mit dem Sternmull (Condylura), dem Ostamerikanischen Maulwurf (Scalopus) oder den Desmanen (Desmana) auf die „Chrysochlorina“ und die „Soricina“.[27]
In der Folgezeit wurden mehrere ähnliche oder leicht variierende Konzepte vorgestellt, so unter anderem von Auguste Pomel 1848[28] und Paul Gervais 1854,[29] doch vereinte erstmals Wilhelm Peters 1863 alle damals bekannten Maulwürfe in einer übergeordneten Gruppe, nicht ohne jedoch die Goldmulle auszuschließen.[30] Dieser Schritt wurde erst rund fünf Jahre später von St. George Jackson Mivart vollzogen. Mivart differenzierte dabei innerhalb der Talpidae mit den „Talpina“ und den „Myogalina“ zwei Unterfamilien heraus. Erstere beinhaltete neben den Eigentlichen Maulwürfen auch den Sternmull und die Neuweltmaulwürfe (Scalopini), letztere die Desmane und die Japanischen Spitzmulle (Urotrichus).[31] Eine ähnliche Gliederung nutzte später, 1875, auch Theodore Gill, doch verwendete er für die Unterfamilien die heute grammatikalisch korrekteren Formen Talpinae und Myogalinae.[32] Die Gliederung wurde dann in der darauffolgenden Zeit weitgehend übernommen und findet sich unter anderem bei William Henry Flower 1884.[33] Eine deutlich stärkere Aufteilung der Maulwürfe bot George Gaylord Simpson im Jahr 1945 in seiner aufbereiteten generellen Klassifizierung der Säugetiere an. Hier setzte er neben den Talpinae und den Desmaninae (die die Position der Myogalinae einnahmen) zusätzlich noch die Uropsilinae (Spitzmausmaulwürfe), die Scalopinae (Neuweltmaulwürfe) und die Condylurinae (Sternmull) ab. Die Talpinae entsprachen bei Simpson weitgehend den Eigentlichen Maulwürfen in der heutigen Auffassung.[34] Ein tatsächlich auf tribunaler Ebene liegendes Taxon Talpini findet sich dann erstmals bei Leigh Van Valen im Jahr 1967. In seiner neuen systematischen Gliederung der Insektenfresser hatte Van Valen die Talpinae als übergeordnete Gruppe herausgearbeitet, denen er die Talpini, die Scalopini und die Condylurini unterordnete.[6] Der Status der Eigentlichen Maulwürfe als Tribus blieb danach weitgehend stabil, auch wenn die genaue Gliederung der Maulwürfe in der Folgezeit mitunter Schwankungen unterlegen war.[35][36] Erst genetische Untersuchungen aus den 2010er Jahren führten zur Etablierung der heute anerkannten Einteilung der Maulwürfe.[20][1]
Die engere Verwandtschaft der Eigentlichen Maulwürfe innerhalb der Maulwürfe wurde aufgrund der Gebissstruktur schon relativ früh erkannt. Als probates Untergliederungskriterium erwies sich die schwankende Zahnformel, was bereits Gill 1875 vorschlug[32] und von zahlreichen Autoren bis heute akzeptiert wird.[37][2][38][39] Einen anderen Weg ging Ernst Schwarz im Jahr 1948, der die teils starken Variationen in der Zahnanzahl bei einigen Gattungen und Arten der Eigentlichen Maulwürfe zum Anlass nahm, alle bekannten Formen innerhalb der Eurasischen Maulwürfe und damit unter der Gattungsbezeichnung Talpa zu vereinen. Sein Schema befürwortete für den westeurasischen Bereich, also dem klassischen Verbreitungsgebiet der Eurasischen Maulwürfe, mehrere Arten. Dahingegen versammelte er alle Vertreter des ost- und südostasiatischen Raumes in einer Art, die er wiederum in mehrere Unterarten aufteilte.[40] Das Konzept fand teils großen Anklang und wurde vielfach übernommen. Für den ostasiatischen Raum hoben dies spätestens in den 1960er Jahren einige japanische Wissenschaftler wieder auf.[17] Als bedeutend in diesem Zusammenhang erwiesen sich die Arbeiten von Ivo Grulich in den 1970er und 1980er Jahren. Durch anatomische Studien konnte er neben den Unterschieden in der Gebissstruktur und schädelmorphologischen Kriterien auch Abweichungen im Beckenaufbau herausstellen. In Kombination ließen sich diese variierenden Merkmale nach Grulich nicht auf innergenerischer oder innerartlicher Ebene erklären.[7][8] Dadurch wurde das Konzept nach Schwartz in den 1980er und 1990er Jahren weitgehend wieder aufgegeben.[39]
Stammesgeschichte
Die frühesten stammesgeschichtlichen Nachweise der Eigentlichen Maulwürfe reichen bis in das Oligozän zurück. Sie werden zumeist der Gattung Geotrypus zugewiesen. Diese Form war extrem weit verbreitet und ließ sich von West- und Mitteleuropa über Südeuropa bis nach Kleinasien belegen.[23][41] Die ältesten Funde von Geotrypus gehören dem Unteren Oligozän an und sind gut 33 Millionen Jahre alt, sie wurden an einzelnen Fundstellen der Fränkischen Alb entdeckt.[42] An zahlreichen Lokalitäten, wo die Gattung dokumentiert wurde, liegen aber nur wenige Fossilreste vor.[23] Ausnahmen bilden einzelne Teilskelette, unter anderem eines aus einem Basalt-Steinbruch bei Enspel in Rheinland-Pfalz. Dieses gehört dem Oberen Oligozän an und repräsentiert ein wohl nahezu ausgewachsenes Tier, dessen Handskelett unter anderem durch einen „Präpollex“ charakterisiert ist und somit eine ausgewiesene Grabbefähigung anzeigt.[13] Weitere Teilskelette kamen aus der Umgebung von Ulm zu Tage und datieren in das Untere Miozän.[43][44] Aus diesem Zeitraum stammen auch die jüngsten Nachweise der Gattung, so etwa aus einer Spaltenfüllung bei Petersbuch in Bayern. Wissenschaftlich eingeführt wurde die Gattung bereits im Jahr 1848 durch Auguste Pomel, der sich dabei auf einen Unterkiefer von Cournon und einen Oberarmknochen von Chauffours bezog.[28] Beide Stücke stammten aus dem Oberen Oligozän des nördlichen Frankreichs und waren schon acht Jahre zuvor von Henri Marie Ducrotay de Blainville im ersten Band seines umfassenden Werkes Osteographie des mammifères abgebildet, aber von ihm unter Talpa geführt worden.[45] Teilweise bestand Unklarheit über die genaue phylogenetische Stellung, da die Gattung teils mit dem Langschwanzmaulwurf (Scaptonychini), teils auch mit den Neuweltmaulwürfen (Scalopini) in Verbindung gebracht wurde. Besondere Kennzeichen finden sich in dem breiten Oberarmknochen, der aber noch nicht die Robustizität wie bei den Neuweltmaulwürfen oder den späteren Eigentlichen Maulwürfen erreicht, aber ein eindeutiges Kennzeichen grabender Aktivitäten bildet. Der vergrößerte vordere Prämolar, der an einen Eckzahn erinnert, kommt zwar auch beim Langschwanzmaulwurf vor, ist ansonsten aber nur bei den Eigentlichen Maulwürfen zu finden. Diese und weitere Merkmale sprechen somit eher für eine Stellung innerhalb letzterer. Im Laufe ihrer stammesgeschichtlichen Entwicklung erwies sich die Gattung Geotrypus als sehr variantenreich, ihr werden gut ein Dutzend Arten zugewiesen.[23][13][43]
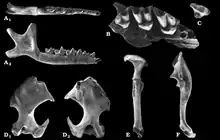
Nahezu zeitgleich mit dem Verschwinden von Geotrypus tritt auch erstmals ein Vertreter der Gattung Talpa in Erscheinung, die vergleichbar zu ihren heutigen Angehörigen auch fossil sehr formenreich gewesen ist und sowohl kleine wie größere Arten einschloss. Mitunter kommt sie gemeinsam mit Geotrypus vor wie aus Ulm oder Petersbuch belegt.[46][44][47][48][49] Anfangs ersetzt Talpa vorerst die kleineren Angehörigen von Geotrypus, später auch die größeren.[23] Das bereits zeitige Auftreten im Unteren Miozän stellt einen frühen Abzweig der Eurasischen Maulwürfe dar, der möglicherweise mit einem verstärkten Vorkommen tiefer grabender Maulwürfe im Zusammenhang steht.[49] Im Oberen Miozän vor rund 8,5 Millionen Jahren verschwindet die Gattung vorerst wieder in Europa. Die nächsten Nachweise konnten dann erst wieder aus dem Pliozän erbracht werden.[50][48][51][52] Talpa erreicht dann eine weit über Europa verteilte Verbreitung und ist von den Gebieten nördlich der Alpen bis auf die Balkanhalbinsel dokumentiert.[53][54][55][56] Etwa im gleichen Zeitraum tritt noch Skoczenia in Erscheinung, eine Form, die über mehrere Gebissreste von verschiedenen Fundstellen in Polen wie Kadzielnia oder die Höhle Zamkowa Dolna beschrieben wurde und sich durch massive Zähne auszeichnet.[24]
Möglicherweise waren die Eigentlichen Maulwürfe bereits im Übergang vom Miozän zum Pliozän im östlichen Asien anwesend, was ein einzelner oberer Prämolar und eine Elle aus Ertemte in der Inneren Mongolei vermuten lassen. Die Funde sind aber keiner speziellen Form zuweisbar.[57] Eindeutigere Belege gehören zeitlich in den weiteren Verlauf des Pliozäns. Hierunter fällt der Erstnachweis der Gattung Scaptochirus. Die frühen Verwandten des Kurzgesichtmaulwurfs sind ebenfalls aus dem nördlichen Ostasien überliefert.[58] Die übrigen heute noch bestehenden Gattungen wie Mogera, Parascaptor, Euroscaptor und Oreoscaptor treten dann im Verlauf des Pleistozäns in Erscheinung. Bis auf Mogera ist der Fossilnachweis aber zumeist spärlich.[59]
Literatur
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4
Einzelnachweise
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- S. U. Stroganov: Morphological characters of the auditory ossicles of recent Talpidae. Journal of Mammalogy 26 (4), 1945, S. 12–42
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Daisuke Koyabu, Misato Hosojima und Hideki Endo: Into the dark: patterns of middle ear adaptations in subterranean eulipotyphlan mammals. Royal Society Open Science 4, 2017, S. 170608, doi:10.1098/rsos.170608
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Ivo Grulich: Zum Bau des Beckens (Pelvis), eines systematisch-taxonomischen Merkmales, bei der Unterfamilie Talpinae. Zoologické Listy 20, 1971, S. 15–28 ()
- Ivo Grulich: Zur Kenntnis der Gattungen Scaptochirus und Parascaptor (Talpini, Mammalia). Folia Zoologica 31, 1982, S. 1–20 ()
- B. Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. American Journal of Anatomy 64, 1939, S. 1–39
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumenting the polydactyly ‚constraint‘: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- Constanze Bickelmann, Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Transcriptional heterochrony in talpid mole autopods. EvoDevo 3, 2012, S. 16, doi:10.1186/2041-9139-3-16
- Achim H. Schwermann und Thomas Martin: A partial skeleton of Geotrypus antiquus (Talpidae, Mammalia) from the Late Oligocene of the Enspel fossillagerstätte in Germany. Paläontologische Zeitschrift 86, 2012, S. 409–439, doi:10.1007/s12542-012-0129-1
- Shin-ichiro Kawada, Nguyen Truong Son und Dang Ngoc Can: Karyological diversity of talpids from Vietnam (Insectivora, Talpidae). In: Dang Ngoc Can, Hideki Endo, Nguyen Truong Son, Tatsuo Oshida, Le Xuan Canh, Dang Huy Phuong, Darrin Peter Lunde, Shin-ichiro Kawada, Akiko Hayashida und Montoki Sasaki (Hrsg.): Checklist of wild mammals of Vietnam. Hanoi, 2008, S. 384–390
- Kai He, Jin-Huan Wang, Wei-Ting Su, Quan Li, Wen-Hui Nie und Xue-Long Jiang: Karyotype of the Gansu mole (Scapanulus oweni): further evidence for karyotypic stability in talpid. Mammal Study 37, 2012, S. 341–348, doi:10.3106/041.037.0408
- Shin-ichiro Kawada, Kohei Kazuma, Haruka Asahina, Takashi Tsuchida, Noriko Tominaga und Motoyoshi Satake: Karyological study of the white tailed mole, Parascaptor leucura, from Myanmar. Bulletin of the National Museum of Nature and Science, Series A 42 (2), 2016, S. 99–104 ()
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture, Hokkaido University 55, 1967, S. 191–265 ()
- Anna A. Bannikova, Elena D. Zemlemerova, Paolo Colangelo, Mustafa Sözen, M. Sevindik, Artem A. Kidov, Ruslan I. Dzuev, Boris Kryštufek und Vladimir S. Lebedev: An underground burst of diversity – a new look at the phylogeny and taxonomy of the genus Talpa Linnaeus, 1758 (Mammalia: Talpidae) as revealed by nuclear and mitochondrial genes. Zoological Journal of the Linnean Society 175, 2015, S. 930–948
- F. David Carmona, Masaharu Motokawa, Masayoshi Tokita, Kimiyuki Tsuchiya, Rafael Jiménez und Marcelo R. Sánchez-Villagra: The Evolution of Female Mole Ovotestes Evidences High Plasticity of Mammalian Gonad Development. Journal of Experimental Zoology 310B (3), 2008, S. 259–266
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Kai He, Akio Shinohara, Xue-Long Jiang und Kevin L. Campbell: Multilocus phylogeny of talpine moles (Talpini, Talpidae, Eulipotyphla) and its implications for systematics. Molecular Phylogenetics and Evolution 70, 2014, S. 513–521, doi:10.1016/j.ympev.2013.10.002
- Shin-ichiro Kawada: Morphological Review of the Japanese Mountain Mole (Eulipotyphla, Talpidae) with the Proposal of a New Genus. Mammal Study 41 (4), 2016, S. 191–205
- Lars W. van den Hoek Ostende: Insectivore faunas from the Lower Miocene of Anatolia – Part 5: Talpidae. Scripta Geologica 122, 2001, S. 1–35
- Barbara Rzebik-Kowalska: Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland. Palaeontologia Electronica 17 (2), 2014, S. 17.2.26A, doi:10.26879/457
- Johann Fischer von Waldheim: Zoognosia tabulis synopticis illustrata. Moskau, 1814, S. X ()
- Johann Fischer von Waldheim: Adversaria zoologica. Mémoires de la Société impériale des naturalistes de Moscou 5, 1817, S. 357–428 ()
- John Edward Gray: An outline of an attempt at the disposition of Mammalia into tribes and families, with a list of the genera apparently appertaining to each tribe. Annals of Philosophy N. S. 10, 1825, S. 337–344 ()
- Auguste Pomel: Etudes sur les carnassiers insectivores (Extrait). Archives des sciences physiques et naturelles, Geneve 1 (9), 1848, S. 159–165 () und 244–251 ()
- Paul Gervais: Histoire naturelle des mammifères, avec l’indication de leurs moeurs, et de leurs rapports avec les arts, le commerce et l’agriculture. Band 1, Paris, 1854, S. 1–420 (S. 239–259) ()
- Wilhelm Peters: Über die Säugethiergattung Selenodon. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1863, S. 1–22 ()
- St. George Jackson Mivart: Notes on the Osteology of the Insectivora. Journal of Anatomy and Physiology 2 (1), 1868, S. 117–154 (S. 150) ()
- Theodore Gill: Synopsis of the insectivorous mammals. Bulletin of the United States Geological and Geographical Survey of the Territories 1, 1874–1875, S. 91–140 (S. 102) ()
- William Henry Flower: Catalogue of the specimens illustrating the osteology and dentition of vertebrated animals, recent and extinct, contained in the museum of the Royal College of Surgeons of England. Part 2. Mammalia other than man. London, 1884, S. 1–779 (S. 641–643) ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 52–53)
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 279–283)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Oldfield Thomas: Description of a new species of mole from China. Annals and Magazine of Natural History 5 (7), 1881, S. 469–471 ()
- Alphonse Milne-Edwards: Sur la classification des taupes de l'ancien continent. Comptes rendus hebdomadaires des séances de l'Académie des sciences 99, 1884, S. 1141–1143 ()
- Shin-ichiro Kawada: The historical notes and taxonomic problems of East Asian moles, Euroscaptor, Parascaptorand Scaptochirus, of continental Asia (Insectivora, Talpidae). Mammal Study 30, 2005, S. S5–S11
- Ernst Schwarz: Revision of the Old‐World Moles of the Genus Talpa Linnaeus. Proceedings of the Zoological Society of London 118 (1), 1948, S. 36–48
- Froukje N. van der Sar, Roos van Glabbeek, Wilma Wessels, Zoran Marković und Hans de Bruijn: Insectivores and marsupials from the upper Oligocene of Banovići (Bosnia and Herzegovina). Journal of Vertebrate Paleontology 37 (6), 2017, S. e1368529, doi:10.1080/02724634.2017.1368529
- Reinhard Ziegler: Moles (Talpidae, Mammalia) from Early Oligocene karstic fissure fillings in South Germany. Geobios 45 (5), 2012, S. 501–513, doi:10.1016/j.geobios.2011.11.017
- Achim H. Schwermann und Richard S. Thompson: Extraordinarily preserved talpids (Mammalia, Lipotyphla) and the evolution of fossoriality. Journal of Vertebrate Paleontology 35 (4), 2015, S. e934828, doi:10.1080/02724634.2014.934828
- Reinhard Ziegler: Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Paläontologie Serie B 167, 1990, S. 1–80
- Henri Marie Ducrotay de Blainville : Osteographie des mammifères insectivores (Talpa, Sorex et Erinaceus L.). In: Ostéographie, ou, Description iconographique comparée du squelette et du système dentaire des mammifères. Tome Premiere. Paris, 1840, S. 1–115 ( und )
- Reinhard Ziegler: Talpiden (Mammalia, Insectivora) aus dem Orleanium und Astaracium Bayerns. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 25, 1985, S. 131–175
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- Reinhard Ziegler: Miocene Insectivores from Austria and Germany - An Overview. Beiträge zur Paläontologie 30, 2006, S. 481–494
- Johannes Klietmann, Doris Nagel, Michael Rummel und Lars W. van den Hoek Ostende: A gap in digging: the Talpidae of Petersbuch 28 (Germany, Early Miocene). Paläontologische Zeitschrift 89, 2015, S. 563–592, doi:10.1007/s12542-014-0228-2
- Thomas Dahlmann: Die Kleinsäuger der unter-pliozänen Fundstelle Wölfersheim in der Wetterau (Mammalia: Lipotyphla, Chiroptera, Rodentia). Courier Forschungsinstitut Senckenberg 227, 2001, S. 1–129 (S. 18–21)
- P. Colangelo, A. A. Bannikova, B. Kryštufek, V. S. Lebedev, F. Annesi, E. Capanna und A. Loy: Molecular systematics and evolutionary biogeography of the genus Talpa (Soricomorpha: Talpidae). Molecular Phylogenetics and Evolution 55, 2010, S. 372–380
- Gabriele Sansalone, Tassos Kotsakis und Paolo Piras: Talpa fossilis or Talpa europaea? Using geometric morphometrics and allometric trajectories of humeral moles remains from Hungary to answer a taxonomic debate. Palaeontologia Electronica 18 (2), 2015, S. 42A ()
- Thomas Dahlmann und Gerhard Storch: Eine pliozäne (ober-ruscinische) Kleinsäugerfauna aus Gundersheim, Rheinhessen. 2. Insektenfresser: Mammalia, Lipotyphla. Senckenbergiana lethaea 76 (1/2), 1996, S. 181–191
- Barbara Rzebik-Kowalska: The Pliocene and Early Pleistocene Lipotyphla (Insectivora, Mammalia) from Romania. Acta zoologica cracoviensia 45 (2), 2002, S. 251–281
- Barbara Rzebik-Kowalska: Insectivora (Mammalia) from the Early and early Middle Pleistocene of Betfia in Romania. II. Erinaceidae Bonaparte, 1838 and Talpidae Gray, 1825. Acta zoologica cracoviensia 43 (1–2), 2000, S. 55–77
- Vasil V. Popov: Late Pliocene Erinaceidae and Talpidae (Mammalia: Insectivora) from Varshets (North Bulgaria). Acta zoologica cracoviensia 47 (1–2), 2004, S. 61–80
- Gerhard Storch und Z. Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Lawrence J. Flynn Und Wen-Yu Wu: The Lipotyphla of Yushe Basin. In: Lawrence J. Flynn und Wen-Yu Wu (Hrsg.): Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals, Volume II: Small Mammal Fossils of Yushe Basin, Vertebrate Paleobiology and Paleoanthropology. Springer Science+Business Media B.V., 2017, S. 11–26
- Yoshinari Kawamura: Quaternary Rodent Faunas in the Japanese Islands (Part 1). Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy 53 (1/2), 1988, S. 31–348 ()