Neuweltmaulwürfe
Die Neuweltmaulwürfe (Scalopini) bilden eine Gattungsgruppe (Tribus) innerhalb der Altweltmaulwürfe (Talpinae). Sie besteht aus insgesamt neun Arten, aufgeteilt auf fünf Gattungen. Neben den Eigentlichen Maulwürfen (Talpini) stellen sie die bekanntesten Vertreter der grabenden Maulwürfe (Talpidae) dar. Der Großteil der Vertreter ist in Nordamerika verbreitet, worauf sich der deutsche Trivialname bezieht. Allerdings sind auch zwei Formen aus Ostasien belegt. Die Tiere ähneln ihren entfernten Verwandten aus der Gruppe der Eigentlichen Maulwürfe. Wie diese sind sie an eine grabende Lebensweise angepasst und besitzen einen walzenförmigen Körper, einen kurzen Hals und grabschaufelartige Vorderbeine. Der Schwanz der Neuweltmaulwürfe ist jedoch deutlich länger. Als Lebensräume nutzen sie geschlossene und offene Landschaften von Tiefland- bis in höhere Gebirgslagen. Der spezielle Körperbau ermöglicht es den Tieren, unterirdische Tunnel und Gänge anzulegen. Die Gangsysteme werden oberflächig durch Maulwurfshügel angezeigt. Die Nahrung setzt sich aus Regenwürmern und Insekten zusammen, teilweise gehören auch pflanzliche Bestandteile dazu. Der Nachwuchs kommt einmal jährlich zur Welt und wird nur für kurze Zeit gesäugt. Als Ordnungseinheit wurden die Neuweltmaulwürfe bereits 1875 definiert. Ihre genaue systematische Stellung variierte im Laufe der Zeit. Fossil traten die Neuweltmaulwürfe erstmals im Übergang vom Oligozän zum Miozän in Erscheinung. Der Fossilbericht ist relativ umfangreich und schließt mehrere ausgestorbene Gattungen ein.
| Neuweltmaulwürfe | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Ostamerikanischer Maulwurf (Scalopus aquaticus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Scalopini | ||||||||||||
| Gill, 1875 |
Merkmale
Habitus

Die Neuweltmaulwürfe repräsentieren kleine bis große Vertreter der Maulwürfe. Die Kopf-Rumpf-Länge variiert von 8,0 bis 17,9 cm, der Schwanz wird zwischen 2,1 und 5,1 cm lang. Das Gewicht reicht dementsprechend von 20 bis 171 g. Die beiden bekannten ostasiatischen Vertreter sind deutlich kleiner als die übrigen nordamerikanischen. Allgemein liegen die Tiere damit aber in der Variationsbreite der Eigentlichen Maulwürfe (Talpini). Im Unterschied zu diesen wird der Schwanz bei den Neuweltmaulwürfen deutlich länger und übertrifft in jedem Fall die Länge des Hinterfußes. Im Körperbau ähneln sich die Individuen beider Gruppen, was auf die Anpassung an eine grabende Lebensweise zurückzuführen ist. Dadurch wirkt der Rumpf walzenförmig, der Hals ist kurz und die die Vorderfüße entsprechen Grabwerkzeugen. Die Handflächen sind äußerst breit und dauerhaft nach außen gedreht, die Hinterfüße hingegen schlank. Sowohl Hände und Füße weisen jeweils fünf Strahlen auf, die Krallen tragen, wobei jene der Hände deutlich kräftiger sind. Als weitere Anpassungen an eine unterirdische Lebensweise sind die Augen stark reduziert und bleiben im Fell verborgen. Äußere Ohrmuscheln lassen sich nicht erkennen. Die Schnauze endet meistens spitz. Das Körperfell zeichnet sich durch eine dunkle bräunliche, dunkelgraue bis schwarze Farbgebung aus. Gelegentlich kommen hellere Flecken vor. Der Schwanz ist bei einigen Vertretern der Neuweltmaulwürfe dicht behaart.[1][2][3]
Schädel- und Gebissmerkmale
Der Schädel ist bei den grabenden Maulwürfen in der Regel recht einheitlich gebaut. Zumeist besitzt er eine konische Form mit der breitesten Stelle im Bereich des Hirnschädels. Letzterer erhebt sich etwas über die Region des Rostrums. Die Oberfläche ist glatt, auffällige Knochenmarken in Form eines Scheitel- oder Hinterhauptskammes sind nicht ausgebildet. Die Jochbögen schließen vollständig, sind aber weitgehend grazil gebaut. Bei mehreren Arten überragt der Mittelkieferknochen deutlich das Nasenbein.[4][5][6] Das Mittelohr zeichnet sich durch einzelne Variationen aus. Die Gehörknöchelchen sind beim Ostamerikanischen Maulwurf (Scalopus) und bei den Westamerikanischen Maulwürfen (Scapanus) nur locker mit dem Ohr verankert, was eine höhere Beweglichkeit ermöglicht. Wahrscheinlich steht dies mit der Wahrnehmung von Geräuschen im niedrigen Frequenzbereich im Zusammenhang. Eine Ausnahme bildet hier der Haarschwanzmaulwurf (Parascalops), der keine derartige Anpassung besitzt. Er ist allerdings auch weniger stark an das Leben im Untergrund angepasst als die übrigen Vertreter der Gruppe. Bei keiner Form kommen stark hypertrophierte Knöcheln vor, wie es etwa von einigen Eigentlichen Maulwürfen berichtet wird. Dennoch ist der Kopf des Hammers beim Ostamerikanischen Maulwurf recht groß und kugelig.[7][8][9]
| Gattung | Zahnformel | Zahnanzahl |
|---|---|---|
| Alpiscaptulus, Parascalops, Scapanus* | 44 | |
| Scalopus | 36 | |
| Scapanulus | 36 |



Ähnlich zu anderen größeren Gruppen der Maulwürfe bestehen auch bei den Neuweltmaulwürfen zwischen den einzelnen Arten und Gattungen Unterschiede im Gebissaufbau. Die Zahnanzahl variiert zwischen 36 und 44. Reduktionen im Gebiss betreffen zumeist die Prämolaren oder den vorderen Gebissabschnitt. Bei einzelnen Arten ist auch Oligodontie belegt, so dass entweder reguläre Zähne fehlen oder überzählige ausgebildet sind. Auch dies wirkt sich zum Großteil bei den Vormahlzähnen aus. Als Unterscheidungsmerkmal zu den Eigentlichen Maulwürfen kann der bei den Neuweltmaulwürfen deutlich vergrößerte obere vordere Schneidezahn herangezogen werden. Das Merkmal teilen sie mit den Desmanen (Desmanini). Dem gegenüber ist der obere Eckzahn der Neuweltmaulwürfe eher klein, bei den Eigentlichen Maulwürfen wiederum sehr groß. Im unteren Gebiss bleibt der vordere Prämolar relativ klein oder ist nicht ausgebildet. Die Eigentlichen Maulwürfe haben hier einen vergrößerten Zahn. Die Molaren verfügen über mehrere Zahnhöckerchen auf der Kauoberfläche. Bei den Oberkieferzähnen sind diese dilambdodont angeordnet, das heißt, sie werden durch eine W-förmige Scherleiste miteinander verbunden. Ebenfalls als Unterschied zu den Eigentlichen Maulwürfen findet bei den Neuweltmaulwürfen ein Zahnwechsel vom Milch- zum Dauergebiss statt.[4][10][11][2]
Skelettmerkmale
Besonderheiten im Skelettbau finden sich unter anderem in der verknöcherten Symphyse des Schambeins am Beckens. Das Schlüsselbein ist wie bei den Eigentlichen Maulwürfen massiv und kurz. Ebenso hat der Oberarmknochen als Anpassung an die unterirdische Lebensweise eine deutliche Überprägung durchlaufen. Er bildet den massivsten Knochen mit breit ausladenden Enden. Der Gelenkkopf sitzt nicht oben, sondern ist seitlich nach unten verlagert. Ihn überragen die beiden Knochenvorsprünge Tuberculum majus und Tuberculum minus deutlich. Am oberen Ende läuft eine markante Knochenrippe quer über den breiten Schaft, die als scalopine ridge bezeichnet wird und bei den Eigentlichen Maulwürfen nur marginal ausgebildet ist.[12][13][14] Das Handskelett besteht als Anpassung an die grabende Lebensweise aus kurzen und breiten Einzelelementen. Ähnlich den Eigentlichen Maulwürfen wird die Handfläche seitlich durch ein zusätzliches Sesambein, der sogenannte Präpollex („Vordaumen“), erweitert. Der Präpollex ist sehr ausgedehnt und erreicht die Länge der Handwurzel einschließlich der Mittelhandknochen,[13][15][16][17] lediglich beim Gansu-Maulwurf (Scapanulus) ist er etwas kürzer. Bereits bei den frühesten Neuweltmaulwürfen aus dem Übergang vom Oligozän zum Miozän lässt sich das Sesambein nachweisen.[18]
Genetische Merkmale
| Gattung | Chromosomensatz | Fundamentale Anzahl | X-Chromosom | Y-Chromosom |
|---|---|---|---|---|
| Alpiscaptulus | ? | ? | ? | ? |
| Parascalops | 2n = 34 | FNa = 56 | metazentrisch | fleckenförmig |
| Scalopus | 2n = 34 | FNa = 64 | metazentrisch | fleckenförmig |
| Scapanulus | 2n = 34 | FNa = 64 | metazentrisch | fleckenförmig |
| Scapanus | 2n = 34 | FNa = 60 | ? | ? |
Alle bisher untersuchten Neuweltmaulwürfe besitzen einen diploiden Chromosomensatz von 2n = 34. Die fundamentale Nummer, also die Anzahl der Arme der Autosomen, variiert zwischen 56 und 64. Im Vergleich zu den Eurasischen Maulwürfen ist die Chromosomenanzahl eher konservativ, bei letzteren variiert sie zwischen 2n = 30 bis 48. Das X-Chromosom ist in der Regel meta- bis submetazentrisch, dass Y-Chromosom klein und fleckenförmig. Da eine übereinstimmende Chromosomenanzahl von 2n = 34 nicht nur in den beiden genannten Linien, sondern auch bei anderen Maulwurfsgruppen vorkommt, wird angenommen, dass es sich hierbei um den ursprünglichen Satz handelt, der dann im Laufe der Stammesgeschichte mehrfach abgewandelt wurde.[19][20][21]
Verbreitung und Lebensraum
Die Neuweltmaulwürfe sind, abweichend von ihrem deutschsprachigen Trivialnamen, in der Neuen Welt und in der Alten Welt verbreitet. Der größere Teil der Arten kommt heute in Nordamerika vor. Hierbei sind die Westamerikanischen Maulwürfe (Scapanus) als formenreichste Gruppe entlang der Pazifikküste vom südlichen Kanada über die USA bis in das nördlichste Mexiko anzutreffen. Der Osten Nordamerikas hingegfen wird sowohl vom Ostamerikanischen Maulwurf (Scalopus) wie auch vom Haarschwanzmaulwurf (Parascalops) bewohnt. Ersterer hat eine deutlich größere Region erschlossen, sie reicht ebenfalls vom Süden Kanadas bis in den Norden Mexikos. Letztere lebt hauptsächlich im nordöstlichen und östlichen Teil der USA. Die beiden derzeit bekannten altweltlichen Vertreter der Neuweltmaulwürfe sind auf Ostasien beschränkt. Der Gansu-Maulwurf (Scapanulus) besiedelt Teile des zentralen Chinas, die Gattung Alpiscaptulus ist lediglich von einzelnen Fundpunkten im Südwesten des Landes bekannt. Häufig werden beide Formen als Reliktarten angesehen, die eventuell auf einen eurasischen Ursprung der Neuweltmaulwürfe hindeuten.[22][23][3] Die Tiere nutzen verschiedenen Landschaftsräume, die von Wäldern bis hin zu Offenland reichen, teilweise auch mit trockenen Bedingungen verbunden. Diese verteilen sich von Tieflandgebieten bis in höhere Gebirgslagen. Häufig werden tiefgründige und feuchte Böden bevorzugt, rein sandiger und steiniger Untergrund jedoch gemieden.[2][3]
Lebensweise

Die Lebensweise der Neuweltmaulwürfe ist in einzelnen Aspekten relativ gut untersucht. Dies trifft aber hauptsächlich auf die nordamerikanischen Vertreter zu, während für die ostasiatischen kaum Daten vorliegen. Die Tiere leben ausnahmslos unterirdisch und graben Gänge und Tunnelsysteme. Diese dienen ihnen als Jagdrevier und als Schlupfwinkel. Meistens verlaufen die Gänge knapp unter der Erdoberfläche, was oberirdisch durch kleine Rippeln erkennbar ist. In kühleren Jahreszeiten werden auch weiter in den Untergrund reichende Tunnel genutzt. Als sichtbares Zeichen dieser Aktivitäten in tieferen Ebenen entstehen häufig charakteristische Auswurfhügel (Maulwurfshügel). In der Regel befindet sich dort auch die Nestkammer, die mit Pflanzen ausgestattet wird. Bei einigen Arten ist als Anpassung an die oft sauerstoffarme und kohlenstoffdioxidreiche Umgebung im Untergrund das Hämoglobin des Blutes modifiziert.[24][25] Jedes Individuum beansprucht ein eigenes Gangsystem. Dadurch leben die Tiere einzelgängerisch und territorial. Einen Winterschlaf halten die Neuweltmaulwürfe nicht. Soweit bekannt, sind sie sowohl tag- als auch nachtaktiv. Der hauptsächliche Aufenthalt im Untergrund wird durch gelegentliche Besuche an der Erdoberfläche unterbrochen.[2]
Die Nahrung der Neuweltmaulwürfe besteht aus Regenwürmern sowie einer Vielzahl an Insekten und deren Larven. Die Zusammensetzung der Nahrung variiert regional. Einige Arten nehmen zusätzlich unterirdische Pflanzenteile zu sich. Die Tiere sind durch eine hohe Stoffwechselrate gekennzeichnet, sie verzehren täglich oft eine Menge entsprechend oder über ihrem eigenen Körpergewicht. Die Fortpflanzungszeit der meisten Arten findet im ersten Jahresquartal statt. In der Regel kommt nur einmal im Jahr Nachwuchs zur Welt. Die Tragzeit beträgt vermutlich vier bis sechs Wochen. Der Wurf des Weibchens umfasst zwei bis fünf Jungtiere. Der Nachwuchs ist nackt und blind. Die Individualentwicklung verläuft recht schnell. Die Saugphase währt nur rund einen Monat, danach verlassen die Jungen das mütterliche Nest. Eigener Nachwuchs wird dann in der Regel im darauffolgenden Jahr gezeugt. Die Lebenserwartung der Tiere kann schätzungsweise bis zu sechs Jahre betragen. Abweichend von den Eigentlichen Maulwürfen ist bei den Neuweltmaulwürfen kein Hermaphroditismus belegt.[26][2]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[27]
|
Die Neuweltmaulwürfe sind eine Tribus aus der Familie der Maulwürfe (Talpidae) und der Ordnung der Insektenfresser (Eulipotyphla). Sie schließen vergleichbar den Eigentlichen Maulwürfen (Talpinia) grabende Vertreter der Familie ein, die sich aber – abweichend von ihrem deutschen Trivialnamen – sowohl auf die Neue als auch auf die Alte Welt verteilen. Die Linien der Neuweltmaulwürfe und der Eigentlichen Maulwürfe sind nicht unmittelbar miteinander verwandt, sondern entwickelten ihre Grabeigenschaften unabhängig voneinander. Andere Vertreter der Familie leben dagegen nur teilweise unterirdisch, bewegen sich oberirdisch fort oder sind an eine semi-aquatische Lebensweise angepasst.[27] In ihren äußeren Merkmalen zeigen die Neuweltmaulwürfe die generellen Kennzeichen der grabenden Maulwürfe. Sie werden somit durch einen kurzen und kräftigen Ober- beziehungsweise Unterarm gekennzeichnet. Die großen und schaufelartigen Hände sind dauerhaft nach außen gedreht. Der Schwanz ist bei den Neuweltmaulwürfen merklich länger als bei den Eigentlichen Maulwürfen. Ein weiteres Kennzeichen bilden die deutlich reduzierten Augen. Die Maulwürfe entstanden molekulargenetischen Studien zufolge bereits im Mittleren Eozän vor rund 47 Millionen Jahren. Die ursprünglichste Gruppe stellen die Spitzmausmaulwürfe (Uropsilinae) dar, sie spalteten sich zuerst von der gemeinsamen Vorfahrenlinie ab. Aus systematischer Sicht stehen sie als eigenständige Unterfamilie den Altweltmaulwürfen (Talpinae) gegenüber und bilden so deren Schwestergruppe. Die Altweltmaulwürfe vereinen alle übrigen Maulwürfe, einschließlich der Neuweltmaulwürfe. Innerhalb dieser trennten sich die Neuweltmaulwürfe im Oberen Eozän vor rund 39 bis 35 Millionen Jahren von den anderen Triben ab. Eine weitere Diversifizierung der Neuweltmaulwürfe setzte im Unteren Miozän ein. Einzelnen genetischen Untersuchungen nach nehmen die Neuweltmaulwürfe eine basale Position innerhalb der Altweltmaulwürfe ein.[28][27][3]
Innere Systematik der Neuweltmaulwürfe nach Chen et al. 2021[3]
|
Die Neuweltmaulwürfe gliedern sich heute in insgesamt fünf Gattungen:[2][3][29]
- Tribus: Neuweltmaulwürfe (Scalopini Gill, 1875)
- Alpiscaptulus Jiang & Chen, 2021; Ostasien (südwestliches China), 1 Art
- Haarschwanzmaulwurf (Parascalops Bachman, 1841); Nordamerika (südliches Kanada sowie nordöstliche und östliche USA), 1 Art
- Ostamerikanischer Maulwurf (Scalopus É. Geoffroy Saint-Hilaire, 1803); östliches Nordamerika (südliches Kanada bis nördliches Mexiko), 1 Art
- Gansu-Maulwurf (Scapanulus Thomas, 1912); Ostasien (zentrales China), 1 Art
- Westamerikanische Maulwürfe (Scapanus Pomel, 1848); westliches Nordamerika (südliches Kanada bis nördliches Mexiko), 5 Arten
Diese fünf Gattungen lassen sich zwei unterschiedlichen Entwicklungslinien innerhalb der Neuweltmaulwürfe zuordnen. Die eine umfasst die Parascalopina mit dem Haarschwanzmaulwurf und dem Gansu-Maulwurf sowie dessen nächster Verwandter Alpiscaptulus. Die andere wird durch die Scalopina repräsentiert und fasst den Ostamerikanischen und die Westamerikanischen Maulwürfe zusammen. Die beiden Linien unterscheiden sich anhand der Ausprägung des Metastylids am unteren zweiten Molar: bei den Scalopina fehlt dieser, bei den Parascalopina kommt er hingegen vor.[30][23] Molekulargenetisch sind beide Entwicklungslinien wenigstens bis in das Untere Miozän vor 21,4 Millionen Jahren zurückverfolgbar.[3]
Innere Systematik der Neuweltmaulwürfe nach Schwermann et al. 2019[23]
|
Neben den heutigen Vertretern sind noch weitere ausgestorbene beschrieben worden:
- Domninoides Green, 1956
- Hugueneya van den Hoek Ostende, 1989
- Leptoscaptor Ziegler, 2003
- Proscapanus Gaillard, 1899
- Mioscalops Ostrander, Mebrate & Wilson, 1986
- Scapanoscapter Hutchinson, 1968
- Yanshuella Storch & Qiu, 1983
- Yunoscaptor Storch & Qiu, 1990
Einige weitere Formen wurden ursprünglich ebenfalls zu den Neuweltmaulwürfen gestellt. Hierzu gehört Proscalops, das im Jahr 1901 von William Diller Matthew benannt worden war.[31] Rund 60 Jahre später lagerte Katherine M. Reed die Gattung zunächst in eine eigenständige Unterfamilie aus,[32] wiederum 20 Jahre darauf hob Anthony D. Barnosky diese mit den Proscalopidae in einen unabhängigen Familienstatus.[33] Unklar ist die Stellung von Mydecodon, das wiederum auf Robert Warren Wilson aus dem Jahr 1960 zurückgeht.[34] Nur wenige Jahre später hatte J. Howard Hutchinson die Form aber mit Mystipterus aus der Gruppe der Spitzmausmaulwürfe gleichgesetzt.[30] Andere Autoren geben die Gattung Mydecodon aber noch als gültigen Vertreter der Neuweltmaulwürfe an.[35] Von Wilson aus dem gleichen Jahr stammt zusätzlich die Gattung Scalopoides. Die Bezeichnung ist aber durch eine fossile Form der Käfer präokkupiert. Daher wurde im Jahr 1986 von Gregg E. Ostrander und Kollegen der Alternativname Mioscalops vorgeschlagen, der jedoch bisher nur selten Gebrauch fand.[36] Von Miklós Kretzoi wiederum stammt die Synonymbezeichnung Wilsonius aus dem Jahr 2000.[23]
Forschungsgeschichte

Der Ostamerikanische Maulwurf gehörte zu den ersten Maulwürfen, die aus Nordamerika bekannt wurden. Bereits Carl von Linné verlieh ihm im Jahr 1758 in seinem Systema naturae den wissenschaftlichen Artnamen Sorex aquaticus und ordnete ihn damit den Spitzmäusen bei. Gleichzeitig führte er den Sternmull ein, den er ebenfalls den Spitzmäusen zuwies.[37] Der heute gültige Gattungsname Scalopus wurde von Étienne Geoffroy Saint-Hilaire im Jahr 1803 etabliert.[38] Dabei verwendete er eine Bezeichnung von Georges Cuvier (Scalope) aus dem Jahr 1800 wieder, die aufgrund der fehlenden Latinisierung als nicht gültig angesehen wird. Jedoch hatte Geoffroy Saint-Hillaire damit erstmals einen nordamerikanischen Maulwurf von den bekannten eurasischen Vertretern, die zu jener Zeit meist unter Talpa geführt wurden, abgesetzt. Auf Scalopus geht auch die wissenschaftliche Bezeichnung der Tribus zurück. Diese stammt von Theodore Gill aus dem Jahr 1875. Gill erarbeitete in einem umfassenden Aufsatz eine systematische Gliederung der Insektenfresser und teilte innerhalb dieser die Maulwürfe in zwei Unterfamilien: die Talpinae (die eigentlichen grabenden Maulwürfe) und die Myogalinae (die heutigen Desmane). Der Unterfamilie der Talpinae wies er drei größere Gruppen zu, die er mit „Talpae“ (die Eigentlichen Maulwürfe), „Condylurae“ (der Sternmull) und „Scalopes“ (die Neuweltmaulwürfe) benannte.[39] Seine höhere systematische Gliederung entlehnte Gill von St. George Jackson Mivart aus dem Jahr 1868.[40] Gelegentlich wird aber auch Édouard Louis Trouessart als Autor der Tribus angegeben, der 1879 ein zu Gill vergleichbares System vorstellte. Hierbei führte er die drei Gruppen der Talpinae mit „Talpeae“, „Condylureae“ und „Scalopeae“.[41] Vergleichbar zu Gill und Trouessart verfuhr auch George Edward Dobson im Jahr 1883.[4]
Die Rangstufung der Neuweltmaulwürfe wurde in der forschungsgeschichtlichen Vergangenheit unterschiedlich bewertet. Während Gill, Trouessart und Dobson diese unterhalb der Unterfamilie einordneten, was in etwa dem Rang einer Tribus entspricht, sah Edward Drinker Cope sie 1889 auf Ebene einer eigenständigen Familie.[42] Von Oldfield Thomas stammt eine Unterteilung der Maulwürfe in fünf Unterfamilien, von denen er eine mit den Scalopinae, also den Neuweltmaulwürfen, angab. Ihnen stellte er die Desmaninae (Desmane), die Talpinae (Eigentliche Maulwürfe), die Uropsilinae (Spitzmausmaulwürfe) und Condylurinae (Sternmull) zur Seite.[43] Dies fand dann Einzug in die generelle Klassifikation der Säugetiere durch George Gaylord Simpson im Jahr 1945.[44] Leigh Van Valen sah dies rund 22 Jahre später differenzierter. Er erkannte nur drei rezente Unterfamilien innerhalb der Maulwürfe an und ordnete die Neuweltmaulwürfe und den Sternmull neben einzelnen weiteren Gruppen den Talpinae als jeweils eigenständige Triben unter.[10] Einige nachfolgende Systematiken wie jene von Malcolm C. McKenna und Susan K. Bell aus dem Jahr 1997 übernahmen dieses Schema weitgehend.[45] Andere wiederum hoben die Neuweltmaulwürfe erneut auf die Ebene der Unterfamilie und vereinten innerhalb dieser die Triben der Scalopini und der Condylurini, womit der Sternmull ein Bestandteil der Neuweltmaulwürfe wurde. Dies findet sich unter anderem im Werk Mammal Species of the World von Don E. Wilson und DeeAnn M. Reeder aus dem Jahr 2005.[46]
Diesen verschiedenen klassischen und zumeist auf anatomischen Merkmalen beruhenden Klassifizierungsschemata stehen molekulargenetische Untersuchungen gegenüber. Dadurch wurde im Jahr 2016 erkannt, dass mit Ausnahme der Spitzmausmaulwürfe alle übrigen Maulwürfe, also die Eigentlichen Maulwürfe, die Neuweltmaulwürfe, der Sternmull, die Desmane, der Langschwanzmaulwurf und die verschiedenen Spitzmulle, eine monophyletische Einheit bilden. Die Neuweltmaulwürfe stehen hierbei relativ basal in der Verwandtschaftsfolge. Der Sternmull erwies sich aber als mit den Desmanen näher verwandt, die sich beide wiederum in die Nähe der Eigentlichen Maulwürfe positionieren. Daher werden in einer modernen Einteilung der Maulwürfe alle diese Formen als eigenständige Triben innerhalb der Unterfamilie der Talpinae geführt. Dies gilt auch für die Neuweltmaulwürfe als Tribus der Scalopini,[27] die damit aus Sicht der deutschen Trivialnamensgebung einen Bestandteil der Altweltmaulwürfe formen. Die Ergebnisse der Molekulargenetik fanden auch im achten Band des Standardwerkes Handbook of the Mammals of the World aus dem Jahr 2018 Berücksichtigung.[2] Die genetischen Daten geben weiter darüber Auskunft, dass die grabenden Maulwürfe in keiner unmittelbar direkten Verwandtschaftsgemeinschaft eingebunden sind. Dadurch kam es offensichtlich in der Evolution der Maulwürfe mehrmals zur Entwicklung von Formen, die auf eine unterirdische Lebensweise spezialisiert sind.[27]
Stammesgeschichte
Die Neuweltmaulwürfe verfügen über einen recht umfangreichen Fossilbericht. Ähnlich den rezenten Vertretern verteilt sich dieser auf die Neue und die Alte Welt. Der Ursprung der Neuweltmaulwürfe wird in Eurasien vermutet. Die dort verbreiteten heutigen Vertreter stellen dadurch eher Reliktformen dar als dass sie die Funktion von Pionierarten einnehmen. Im Laufe der Stammesgeschichte kam es zu einer mehrfachen Migration zwischen den Kontinenten.[22][23][3]
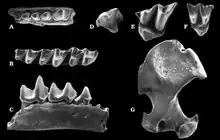
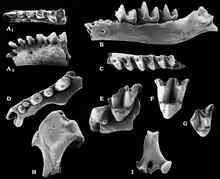
Die ältesten Funde der Neuweltmaulwürfe sind aus Eurasien nachweisbar. Sie datieren in den Übergang vom Oberen Oligozän zum Unteren Miozän und liegen vom westlichen Bereich des Großkontinentes vor. Hier ist die Gattung Hugueneya zu nennen, deren Typusfundstelle Eggingen bei Ulm in Baden-Württemberg bildet. Von dort ist sie weitgehend über Zähne bekannt.[47][48] Diese recht große Maulwurfsfform trat möglicherweise im gleichen Zeitraum sehr weit östlich im Gebiet nördlich des heutigen Aralsees in Kasachstan auf, wo sie mit einzelnen Unterkiefer- und Langknochenresten aus der Aral-Formation vorgestellt wurde.[49][50] Im Unteren Miozän erschien die Gattung Proscapanus, die eine recht weite Verbreitung erreichte. Sie ist unter anderem von Sansan in der südfranzösischen Region Okzitanien beschrieben worden, von wo auch das Typusmaterial der Gattung in Form einzelner Gebissfragmente stammt, das Claude Gaillard im Jahr 1899 zu ihrer Aufstellung nutzte.[51] Als ein weiterer bedeutender Fundpunkt können verschiedenste Spaltenfüllungen von Petersbuch in Bayern angegeben werden, deren zeitliche Reichweite das Untere und Mittlere Miozän vor 18 bis 11 Millionen Jahren umfasst.[52][53][54][55][56] Relativ späte Funde sind mit einem Alter von rund 9,6 Millionen Jahren aus einzelnen Lokalitäten in Österreich wie Schernham berichtet worden. Fast alle mitteleuropäischen Fundbereiche liegen im heutigen Molassegebiet. Das Fossilmaterial von Proscapanus besteht weitgehend aus Einzelfunden, weist aber zahlreiche Variationen auf und wird so mehreren Arten zugewiesen. Aus Viehhausen bei Regensburg wurde darüber hinaus ein Teilskelett geborgen, bei dem der Schädel, die Wirbelsäule und die Vordergliedmaßen einschließlich verschiedener Partien des Handskeletts überliefert sind. Es zeigt den typisch vergrößerten vorderen Schneidezahn der Neuweltmaulwürfe an und lässt einen stark spezialisierten Gräber erkennen. An der Hand setzte seitlich das Sesambein an, das bei heutigen Neuweltmaulwürfen den „Vordaumen“ bildet.[18] Die Spaltenfüllungen von Petersbuch sind mit einer weiteren Form der Neuweltmaulwürfe assoziiert, die im Jahr 2003 als Leptoscaptor wissenschaftlich eingeführt worden war. Es handelt sich um eine mittelgroße Form, die lokal recht große Häufigkeit erreichen kann. Eine Besonderheit an dem umfangreichen Gebissmaterial ist die gegenüber Proscapanus beginnende Reduktion der Prämolaren.[54]
Aus dem Übergang vom Oberen Miozän zum Pliozän ist vor allem in Ostasien die Gattung Yanshuella belegt, ein kleiner Maulwurf mit grazilen Gliedmaßen, eng stehenden vorderen Zähnen, einer reduzierten Anzahl an Prämolaren und hohen Höckerchen auf den Mahlzahnkronen. Besonders umfangreiches Material stammt von den Fundstellen Ertemte und Harr Obo in der Inneren Mongolei. Allein erstere erbrachte mehrere hundert isolierte Zähne, fast 70 Ober- und Unterkieferfragmente sowie ähnlich viele Langknochen, von denen der weitaus größte Teil auf den Oberarmknochen entfällt. Der letztere Fundpunkt war nicht ganz so fundreich, zeigt aber ein ähnliches Spektrum. Beide Lokalitäten sind schon seit dem Anfang des 20. Jahrhunderts bekannt. Einen ersten Überblick über die Maulwürfe dort verschaffte im Jahr 1924 Max Schlosser, der das Material aber einer frühen Form des Kurzgesichtmaulwurfs (Scaptochirus) zuwies. Dies wurde erst rund 60 Jahre später durch Gerhard Storch und Qiu Shuding korrigiert, als sie die neue Gattung Yanshuella einführten.[57] Recht zahlreiche Gebissstücke und einzelne Gleidmaßenelemente sind auch aus der etwa zeitgleichen Mahui-Formation des Yushe-Beckens in der nordchinesischen Provinz Shanxi aufgedeckt worden.[58] Die Form kam möglicherweise auch in Nordamerika vor, wie einzelne Unterkieferfragmente vom Columbia River im US-Bundesstaat Oregon zeigen.[30][57] Die Mahui-Formation und zusätzlich die zeitlich etwas jüngere Gaozhuang-Formation im Yushe-Becken förderten auch Reste von Yunoscaptor zu Tage. Diese bestehen aus einzelnen Unterkieferfragmenten, isolierten Zähnen sowie Oberarmknochen. Das Fossilmaterial verweist auf einen gegenüber Yanshuella noch kleineren Maulwurf mit sehr niederkronigen Mahlzähnen und einem fehlenden Vormahlzahn in der unteren Zahnreihe.[58] Das Belegmaterial der Gattung wurde wiederum im Jahr 1990 aus der obermiozänen Shihuiba-Formation in Lufeng in der chinesischen Provinz Yunnan beschrieben. Es setzt sich aus einem Unterkiefer, Einzelzähnen und Langknochen zusammen.[59]
In Nordamerika, dem heutigen Hauptverbreitungsgebiet der Neuweltmaulwürfe, reichen Fossilfunde bis in das Miozän zurück. Bedeutend in diesem Zusammenhang ist die Observation Quarry im Dawes County im US-Bundesstaat Nebraska, von wo Unterkieferfragmente und ein Oberarmknochen von Mioscalops berichtet wurden. Aus biostratigraphischer Sicht lassen sich die Reste dem Übergang vom Unteren zum Mittleren Miozän zuweisen.[60] Das Typusmaterial der Gattung wurde aus der Pawnee-Creek-Formation im Logan County in Colorado geborgen. In seiner Erstbeschreibung aus dem Jahr 1960 bezeichnete Robert W. Wilson diese noch mit Scalopoides und gründete sie auf einige Unterkieferteile. Erst mehr als 20 Jahre später wurde sie in Mioscalops umbenannt. Eine umfangreiche Kollektion bestehend aus Gebissresten und Teilen des Bewegungsapparates ist aus dem Quartz Basin im Osten des US-Bundesstaates Oregon überliefert. Dieses Material datiert in das Mittlere Miozän. Mioscalops wies bereits eine leicht reduzierte Zahnformel auf, da zumindest im Unterkiefer der vordere Prämolar fehlte. Die Backenzähne waren außerdem nur mäßig hochkronig. Die Form wich im Körperbau etwas von den heutigen Neuweltmaulwürfen ab und zeigte einzelne Übereinstimmungen mit dem Sternmull (Condylura´) und dem Amerikanischen Spitzmull (Neurotrichus). Dies betrifft unter anderem die schwach entwickelten Vorderbeine oder das langgestreckte Schlüsselbein, beides sind möglicherweise Ausdruck weniger gut ausgeprägter Grabeigenschaften. Der Oberarmknochen wies aber eine markante scalopine ridge als Charakteristikum der Neuweltmaulwürfe auf.[34][30][23] Teilweise wurde Mioscalops auch aus Europa berichtet, doch ist die Zuweisung in keinem Fall gesichert.[47][61][23] Im Mittleren und Oberen Miozän ist in Nordamerika zusätzlich noch Domninoides belegt. Einige wenige Kieferbruchstücke und Zähne mit mittelmiozäner Zeitstellung brachten verschiedene Fundstellen in Oregon zu Tage, so unter anderem das Quartz Basin.[30] Weiteres Fundmaterial lieferte die Valentine-Formation mit einzelnen Lokalitäten im Cherry County und im Keya Paha County von Nebraska. Die recht reiche Kollektion aus dem Keya Paha County beinhaltet auch einen Teilschädel, während jene aus dem Cherry County aus einem Kieferbruchstück, Einzelzähnen und Skelettelementen besteht.[62][63] Bereits in das Untere Pliozän datiert die Ogallala-Formation im Trego County von Kansas, aus der ebenfalls isolierte Zähne und Gliedmaßenknochen geborgen wurden.[64] Die Typusfundstelle der Gattung mit einem nahezu vollständigen linken Unterkieferast befindet sich hingegen mit dem Wolf Creek im Shannon County von South Dakota. Sie ist Teil der Ogallala-Fauna mit einer vergleichbaren Datierung zu den Funden aus dem Trego County. Morton Green, der Erstbeschreiber von Domninoides führte die Form im Jahr 1956 zuerst als Vertreter der Spitzmäuse ein,[65] der maulwurfartige Charakter der Funde wurde fünf Jahre später erkannt. Die Tiere von den Ausmaßen des Ostamerikanischen Maulwurfs '(Scalopus) waren stark an die grabende Lebensweise angepasst und besaßen ein reduziertes Gebiss.[30][63] Wiederum schon im Mittleren Miozän trat Scapanoscapter auf, dessen Typusmaterial dem Red Basin im zentralen Osten von Oregon entstammt. Im Jahr 1968 von J. Howard Hutchinson basierend auf Kiefer- und Gliedmaßenfragmenten definiert, besteht wahrscheinlich eine engere Bindung an die rezente Gattung Scapanus mit den Westamerikanischen Maulwürfen, mit denen die Tiere eine vollständige Bezahnung teilten. Abweichend von den heutigen Arten besaßen bei Scapanoscapter alle Prämolaren noch zwei Wurzeln. Außerdem waren die Mahlzähne eher niederkronig und der zweite untere Schneidezahn noch nicht vergrößert.[30]
Die Vorformen der heute noch bestehenden Arten traten schon recht früh in Erscheinung. So ist die Gattung Scapanulus erstmals im Unteren Miozän in Südostasien belegbar.[66] Die amerikanischen Neuweltmaulwürfe haben ihre frühesten Nachweise mit Scapanus im Mittleren Miozän und mit Scalopus im Oberen Miozän. Kurz darauf ist auch Parascalops fassbar, wobei frühe Funde sowohl aus Nordamerika als auch aus Europa überliefert sind.[30][67][35][68]
Literatur
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. The Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4
Einzelnachweise
- Robert S. Hoffmann und Darrin Lunde: Order Soricomorpha – Shrews and Moles. In: Andrew T. Smith und Yan Xie (Hrsg.): A Guide to the Mammals of China. Princeton University Press, 2008, ISBN 978-0-691-09984-2, S. 321
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- Zhong-Zheng Chen, Shui-Wang He, Wen-Hao Hu, Wen-Yu Song, Kenneth O Onditi, Xue-You Li und Xue-Long Jiang: Morphology and phylogeny of scalopine moles (Eulipotyphla: Talpidae: Scalopini) from the eastern Himalayas, with descriptions of a new genus and species. Zoological Journal of the Linnean Society, 2021, S. zlaa172, doi:10.1093/zoolinnean/zlaa172
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Frederick W. True: A revision of the American moles. Proceedings of the United States National Museum 19 (1101), 1896, S. 1–112
- Hartley H. T. Jackson: A review of the American moles. North American Fauna 38, 1915, S. 1–100
- S. U. Stroganov: Morphological characters of the auditory ossicles of recent Talpidae. Journal of Mammalogy 26 (4), 1945, S. 12–42
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Daisuke Koyabu, Misato Hosojima und Hideki Endo: Into the dark: patterns of middle ear adaptations in subterranean eulipotyphlan mammals. Royal Society Open Science 4, 2017, S. 170608, doi:10.1098/rsos.170608
- Leigh Van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- B. Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. American Journal of Anatomy 64, 1939, S. 1–39
- Charles A. Reed: Locomotion and Appendicular Anatomy in Three Soricoid Insectivores. The American Midland Naturalist 45, 1951, S. 513–671 ()
- Marcelo R Sánchez-Villagra, Peter R. Menke und Jonathan H. Geisler: Patterns of evolutionary transformation in the humerus of moles (Talpidae, Mammalia): A character analysis. Mammal Study 29 (2), 2004, S. 163–170, doi:10.3106/mammalstudy.29.163
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumenting the polydactyly ‚constraint‘: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- Constanze Bickelmann, Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Transcriptional heterochrony in talpid mole autopods. EvoDevo 3, 2012, S. 16, doi:10.1186/2041-9139-3-16
- Achim H. Schwermann und Richard S. Thompson: Extraordinarily preserved talpids (Mammalia, Lipotyphla) and the evolution of fossoriality. Journal of Vertebrate Paleontology 35 (4), 2015, S. e934828, doi:10.1080/02724634.2014.934828
- Shin-ichiro Kawada, Masashi Harada, Kazuhiru Koyasu und Sen-ichi Oda: Karyological note on the short-faced mole, Scaptochirus moschatus (Insectivore, Talpidae). Mammal Study 27, 2002, S. 91–94, ()
- Shin.ichiro Kawada, Song Li, Ying-Xiang Wang, Orin B. Mock, Sen-ichi Oda und Kevin L. Campbell: Karyotype evolution of shrew moles (Soricomorpha: Talpidae). Journal of Mammalogy 89 (6), 2008, S. 1428–1434
- Kai He, Jin-Huan Wang, Wei-Ting Su, Quan Li, Wen-Hui Nie und Xue-Long Jiang: Karyotype of the Gansu mole (Scapanulus oweni): further evidence for karyotypic stability in talpid. Mammal Study 37, 2012, S. 341–348
- Marcelo R. Sánchez-Villagra, Inés Horovitz und Masaharu Motokawa: A comprehensive morphological analysis of talpid moles (Mammalia) phylogenetic relationships. Cladistics 22, 2006, S. 59–88
- Achim Schwermann, Kai He, Benjamin J. Peters, Thorsten Plogschties und Gabrielle Sansalone: Systematics and macroevolution of extant and fossil scalopine moles (Mammalia, Talpidae). Palaeontology 62 (4), 2019, S. 661–676, doi:10.1111/pala.12422
- Kevin L. Campbell, Jay F. Storz, Anthony V. Signore, Hideaki Moriyama, Kenneth C. Catania, Alexander P. Payson, Joseph Bonaventura, Jörg Stetefeld und Roy E. Weber: Molecular basis of a novel adaptation to hypoxic-hypercapnia in a strictly fossorial mole. BMC Evolutionary Biology 10, 2010, S. 214, doi:10.1186/1471-2148-10-214
- Anthony V. Signore, Jörg Stetefeld, Roy E. Weber und Kevin L. Campbell: Origin and mechanism of thermal insensitivity in mole hemoglobins: a test of the ‘additional’ chloride binding site hypothesis. Journal of Experimental Biology 215 (3), 2012, S. 518–525, doi:10.1242/jeb.063669
- N. M. Rubenstein, G. R. Cunha, Y. Z. Wang, K. L. Campbell, A. J. Conley, K. C. Catania, S. E. Glickman und N. J. Place: Variation in ovarian morphology in four species of New World moles with a peniform clitoris. Reproduction 126, 2003, S. 713–719
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. In: Molecular Biology and Evolution. 34 (1), 2016, S. 78–87
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Sergio Ticul Álvarez-Castañeda und Patricia Cortes-Calva: Revision of moles in the genus Scapanus. Therya 12 (2), 2021, S. 275–281, doi:10.12933/therya-21-1174
- J. Howard Hutchinson: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117 (S. 58–96)
- William Diller Matthew: Fossil mammals of the Tertiary of northeastern Colorado. Memoirs of the American Museum of Natural History 1 (7), 1901, S. 353–447 (S. 376–376)
- Katherine M. Reed: The Proscalopinae, a new subfamily of talpid insectivores. Bulletin of the Museum of Comparative Zoology at Harvard College 125, 1961, S. 473–494 ()
- Anthony D. Barnosky: A Skeleton of Mesoscalops (Mammalia, Insectivora) from the Miocene Deep River Formation, Montana, and a Review of the Proscalopid Moles: Evolutionary, Functional, and Stratigraphic Relationships. Journal of Vertebrate Paleontology 1 (3/4), 1981, S. 285–339
- Robert W. Wilson: Early Miocene rodents and insectivores from northeastern Colorado. University of Kansas Paleontological Contributions, Vertebrata, Article 7, 1960, S. 1–92 (S. 38–49)
- Barbara Rzebik-Kowalska: Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland. Palaeontologia Electronica 17 (2), 2014, S. 17.2.26A, doi:10.26879/457
- Gregg E. Ostrander, Assefa Mebrate und Robert W. Wilson: Type and figured specimens of fossil vertebrates in the collection of the University of Kansas Museum of Natural History. Part IV. Fossil mammals. University of Kansas Museum of Natural History Miscellaneous Publication 79, 1986, S. 1–83 (S. 9) ()
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 53 ()
- Étienne Geoffroy Saint-Hilaire: Catalogue des Mammifères du Museum National d'histoire naturelle. Paris, 1803, S. 1–272 (S. 77) ()
- Theodore Gill: Synopsis of the insectivorous mammals. Bulletin of the United States Geological and Geographical Survey of the Territories 1, 1874–1875, S. 91–140 (S. 106) ()
- St. George Jackson Mivart: Notes on the Osteology of the Insectivora. Journal of Anatomy and Physiology 2 (1), 1868, S. 117–154 (S. 150) ()
- Édouard Louis Trouessart: Catalogue des mammifères vivants et fossiles. Insectivores. Revue et Magasin de Zoologie 3 (7), 1879, S. 219–285 (S. 267–272) ()
- Edward Drinker Cope: Synopsis of the Families of Vertebrata. The American Naturalist 23 (274), 1889, S. 849–877
- Oldfield Thomas: On the collection of mammals from the Tsin-ling mountains, Central China, presented by Mr. G. Fenwick Owen to the National Museum. Annals and Magazine of Natural History 8 (10), 1912, S. 395–403 ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 52–53)
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 279–283)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- J. Howard Hutchinson: Notes on type specimens of European Miocene Talpidae and a tentative Classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios 7 (3), 1974, S. 211–256
- Lars W. van den Hoek Ostende: The Talpidae (Insectivora, Mammalia) of EggingenMittelhart (Baden-Württemberg, F.R.G.) with special reference to the Paratalpa-Desmanodon lineage. Stuttgarter Beiträge zur Naturkunde Serie B 152, 1989, S. 1–29
- Spencer George Lucas, Elena G. Kordikova und Robert J. Emry: Oligocene stratigraphy, sequence stratigraphy, and mammalian biochronology north of the Aral Sea, Western Kazakstan. Bulletin of the Carnegie Museum of Natural History 34, 1998, S. 313–348 ()
- Alexey V. Lopatin: Early Miocene small mammals from the North Aral region (Kazakhstan) with special reference to their biostratigraphic significance. Paleontological Journal 38 (Suppl. 3), 2004, S S217–S323
- Claude Gaillard: Mammifères Miocènes nouveaux ou peu connus de La Grive-Saint-Alban (Isère). Archives du Muséum d'histoire naturelle de Lyon 7, 1899, S. 1–68 ()
- Reinhard Ziegler: Talpiden (Mammalia, Insectivora) aus dem Orleanium und Astaracium Bayerns. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 25, 1985, S. 131–175
- Reinhard Ziegler: Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Paläontologie Serie B 167, 1990, S. 1–80
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- Reinhard Ziegler: Miocene Insectivores from Austria and Germany - An Overview. Beiträge zur Paläontologie 30, 2006, S. 481–494
- Johannes Klietmann, Doris Nagel, Michael Rummel und Lars W. van den Hoek Ostende: A gap in digging: the Talpidae of Petersbuch 28 (Germany, Early Miocene). Paläontologische Zeitschrift 89, 2015, S. 563–592, doi:10.1007/s12542-014-0228-2
- Gerhard Storch und Shuding Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Lawrence J. Flynn und Wen-Yu Wu: The Lipotyphla of Yushe Basin. In: Lawrence J. Flynn und Wen-Yu Wu (Hrsg.): Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals, Volume II: Small Mammal Fossils of Yushe Basin, Vertebrate Paleobiology and Paleoanthropology. Springer Science+Business Media B.V., 2017, S. 11–26
- Gerhard Storch und Shuding Qiu: Insectivores (Mammalia: Erinaceidae, Soricidae, Talpidae) from the Lufeng hominoid locality, late Miocene of China. Geobios 24 (5), 1991, S. 601–621
- William W. Korth und Robert L. Evander: Lioptyphla, chiroptera, lagomorpha, and rodentia (Mammalia) from Observation Quarry, earliest Barstovian (Miocene), Dawes County, Nebraska. Annals of Carnegie Museum 83 (3), 2016, S. 219–254
- Vasil V. Popov: Late Pliocene Erinaceidae and Talpidae (Mammalia: Insectivora) from Varshets (North Bulgaria). Acta zoologica cracoviensia 47 (1–2), 2004, S. 61–80
- Katherine M. Reed: Two new species of the fossil talpid insectivores. Breviora 168, 1962, S. 1–7 ()
- Patricia Waring Freeman: Redescription and Comparison of a Highly Fossorial Mole, Domninoides mimicus (Insectivora, Talpidae), from the Clarendonian. American Museum Novitates 2667, 1979, S. 1–16
- Richard Leland Wilson: Systematics and faunal analysis of a Lower Pliocene vertebrate assemblage from Trego County, Kansas. Contributions from the Museum of Paleontology, The University of Michigan 22 (7), 1968, S. 75–126
- Morton Green: The Lower Pliocene Ogallala-Wolf Creek Vertebrate Fauna, South Dakota. Journal of Paleontology 30 (1), 1956, S. 146–169
- Pierre Mein und Léonard Ginsburg: Les mammifères du gisement miocène inférieur de Li Mae Long, Thaïlande: systématique, biostratigraphie et paléoenvironnement. Geodiversitas 19 (4), 1997, S. 783–844
- Walter W. Dalquest: Mammals of the Coffee Ranch local fauna, Hemphilian of Texas. The Pearce-Sellards Series 38, 1983, S. 1–41
- Danielle Oberg: Fossil Moles from the Gray F ossil Site, TN: Implications for Diversification and Evolution of North American Talpidae. East Tennessee State University, 2018, S. 1–95