Y-Chromosom
Das Y-Chromosom ist ein Geschlechtschromosom (Gonosom). Es bewirkt die Ausbildung des männlichen Phänotyps.
Bei vielen Arten wird das Geschlecht eines Individuums durch eine chromosomale Geschlechtsbestimmung festgelegt. Wenn in weiblichen Individuen zweimal das gleiche Chromosom (homozygot; XX) und in männlichen Individuen zwei unterschiedliche Chromosomen (hemizygot; XY) vorkommen, heißen diese per Definition X-Chromosomen und Y-Chromosomen.
Das XX/XY-System der Geschlechtsdetermination kommt bei Säugetieren (Theria, d. h. Beuteltiere und Höhere Säugetiere (Eutheria), nicht aber Eierlegende Säugetiere), einigen Insektenarten und noch einigen anderen Tiergruppen vor (siehe Geschlechtschromosom). Bei manchen Lebewesen wie den Vögeln besitzen dagegen männliche Individuen zwei gleiche Z-Chromosomen und weibliche je ein W- und ein Z-Chromosom.
Theorie der Entstehung des Y-Chromosoms bei Säugetieren
Zum Y-Chromosom existiert im diploiden Chromosomensatz kein vollkommen homologes Chromosom. Es hat nur ein Drittel der Größe des X-Chromosoms und kann nur auf fünf Prozent seiner Länge (in den Telomer-nahen pseudoautosomalen Regionen) mit dem X-Chromosom rekombinieren, obwohl es noch zahlreiche Gene des allgemeinen Stoffwechsels aufweist, die auch auf dem X-Chromosom zu finden sind. Viele Gene des Y-Chromosoms haben aber kein Gegenstück auf dem X-Chromosom. Es sind vor allem Gene der Spermienproduktion. Aufgrund der Kenntnis der Basensequenz des menschlichen Genoms durch das Humangenomprojekt lässt sich ein Entwicklungsweg dieses Chromosoms konstruieren.
Die gängigste Theorie zur Entstehung des Y-Chromosoms bei Säugetieren besagt, dass die beiden Gonosomen X und Y das Ergebnis von Mutationen in einem gemeinsamen Vorläuferchromosom (das jeweils analog den Autosomen diploid vorlag) sind.[1][2] Dieses enthielt die Gene, die zur Ausbildung beider Geschlechter notwendig sind; für die Differenzierung zum männlichen und weiblichen Geschlecht waren exogene Einflüsse wie die Temperatur verantwortlich. Der entscheidende Schritt seien Mutationen gewesen, die einem dieser Chromosomen Gene einbrachten, die eindeutig für die Entwicklung hin zum männlichen Geschlecht verantwortlich gemacht werden können. Gleichzeitig müssen diese Mutationen dazu geführt haben, dass sich die beiden Geschlechtschromosomen in ihrer Sequenz so stark unterschieden, dass eine Rekombination zwischen ihnen ausgeschlossen wurde, so dass das neue Männlichkeitsgen nicht auf das unmutierte Geschlechtschromosom verschoben werden konnte. Diese „große“ Mutation soll eine Inversion auf dem langen Arm des Y-Chromosoms gewesen sein. Es entstand das SRY-Gen (sex determinating region on Y chromosome), das für den testis determinating factor (TDF) codiert; durch die Inversion wurde die Paarung mit dem vormals homologen Bereich auf dem nicht-mutierten Chromosom verhindert.[3]
Das Geschlecht eines Individuums auf diese Weise zufällig zu bestimmen, hat den Effekt der relativen Ausgeglichenheit der Geschlechterverteilung, während zuvor Populationen mit einem stark verschobenen Geschlechterverhältnis möglich waren.
Im weiteren Verlauf habe das Y-Chromosom immer wieder Gene verloren,[4] die nicht mit der Entwicklung des männlichen Geschlechts assoziiert waren, während sich andererseits Gene, die beispielsweise Bedeutung für die Fruchtbarkeit der Männchen hatten, mehr und mehr auf dem Y-Chromosom versammelten. Allerdings existieren nach wie vor auf X und Y homologe Gene, welche letztlich auch die Basis der gesamten Theorie des gemeinsamen Vorläuferchromosoms sind.
Mit dem Verlust autosomaler Gene allein auf dem Y-Chromosom ging allerdings einher, dass zwischen männlichen und weiblichen Mitgliedern einer Spezies beträchtliche Unterschiede in der Aktivität jener Gene entstanden, die nun nur noch auf dem X-Chromosom vorlagen (Frauen haben die doppelte Gen-Dosis und somit theoretisch die doppelte Genaktivität). Da aber jede Frau eines dieser X-Chromosomen auch wieder an einen Sohn weitergeben können muss, musste eine Lösung des Dosisproblems gefunden werden, die für beide Geschlechter gleichermaßen funktioniert.
Zum einen finden sich in den Theria (d. h. Beuteltieren und Höheren Säugetieren) viele Gene des ehemaligen gemeinsamen Vorläuferchromosoms auf Autosomen – so besitzen männliche und weibliche Individuen jeweils die gleiche (diploide) Gendosis. Zum anderen wird in jeder weiblichen Zelle eins der beiden X-Chromosomen inaktiviert (siehe Barr-Körperchen). Allerdings betrifft diese Deaktivierung offenbar nicht alle Gene auf dem betreffenden X-Chromosom, so dass Frauen zum Teil doch eine höhere Genaktivität aufweisen, was gern auch populärwissenschaftlich zur Erklärung einiger geschlechtsspezifischer Unterschiede (wie der höheren Sprachbegabung und dem ausgeprägteren Sozialverhalten von Frauen im Vergleich zu Männern) herangezogen wird.[5]
Schließlich hat die Evolution vom gemeinsamen Vorläuferchromosom hin zum X-Y-System auch Nachteile für die männlichen Individuen einer Spezies. Denn X-chromosomal-rezessive Gendefekte, die bei Frauen meist durch die zufällige Inaktivierung eines X-Chromosoms wenig ins Gewicht fallen, können beim männlichen Genotyp nicht kompensiert werden. Ein Beispiel: Eine Mutation auf dem X-Chromosom führt zur Rot-Grün-Blindheit. Frauen besitzen durch die zufällige Inaktivierung eines X-Chromosoms also rot-grün-empfindliche und -unempfindliche Rezeptorzellen in der Netzhaut. Söhne dieser Frauen (Konduktorinnen) haben das 50-prozentige Risiko, das defekte X-Chromosom der Mutter zu erben, und können diesen Defekt dann auch nicht kompensieren. So treten heterozygote Mütter bei X-chromosomal rezessiv vererbten Erkrankungen immer als klinisch nicht oder nur milde betroffene Überträgerinnen auf.
Veränderungen im Laufe der Zeit
Es wird angenommen, dass das Y-Chromosom ein Homolog zum X-Chromosom war, also dieselbe Struktur und dieselben Genorte besaß.
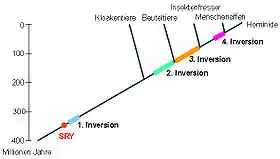
Möglicherweise vor 350 Millionen Jahren entstand auf dem längeren Arm des einen X-Chromosoms aus dem Gen SOX3 der Vorläufer des geschlechtsbestimmendes SRY-Gens (Sex determining region of Y). SRY codiert ein Signalprotein, das verschiedene Gene aktiviert, die die Entwicklung der Hoden im Embryo bewirken. Dieses neue Gen hat vermutlich die Entwicklungsmöglichkeiten eines Individuums zum männlichen Geschlecht stärker gefördert, als es zuvor möglich war. Bei einigen Sauropsida ("Reptilien") wird die Geschlechtsentwicklung durch Umwelteinflüsse, wie die Umgebungstemperatur, beeinflusst, sie besitzen kein SRY-Gen. Da bereits die Kloakentiere – und alle anderen Säugetiere – dieses Gen aufweisen, entstand dieses Gen möglicherweise zu dieser Zeit der Abspaltung der frühen Säugetiere von den Reptilien.
Vor 320 bis 240 Millionen Jahren fand im längeren Arm des Y-Chromosoms eine Inversion statt, die fast den ganzen Arm betraf. Dadurch konnten in diesem Abschnitt zwischen X- und Y-Chromosom keine Rekombinationen mehr stattfinden. Dies führte zu stärkeren Abweichungen von den homologen Genorten im X-Chromosom oder sogar zu Genverlusten (Deletionen). Da im weiblichen Geschlecht die volle Rekombinationsfähigkeit zwischen den homologen X-Chromosomen und damit die Reparaturfähigkeit erhalten blieb, kam es auf dem X-Chromosom nicht zu Genverlusten.
Es werden drei weitere Inversionen vor 170 bis 130, 130 bis 80 und 50 bis 30 Millionen Jahren im kurzen Arm des Y-Chromosoms angenommen, die die Rekombinationsfähigkeit weiter einschränkten und Verluste förderten, wodurch das Y-Chromosom weiter verkürzt wurde.
Zu einem nicht rekonstruierbaren Zeitpunkt wurde das SRY-Gen vom langen in den kurzen Arm des Y-Chromosoms verlagert.
Nicht alle Säuger (Mammalia) haben das X-Y-System der Geschlechtschromosomen. Bei den basalen Eierlegenden Säugetieren (Monotremata) findet man ein abweichendes System, so etwa beim Schnabeltier.
Kompensationsmechanismen
Einige Gene des Grundstoffwechsels auf dem Y-Chromosom sind gegenüber den entsprechenden Genorten des X-Chromosoms sogar in den invertierten Abschnitten kaum verändert. Hätten bei diesen Genen stärkere Veränderungen stattgefunden, wäre die Überlebensfähigkeit der männlichen Genträger so herabgesetzt worden, dass diese Mutationen durch Selektion ausgemerzt worden wären.
Mit dem Verlust vieler Gene auf dem Y-Chromosom hätten manche Gene im männlichen Geschlecht nur die halbe Aktivität. Im Laufe der Evolution kam es zu einer Kompensation: Um die verminderte Aktivität aufgrund der fehlenden zweiten X-Gene im männlichen Geschlecht auszugleichen, wurde die Aktivität der entsprechenden Gene auf dem X-Chromosom verdoppelt. Dadurch hätte aber im weiblichen Geschlecht eine zu hohe Genaktivität vorgelegen. Dies wurde dadurch kompensiert, dass die Gene auf einem der beiden X-Chromosomen im Laufe der Entwicklung inaktiviert werden.
Bei der Fruchtfliege Drosophila melanogaster verdoppeln die Männchen die Aktivität der Gene auf dem X-Chromosom, die keine Entsprechung auf dem Y-Chromosom haben.
Ansammlung von Fertilitätsgenen
Neben dem Verlust von Genen kann aber auch ein Erwerb vor allem der Fertilitäts-Gene beim Y-Chromosom festgestellt werden:
- Zum einen entstanden durch Mutationen auf dem Y-Chromosom neue Gene.
- Zum anderen sammelten sich Gene von anderen Chromosomen durch Translokation auf dem Y-Chromosom.
Es ist noch nicht geklärt, welcher Evolutionsmechanismus zu dieser Ansammlung führte. Möglicherweise müssen sich die Fertilitätsgene auf dem Y-Chromosom sammeln, da die Weibchen ohne Schaden zu erleiden auf diese nur für die Reifung der Spermien wichtigen Gene verzichten können.
Stabilität der Fertilitätsgene
Eigentlich müssten die Fertilitätsgene, die nur auf dem Y-Chromosom vorkommen, aufgrund der fehlenden Rekombinationsmöglichkeit auch zugrunde gehen. Sie liegen jedoch auf einem Chromosom in mehreren Kopien vor, die einzelne Verluste kompensieren können.
Bekannte Gene auf dem Y-Chromosom
Das Y-Chromosom enthält unter anderem folgende Gene:
- AMELY Amelogenin, Y Isoform
- RPS4Y1 40S ribosomales Protein S4, Y Isoform1
- RPS4Y2 40S ribosomales Protein S4, Y Isoform2
- AZF1 Azoospermia Factor 1
- BPY2 Testis-specific basic protein Y 2
- DAZ1 Deleted in azoospermia protein 1
- DAZ2 Deleted in azoospermia protein 2
- DDX3Y DEAD-box helicase 3, Y-linked
- PRKY Serin/Threonin-Proteinkinase PRKY
- RBMY1A1 RNA binding motif protein, Y-linked, family 1, member A1
- SRY sex-determining region of Y
- TSPY Testis-specific Y-encoded protein 1
- USP9Y Ubiquitin specific peptidase 9, Y-linked
- UTY
- ZFY Zinc finger Y-chromosomal protein
Von den Genen des Y-Chromosoms sind in Mäusen zwei Gene essentiell, SRY und Eif2s3y.[6] Werden diese beiden Gene auf anderen Chromosomen untergebracht, können (männliche) Mäuse ohne Y-Chromosom erzeugt werden.[6]
Synonyme
Das Y-Chromosom wird in der Bioinformatik auch als 24. Chromosom bezeichnet, wenn bestimmte Datenbanken nur die Eingabe von Zahlen zulassen.[7]
Siehe auch
Einzelnachweise
- S. Sun, J. Heitman: Should Y stay or should Y go: the evolution of non-recombining sex chromosomes. In: BioEssays : news and reviews in molecular, cellular and developmental biology. Band 34, Nummer 11, November 2012, S. 938–942, ISSN 1521-1878. doi:10.1002/bies.201200064. PMID 22948853. PMC 3700811 (freier Volltext).
- M. A. Jobling: The impact of recent events on human genetic diversity. In: Philosophical transactions of the Royal Society of London. Series B, Biological sciences. Band 367, Nummer 1590, März 2012, S. 793–799, ISSN 1471-2970. doi:10.1098/rstb.2011.0297. PMID 22312046. PMC 3267116 (freier Volltext).
- P. Manolakou, G. Lavranos, R. Angelopoulou: Molecular patterns of sex determination in the animal kingdom: a comparative study of the biology of reproduction. In: Reprod Biol Endocrinol. 4, 13. November 2006, S. 59. PMID 17101057
- J. H. Malone, B. Oliver: The sex chromosome that refused to die. In: BioEssays : news and reviews in molecular, cellular and developmental biology. Band 30, Nummer 5, Mai 2008, S. 409–411, ISSN 1521-1878. doi:10.1002/bies.20752. PMID 18404685. PMC 2696266 (freier Volltext).
- C. M. Disteche: Dosage compensation of the sex chromosomes. In: Annual review of genetics. Band 46, 2012, S. 537–560, ISSN 1545-2948. doi:10.1146/annurev-genet-110711-155454. PMID 22974302. PMC 3767307 (freier Volltext).
- Y. Yamauchi, J. M. Riel, V. A. Ruthig, E. A. Ortega, M. J. Mitchell, M. A. Ward: Two genes substitute for the mouse Y chromosome for spermatogenesis and reproduction. In: Science. Band 351, Nummer 6272, Januar 2016, S. 514–516, doi:10.1126/science.aad1795. PMID 26823431.
- Homo sapiens chromosome Y, GRCh38.p13 Primary Assembly. 29. Mai 2020 (nih.gov [abgerufen am 5. Juli 2020]).
Weblinks
- The Y Chromosome Consortium (YCC) (englisch)
- Human Y Chromosome information (Oak Ridge National Laboratory) (englisch)
| Evolutionsbaum Haplogruppen Y-chromosomale DNA (Y-DNA) | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adam des Y-Chromosoms | ||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0’1'2’3'4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1’2'3’4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A1 | A2’3'4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2’3 | A4=BCDEF | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2 | A3 | B | CT | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| G | IJK | H | ||||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| G1 | G2 | IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | L | K(xLT) | T | ||||||||||||||||||||||||||||||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||||||||||||||||||||||||||
| I1 | I2 | J1 | J2 | M | NO | P | S | |||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| R1 | R2 | |||||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| R1a | R1b | |||||||||||||||||||||||||||||||||||||||||||||||||
Chromosom 1 | Chromosom 2 | Chromosom 3 | Chromosom 4 | Chromosom 5 | Chromosom 6 | Chromosom 7 | Chromosom 8 | Chromosom 9 | Chromosom 10 | Chromosom 11 | Chromosom 12 | Chromosom 13 | Chromosom 14 | Chromosom 15 | Chromosom 16 | Chromosom 17 | Chromosom 18 | Chromosom 19 | Chromosom 20 | Chromosom 21 | Chromosom 22 | X-Chromosom | Y-Chromosom