Nemegt-Formation
Die Nemegt-Formation ist eine lithostratigraphische Einheit aus der Oberkreide des Nemegt-Beckens und angrenzender Gebiete in den Provinzen Ömnö-Gobi-Aimag und Bajanchongor-Aimag im Süden der heutigen Mongolei. Die Formation wurde weltweit bekannt durch ihren Reichtum an Wirbeltierfossilien aus dem obersten Campanium bis unteren/mittleren Maastrichtium, der obersten Stufe der Oberkreide.
Definition und Typlokalität
Für die Transliteration der mongolischen Namen für Lokationen und stratigraphische Einheiten existiert keine einheitliche Regelung. In der Fachliteratur finden sich daher oft sehr unterschiedliche Schreibweisen für dieselben Begriffe. In diesem Artikel hier wird eine Schreibweise in Anlehnung an die Vorschläge von Michael J. Benton et al. verwendet.[1] Alternative Transliterationen werden einmalig in Klammer gleichgesetzt, falls dies für das Verständnis der zitierten Fachliteratur angebracht scheint.

Die Nemegt-Formation ist eine mehr als 235 m mächtige Abfolge[2] von meist nur schwach verfestigten, hellen, meist gelblichen bis graubraunen, seltener roten oder orangen Sanden. Geringmächtige Lagen von sandigen Ton-/Schluffsteinen, Sandsteinen oder Konglomeraten/Kiesen treten zwar häufig auf, nehmen jedoch nur einen geringen Anteil des Gesamtvolumens ein. Die Formation wird im Wesentlichen von jeweils etwa 2–9 m mächtigen fining-upward Zyklen aufgebaut; das bedeutet, dass innerhalb der Formation immer wieder ähnliche Sedimentabfolgen auftreten, wobei in jeder Abfolge die durchschnittliche Korngröße vom Liegenden ins Hangende abnimmt. Die Basis jeder dieser Zyklen bildet in der Regel eine Erosionsfläche. Schrägschichtung, Kreuzschichtungskörper innerhalb der einzelnen Schichten und Erosionsrinnen sind typische Sedimentstrukturen der Formation.[3]
Die Sedimente der Nemegt-Formation liegen konkordant über den Sandsteinen der Baruungoyot-Formation (= Barun-Goyot-Formation), wobei im Liegendsten, mindestens 23 m mächtigen, Abschnitt der Nemegt-Formation Sandsteine in der für die Baruungoyot-Formation typischen Fazies mit den charakteristischen Sedimenten der Nemegt-Formation verzahnt sind.[4] Die Formation endet im Hangenden mit einer Erosionsfläche auf der Sedimente des Paläozän abgelagert wurden.[3]
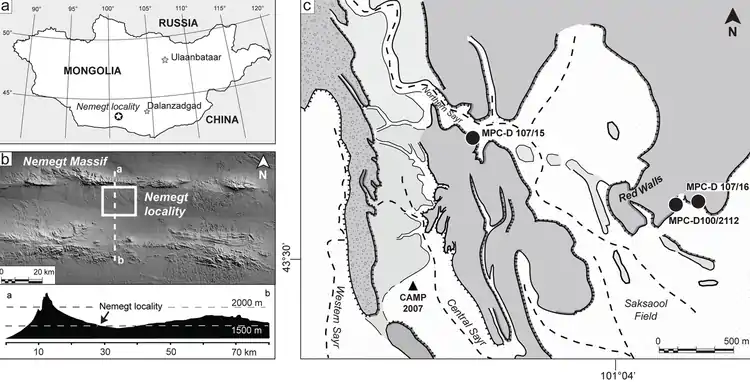
Aufschlüsse der Nemegt-Formation finden sich vor allem in den Badlands des Nemegt-Beckens. An der Typlokalität Nemegt ist die Basis der Formation mit dem Übergang zur unterlagernden Baruungoyot-Formation aufgeschlossen. Referenzprofile zur Definition der Formation wurden bei Altan Uul IV (= Altan Ula IV) und bei Tsagaan Khushuu (= Tsagan Khushu) aufgenommen, wobei an letzterer Lokation die Hangendgrenze der Formation aufgeschlossen ist. Weitere Vorkommen innerhalb des Nemegt-Beckens liegen bei Altan Uul II und III, Naran Bulag (= Naran Bulak), Ulaan Bulag (= Ulan Bulak).[3] Die Vorkommen bei Bügiin Tsav (= Bugeen Tsav = Bugin Tsav = Bugin Cav), Khaichin Uul (= Khaichin Ula) und Guriliin Tsav im Nordwesten sowie bei Hermiin Tsav (= Khermeen Tsav) im Südwesten außerhalb des eigentlichen Nemegt-Beckens werden aufgrund ihres Fossilinhalts entweder als Teil der Nemegt-Formation oder als deren zeitgleiches Äquivalent betrachtet.[3][5]
Als Leitfossilien der Formation werden der Tyrannosauride Tarbosaurus bataar, der Hadrosaurier Saurolophus angustirostris, der Ornithomimosaurier Gallimimus bullatus, der Ankylosaurier Tarchia teresae (= Tarchia gigantea = Dyoplosaurus giganteus) sowie die beiden Gastropoden Mesolanistes efremovi und Mesolanistes mongoliensis genannt.[3]
Stratigraphie und Alterszuordnung

Sowohl die Baruungoyot- als auch die Nemegt-Formation lassen sich stratigraphisch noch weiter untergliedern. In der Gesamtabfolge werden sieben stratigraphische Zonen unterschieden, wobei die Zonen 1–3 ausschließlich die Baruungoyot-Formation betreffen.[2]
Als Zone 4 („Übergangszone“) wird die Faziesverzahnung mit der Baruungoyot-Formation im Liegenden der Nemegt-Formation bezeichnet. Sie besteht im Wesentlichen aus fluviatilen (Nemegt-Formation) und äolischen (Baruungoyot-Formation) Sedimenten. Diese Übergangszone ist an den Lokationen Nemegt und Hermiin Tsav aufgeschlossen.[2]
Die Zone 5 („Untere Nemegt-Formation“) wird hauptsächlich von fluviatilen Sandsteinen aufgebaut. Ton-/Schluffsteine treten nur untergeordnet auf. Typische Vorkommen sind, neben Nemegt und Hermiin Tsav, vor allem Altan Uul III und IV.[2]
In der Zone 6 („Mittlere Nemegt-Formation“) treten neben den fluviatilen Sandsteinen auch verstärkt Ton-/Schluffsteine und Paläoböden in Erscheinung. Typische Profile dieser Zone sind bei Altan Uul II und bei Bügiin Tsav aufgeschlossen.[2]
Die Zone 7 („Obere Nemegt-Formation“) besteht hingegen wiederum hauptsächlich aus fluviatilen Sandsteinen. Typische Vorkommen sind ebenfalls bei Altan Uul II und Bügiin Tsav zu finden.[2]
Für die Nemegt-Formation liegen, aus Mangel an dafür geeigneten Lithologien, keine absoluten radiometrischen Altersdaten vor.[6] Marine Leitfossilien, die eine globale Korrelation ermöglichen würden, fehlen in der Nemegt-Formation ebenso wie in der gesamten mesozoischen Abfolge im Gebiet der heutigen Gobi.[7] Die zeitliche Zuordnung in das oberste Campanium bis unteren/mittleren Maastrichtium basiert im Wesentlichen auf Vergleiche mit der besser datierbaren Wirbeltierfauna der Oberkreide Nordamerikas. Der Hadrosaurier Saurolophus angustirostris nimmt diesbezüglich eine Schlüsselstellung ein, da die Gattung mit Saurolophus osborni sonst nur aus der entsprechend alten Horseshoe-Canyon-Formation in der kanadischen Provinz Alberta bekannt ist.[3][8] Der paläontologische Befund für die Nemegt-Formation wird durch magnetostratigraphische Untersuchungen gestützt.[9]
Palökologie

Die Sedimente der Nemegt-Formation wurden im Bereich eines mäandrierenden und/oder verflochtenen Flusssystems abgelagert. Vor allem in der Mittleren Nemegt-Formation („Zone 6“) lassen die sedimentologischen Befunde zudem Rückschlüsse auf ausgedehnte Seen und Sumpfgebiete zu. Der Ablagerungsraum der Nemegt-Formation war vermutlich mit von Araukarien dominierten Baumbeständen bewaldet. Der paläobotanische Befund wird durch die Analyse der δ13C-Werte aus dem Zahnschmelz von herbivoren und karnivoren Dinosauriern aus der Formation gestützt.[8]
Die Analyse von stabilen Sauerstoffisotopen (δ18O-Werte) aus derselben Quelle weist auf ein kühlgemäßigtes Klima mit durchschnittlichen Jahrestemperaturen von 7,6–8,7 ± 3,9 °C hin. Die Daten lassen zudem Rückschlüsse auf einen jahreszeitlichen Wechsel mit trockenen Wintern und niederschlagsreicheren Sommern zu. Die durchschnittliche Jahresniederschlagsmenge lässt sich mit etwa 775–835 mm angeben.[8]
Abgesehen von den klimatischen Bedingungen lässt sich der Ablagerungsraum der Nemegt-Formation in Kombination mit der Baruungoyot- und der Djadokhta-Formation gut mit den Verhältnissen im rezenten Okavangodelta vergleichen.[10]
Wirbeltierreste finden sich häufig als artikulierte (in anatomischem Zusammenhang stehende) bis semi-artikulierte Skelette oder Teilskelette in den Kies- und Sandablagerungen von Flussinseln oder an den Gleithängen ehemaliger Flussschlingen. Die Unterseite des Skeletts liegt dabei oft in den Konglomeraten an der Basis eines fining-upward Zyklus, während der Rest von zunehmend feinkörnigeren Sanden bedeckt wurde. Die Skelette bipeder Dinosaurier liegen meist auf einer Seite und nehmen eine typische Haltung mit zum Rücken hin zurückgebogenem Hals und Schwanz ein, was darauf hinweist, dass die Kadaver nicht weit transportiert wurden und der Ort der Fossilwerdung in der Nähe des Sterbeortes liegt. Gemeinsam mit den Skelettresten finden sich häufig Hautabdrücke der Tiere, wodurch belegt ist, dass die Einbettung im Sediment in der Regel erfolgte noch bevor der Verwesungsvorgang vollständig abgeschlossen war.[8]

Die Nemegt-Formation ist eine der wenigen Formationen weltweit, in der nicht nur Skelettreste von Dinosauriern, sondern auch deren Spurenfossilien häufig auftreten. Einzelne Trittsiegel oder ganze Fährten lassen sich dadurch leichter ihren Verursachern zuordnen und liefern zusätzliche Informationen zur Palökologie des Ablagerungsraumes.[11][12] Im Gegensatz zu den Skelettresten wurden die Trittsiegel bevorzugt in den feinkörnigen, tonig-schluffigen Sedimenten am Top eines fining-upward Zyklus angelegt, wo sie von den grobkörnigen Sedimenten des nächstfolgenden Zyklus aufgefüllt und meist als konvexes Hyporelief erhalten blieben. Die feinkörnigen Sedimente in denen die Trittsiegel angelegt wurden entsprechen den Ablagerungen der Überschwemmungsebene oder Altwassern. Mehrere solcher Horizonte aus unterschiedlichen Ablagerungszyklen der Lokation Nemegt zeigen neben gut erhaltenen Trittsiegeln, zum Teil mit Hautabdrücken, auch Spurenfossilien von Wirbellosen. Trockenrisse konnten hingegen nicht festgestellt werden. Dies weist darauf hin, dass der nächste Überschwemmungszyklus bereits einsetzte noch bevor die Sedimente des vorhergehenden Zyklus vollständig ausgetrocknet waren.[11]
Abschätzungen der Zusammensetzung der Dinosaurier-Fauna der Nemegt-Formation auf Basis von Knochenfunden und auf Basis von Spurenfossilien liefern unterschiedliche Ergebnisse. Die Skelkettfunde alleine, für sich gesehen, deuten in Bezug auf die Anzahl an Einzelindividuen eine deutliche Dominanz des großen Karnivoren Tarbosaurus bataar gegenüber seinen potenziellen Beutetieren wie Hadrosauriern oder Sauropoden an. Der Befund wirft einige Fragen auf, da eine solche Faunenzusammensetzung keinem stabilen Ökosystem entspricht. Die Auswertung von Spurenfossilien zeichnet dagegen ein anderes Bild mit einer deutlichen Dominanz von Hadrosauriern, gefolgt von Sauropoden und eher selten auftretenden großen Theropoden, wie Tarbosaurus bataar.[11][12][13][14]
Die Ursachen für die unterschiedlichen Befunde sind noch nicht zur Gänze geklärt. Diskutiert werden unter anderem die Möglichkeiten, dass die großen Karnivoren empfindlicher auf regelmäßig auftretende, widrige Witterungsbedingungen reagierten, dass die großen Karnivoren sehr effektiv bei der Beseitigung der Kadaver herbivorer Dinosaurier waren und deshalb deren Wahrscheinlichkeit einer Fossilerhaltung wesentlich geringer war oder, dass es im Ablagerungsraum der Nemegt-Formation saisonal zu Massenansammlungen von Tarbosaurus kam, die mit besonders günstigen Bedingungen für eine Fossilerhaltung zusammenfielen. Allerdings liegen für keine der vorgeschlagenen Hypothesen Belege vor, mit denen sie gestützt oder widerlegt werden könnten.[13]
Forschungsgeschichte
Bereits in den 1920er-Jahren hatten mehrere Expeditionen des American Museum of Natural History unter der Leitung von Roy Chapman Andrews in der Wüste Gobi reiche Fossillagerstätten entdeckt. Die Funde neuer Dinosaurierarten und deren Eier erregten weltweit Aufmerksamkeit, die paläontologische Erforschung des Gebietes kam ab 1930 jedoch praktisch vollständig zum Erliegen.[13][15] Nahezu zeitgleich begannen ab 1925 Geologen der Akademie der Wissenschaften der UdSSR im Gebiet der damaligen Mongolischen Volksrepublik nach Bodenschätzen zu suchen. Dabei wurden unter anderem auch Fundstellen kreidezeitlicher Dinosaurier im Osten des Landes untersucht. Teams der Staatlichen Universität des Fernen Ostens in Wladiwostok waren ebenfalls hauptsächlich im zentralen und östlichen Teil der Mongolei tätig.[16]
Sowjetisch-Mongolische Paläontologische Expeditionen (1946–1949)
Ende 1940 schickte das Wissenschaftliche Komitee der Mongolischen Volksrepublik eine Einladung an das Paläontologische Institut (PIN) der Akademie der Wissenschaften der UdSSR, mit dem Vorschlag im Jahr 1941 eine paläontologische Expedition in der Mongolei durchzuführen. Dem Vorschlag wurde von sowjetischer Seite zugestimmt. Juri Alexandrowitsch Orlow und Iwan Antonowitsch Jefremow waren als Expeditionsleiter und stellvertretender Expeditionsleiter vorgesehen. Die Expedition sollte mit insgesamt 10 Personen ab Ende Mai 1941 für etwa 3,5–4 Monate den Süden und Westen der Mongolei sowie die Trans-Altai- und die Mittlere Gobi erkunden. Da sich die Ausstellung der erforderlichen Reisepässe verzögert hatte, wurde die Expedition auf das Jahr 1942 verschoben. Am 22. Juni 1941 begann mit dem Überfall der deutschen Wehrmacht auf die Sowjetunion allerdings der Deutsch-Sowjetische Krieg und die Expeditionspläne mussten endgültig auf unbestimmte Zeit verschoben werden.[16]
Bereits wenige Monate nach Kriegsende, noch Ende des Jahres 1945, beantragte Orlow eine Wiederaufnahme der Expeditionsvorbereitungen, die im Frühjahr 1946 bewilligt wurde. Die Expedition war nun auf eine Dauer von sieben Monaten ausgelegt. Jefremow wurde mit der Expeditionsleitung beauftragt und Orlow selbst übernahm eine Funktion als wissenschaftlicher Berater. Das Team verließ Moskau Anfang August 1946, erreichte am 10. August Ulaanbaatar und begann am 1. September mit der Geländearbeit. Bis zu ihrer Rückkehr nach Ulaanbaatar am 4. November legten die Fahrzeuge der Expedition etwa 4700 km zurück. Dabei wurden im Nemegt-Becken südlich des Nemegt-Massivs auch die fossilreichen Fundstellen von Nemegt und Altan Uul entdeckt.[16]

Nach ihrer Rückkehr nach Moskau im Januar 1947 begannen die russischen Wissenschaftler sofort mit den Vorbereitungen einer zweiten Expedition. Noch im November des gleichen Jahres wurden Fahrzeuge und Ausrüstung nach Ulaanbaatar gebracht. Im Dezember folgten der Expeditionsleiter Jefremow und sein wissenschaftlicher Berater Orlow. Die Wintermonate wurden für den Aufbau einer Expeditionsbasis in Dalandsadgad genutzt. Während die erste Expedition von 1946 im Wesentlichen der Vorerkundung gedient hatte, sollten diesmal gezielte Grabungen in zwei ausgewählten Gebieten, südlich von Sainschand im Dorno-Gobi-Aimag und im Nemegt-Becken im Ömnö-Gobi-Aimag, durchgeführt werden. Die Grabungsarbeiten begannen im März 1948 im Dorno-Gobi-Aimag und verlagerten sich ab Anfang Mai zur Nemegt-Fundstelle im Ömnö-Gobi-Aimag, wo neben den Überresten des großen Hadrosauriers Saurolophus angustirostris auch zahlreiche Fossilbelege für weitere Dinosaurier, große Schildkröten, Krokodile und Fische geborgen werden konnten. Der spektakulärste Fund gelang jedoch in der benachbarten Lokation Altan Uul II, die in einem Bonebed, das später als „Dragon's Tomb“ („Drachengrab“) berühmt wurde, vollständige Skelette von Tarbosaurus und Saurolophus, zum Teil noch mit fossilen Hautabdrücken lieferte.[16]
Die Arbeiten im Nemegt-Becken dauerten während des ganzen Sommers 1948 an und wurden ab August 1949 im Rahmen der dritten Sowjetisch-Mongolischen Paläontologischen Expedition an den Fundstellen Nemegt, Altan Uul und Tsagaan Khushuu (= Tsagaan Uul) fortgesetzt. Für das Jahr 1950 war eine vierte Expedition vorgesehen. Im Mai dieses Jahres befanden sich Fahrzeuge und Ausrüstung bereits in Ulaanbaatar, wurden dort aber völlig überraschend für ein landwirtschaftliches Projekt abgezogen. Die Hintergründe für den Abbruch der Expedition sind bis heute nicht restlos geklärt. Vermutet werden diplomatische Verwicklungen in Zusammenhang mit der Gründung der Volksrepublik China am 1. Oktober 1949.[16]
Polnisch-Mongolische Paläontologische Expeditionen (1963–1971)

Die Idee einer gemeinschaftlichen Polnisch-Mongolischen Paläontologischen Expedition (PMPE) wurde im März 1961 bei einem Treffen von Repräsentanten der „wissenschaftlichen Akademien der Volksrepubliken“ in Warschau geboren. Initiator war der polnische Paläontologe Roman Kozłowski. Im September 1961 reiste Kozłowski mit einer offiziellen Delegation der Polnischen Akademie der Wissenschaften (PAN) nach Ulaanbaatar und unterzeichnete ein Kooperationsabkommen mit der Akademie der Wissenschaften der Volksrepublik Mongolei. Das Abkommen sah mehrere gemeinschaftliche Expeditionen über einen Zeitraum von drei Jahren vor. Auf polnischer Seite wurden Organisation und wissenschaftliche Leitung der Paläontologin Zofia Kielan-Jaworowska übertragen, die 1965 auch die Leitung des polnischen Expeditionsteams übernahm. Auf mongolischer Seite war Naydin Dovchin von 1963 bis 1965 für Organisation und Expeditionsleitung verantwortlich.[17]
Die erste Expedition 1963 diente, ebenso wie bereits die erste sowjetische Expedition 1946, hauptsächlich der Vorerkundung. Arbeiten im Nemegt-Becken wurden ab Anfang August 1963 aufgenommen. Von den damals bekannten Aufschlüssen der Nemegt-Formation, Nemegt, Altan Uul und Tsagaan Khushuu, wurde nur Letzterer besucht und es konnten zahlreiche Fragmente von Dinosaurierknochen und Schildkrötenpanzern geborgen werden.[17]
Gezielte Grabungen im Nemegt-Becken wurden erst ab der zweiten Expedition 1964 durchgeführt. Am 19. Juni begannen Grabungen bei Tsagaan Khushuu, die die Überreste von zwei Individuen von Tarbosaurus sowie zwei Skelette von Vertretern der Ornithomimidae lieferten. Zudem wurde eine Fundschicht mit zahlreichen Schildkrötenpanzen, Überresten von Dinosauriern, Krokodilen, Fischen, Muscheln und fossilen Hölzern entdeckt. Parallel zu den Grabungen erkundeten kleinere Trupps des Expeditionsteams die weitere Umgebung. Bei diesen Ausflügen wurde auch die ehemalige Grabungsstelle am „Dragon's Tomb“ (Altan Uul II) und weitere Vorkommen der Nemegt-Formation (Altan Uul III, Altan Uul IV) gefunden. Die Ausflüge erbrachten unter anderem auch zwei weitere Schädel von Tarbosaurus und das Schwanzfragment eines großen Ankylosauriers. Am 12./13. Juli wurde das Basislager von Tsagaan Khushuu zur Nemegt-Lokation verlegt, wo ein weiteres Skelett eines Tarbosaurus geborgen werden konnte.[17]

Während der dritten Expedition 1965 wurde das Team zunächst in zwei Gruppen geteilt. Eine Gruppe erkundete die Djadokhta-Formation (= Djadochta-Formation) bei Bajandsag, während die andere sich wieder dem Nemegt-Becken zuwandte. Bei Altan Ula IV wurde das nahezu vollständige postcraniale Skelett eines großen Sauropoden entdeckt und bei der Nemegt-Lokation der isolierte Schädel eines weiteren Sauropoden.[17] Die beiden Funde wurden später als Opisthocoelicaudia skarzynskii und Nemegtosaurus mongoliensis beschrieben.[18] Zu den weiteren Funden zählten erneut mehrere Individuen von Tarbosaurus, ein kleiner Vertretern der Ornithomimidae, das Teilskelett eines großen Ankylosauriers sowie der Schultergürtel und die vorderen Gliedmaßen eines unbekannten, vermeintlich karnivoren Dinosauriers,[17] der 1970 als Deinocheirus mirificus beschrieben wurde.[19]
Für das Jahr 1966 wurden die gemeinsamen polnisch-mongolischen Expeditionen ausgesetzt. Von 1967 bis 1969 wurden nur kleinere Expeditionen durchgeführt, die sich ausschließlich auf die Djadokhta-Formation bei Bajandsag konzentrierten. Erst für die Jahre 1970 und 1971 waren wieder zwei größere Expeditionen geplant, bei denen auch wieder Grabungen im Nemegt-Becken durchgeführt werden sollten. Die Kosten für die beiden Expeditionen wurden diesmal ausschließlich von polnischer Seite getragen. Die Leitung der jeweils 12-köpfigen polnischen Teams wurde erneut von Kielan-Jaworowska übernommen. Als Leiter des jeweils drei-köpfigen mongolischen Teams wurde Rinchen Barsbold benannt, der jedoch an keiner der beiden Expeditionen aktiv teilnahm, sondern lediglich 1970 die Grabungsstellen zweimal kurz besuchte.[20]
Neben der Suche nach weiteren Wirbeltierfossilien in der Nemegt-Formation lag ein Schwerpunkt der beiden Expeditionen auch in der weiteren Erforschung der stratigraphischen Verhältnisse der Sedimente des Nemegt-Beckens und in der Gewinnung von Proben für mikropaläontologische Untersuchungen.[20]
Abgesehen von den zahlreichen Fossilfunden liegt die besondere Bedeutung der polnisch-mongolischen paläontologischen Expeditionen in der Aufschlüsselung der stratigraphischen Abfolge der Sedimente des Nemegt-Beckens und der Erstellung von Kartenskizzen der einzelnen Lokationen. Das Kartenmaterial, das den Wissenschaftlern zur Verfügung stand, war für ihre Zwecke völlig ungeeignet. Um sich zu behelfen, schlichteten sie bei jeder Lokation an strategisch günstigen Stellen Steinmänner als Vermessungsmarken auf, wobei darauf geachtet wurde, dass jeder Steinmann von zumindest zwei anderen aus eingesehen werden konnte.[13] Kleinere Steinmänner dienten als Markierung für einzelne Fundstellen.[18] Trianguliert wurde mit dem Geologenkompass. Die ersten dieser Kartenskizzen wurden bereits 1969 veröffentlicht[21] und dienten als Arbeitsgrundlage für zahlreiche weitere Expeditionen. Insbesondere die Kartenskizzen der Lokationen Altan Uul IV und Nemegt wurden noch 2018 von Philip J. Currie und Co-Autoren als „geschickt gezeichnet“ und „bemerkenswert genau“ gewürdigt.[18]
Sowjetisch/Russisch-Mongolische Paläontologische Expeditionen (ab 1969)
Ab Ende 1960 setzte sich Orlow erneut bei der Akademie der Wissenschaften der UdSSR um die Bewilligung erdwissenschaftlicher Expeditionen in der Mongolei ein. 1964 erhielten zwei sowjetische Paläontologen Gelegenheit sich an der Erkundung einer kurz zuvor von Einheimischen entdeckten Fossilfundstelle bei Bügiin Tsav zu beteiligen.[16] Im Rahmen dieser kleinen Expedition konnten aus Schichten, die später der Nemegt-Formation zugeordnet wurden, auch die Überreste eines kleinen Theropoden (Saurornithoides junior = Zanabazar junior) geborgen werden.[22]
Aufgrund bürokratischer und politischer Probleme kam die erste reguläre Sowjetisch-Mongolische Geologische Expedition erst 1967, im Jahr nach Orlows Tod, zustande.[16] Dabei wurde unter anderem auch die Nemegt-Lokation besucht. Erst ab 1969 gab es auch wieder eigenständige Sowjetisch-Mongolische Paläontologische Expeditionen (SMPE). Seitdem waren nahezu jedes Jahr während der Grabungssaison entsprechende Expeditionen in der Mongolei tätig. Die Geländearbeiten wurden auch nach dem Zusammenbruch der Sowjetunion weitgehend ungehindert fortgesetzt und ab 1992 als Russisch-Mongolische Paläontologische Expeditionen (RMPE) bezeichnet.[15]
Bereits während einer der ersten dieser Expeditionen wurde 1969 oder 1970 die Lokation Hermiin Tsav entdeckt,[15] deren hangendste Schichtglieder („upper white beds“) der Nemegt-Formation zugeordnet werden.[3]
Expeditionen ab 1990
Nach dem Zusammenbruch der Mongolischen Volksrepublik und der Umwandlung in den Mongolischen Staat im Jahr 1990 zeigten auch westliche Institutionen wieder Interesse an einer gemeinschaftlichen Erforschung der Nemegt-Formation. Das American Museum of Natural History (AMNH) führte seit 1990 gemeinsam mit der Mongolischen Akademie der Wissenschaften nahezu jährlich Expeditionen durch, die sich zunächst hauptsächlich auf die Nemegt- und die Baruungoyot-Formation konzentrierten. Nach der Entdeckung der Fossilvorkommen von Ukhaa Tolgod verlagerte das AMNH seine Aktivitäten ab 1994 jedoch zunehmend an diese Fundstelle der Djadokhta-Formation. Im Rahmen einer der wenigen Ausnahmen konnte 2001 das weitgehend vollständige Skelett eines Alioramus von der Fundstelle Tsagaan Khushuu innerhalb der Nemegt-Formation geborgen werden.[15][23]
Das von einer japanischen Biochemie-Firma privat betriebene Hayashibara Museum of Natural Sciences in Okayama pflegte ebenfalls gute Beziehungen zur Mongolischen Akademie der Wissenschaften. Von 1993 bis 2008 wurde eine ganze Serie von Expeditionen durchgeführt,[24] die sich in Bezug auf die Nemegt-Formation hauptsächlich mit der Geologie und Paläontologie der Lokationen Bügiin Tsav und Hermiin Tsav beschäftigten.[15] Zu den bemerkenswertesten Funden dieser Expeditionen zählen das Nest eines Oviraptoriden mit in den Eiern noch erhaltenen Skelettresten der Embryonen[25] und ein weitgehend vollständiges Skelett eines Jungtieres von Tarbosaurus.[26]

Kanadische Wissenschaftler des Royal Tyrrell Museum of Palaeontology in Drumheller und der University of Alberta in Edmonton kooperierten von 1996 bis 2007 und 2011[13] in zahlreichen Expeditionen mit der Mongolischen Akademie der Wissenschaften. Die Expeditionen wurden vom privaten Reiseveranstalter Nomadic Expeditions unterstützt und wurden unter der Bezeichnung „Dinosaurs of the Gobi“ bekannt. Nach 1999 waren diese Expeditionen überwiegend in der Nemegt-Formation bei Altan Uul, Bügiin Tsav, Hermiin Tsav, Nemegt, Tsaagan Khushuu und Ulaan Khushuu tätig.[15] Ziele dieser Expeditionen waren unter anderem auch eine genauere Erfassung der stratigraphischen Abfolge der Nemegt-Formation und deren Beziehung zur Baruungoyot-Formation,[4][2] sowie die Erstellung einer Datenbank mit der genauen Lage und stratigraphischen Position aller bekannter Dinosaurierfundstellen der Nemegt-Formation.[13] Im Jahr 2002 wurde eine dieser Expeditionen auch wieder von Zofia Kielan-Jaworowska und ihrem Ehemann Zbigniew Jaworowski begleitet.[13]
Sowohl in Bezug auf die Expeditionsziele als auch in Bezug auf die Teilnehmer gingen die „Dinosaurs of the Gobi“-Expeditionen ab 2006 nahezu fließend in die Expeditionen des Koreanisch-Mongolischen Internationalen Dinosaurierprojektes (KID) über.[15] Zu den spektakulärsten Funden dieser Expeditionen zählten 2006 und 2009 zwei Skelettfunde von Deinocheirus mirificus, mit denen, fast 50 Jahre nach der Entdeckung des Holotypus, endlich die systematische Zugehörigkeit dieser Gattung geklärt werden konnte.[27] Ebenfalls 2006 wurde bei der Lokation Nemegt ein von Raubgräbern bereits schwer beschädigtes Bonebed mit den Überresten von mindestens 18 Individuen der Gattung Avimimus gefunden. Nach „Dragon's Tomb“ war dies erst der zweite Nachweis für ein monodominantes Dinosaurier-Bonebed aus der Nemegt-Formation.[28]
Unwesen durch Raubgräber und Schmuggler
_officials_return_a_Tarbosaurus_bataar_skeleton_to_the_government_of_Mongolia_during_a_repatriation_ceremony_May_6%252C_2013%252C_at_a_Manhattan_hotel_in_New_York_130506-H-ZZ999-002.jpg.webp)
Die Verwüstung von Fundstellen der Nemegt-Formation durch illegale Raubgräber war während des 20. Jahrhunderts kein besonderes Problem. Im Jahr 2000 wurde erstmals festgestellt, dass sechs Skelette von Tarbosaurus von der Lokation Bügiin Tsav von Raubgräbern zerstört worden waren. Die Skelette waren bereits während der einheimischen Expedition von 1964 entdeckt, aber nicht geborgen, sondern von Regierungsseite lediglich unter Schutz gestellt worden. In den folgenden zehn Jahren eskalierte die Situation auf alarmierende Weise. 2013 berichteten Philip J. Currie und seine Ehefrau Eva Koppelhus von Zerstörungen durch Raubgräber an 98 Skeletten von Tyrannosauriden, 86 Ornithomimidae, 53 Hadrosauriern, 32 Sauropoden, 20 Ankylosauriern und mehr als 20 anderen Dinosaurierfunden.[29]
Die Raubgräber hatten es in der Regel nur auf jene Skelettteile abgesehen, die sich am leichtesten verkaufen lassen, wie etwa einzelne Zähne und Krallen, bezahnte Kiefer oder ganze Schädel. Um an diese Teile zu gelangen wurde jedoch häufig die weitgehende Zerstörung des ganzen Skeletts in Kauf genommen. Die Fossilien wurden vermutlich mit regulären Kohletransporten nach China außer Landes geschmuggelt und von dort aus auf den internationalen Markt gebracht.[29]

Dass Raubgräber ganze Skelette aus der Nemegt-Formation bargen, blieb dagegen die Ausnahme. Eine dieser Ausnahmen führte in den USA zu einem Gerichtsprozess, der nicht nur in paläontologischen und juristischen Fachkreisen, sondern auch in den Medien für einige Aufmerksamkeit sorgte. Der Rechtsfall wurde unter der Bezeichnung „United States v. One Tyrannosaurus Bataar (sic!) Skeleton“ („[Die] Vereinigten Staaten gegen Ein Tyrannosaurus Bataar Skelett“) bekannt.[30] Im Frühjahr 2012 hatte ein New Yorker Auktionshaus das weitgehend vollständige Skelett eines großen Tyrannosauriden medienwirksam zur Versteigerung angeboten. Die mongolischen Paläontologin Bolortsetseg Minjin erkannte den spekatulären Auktionsposten als Tarbosaurus bataar und informierte ihre Regierung in Ulaanbaatar.[31] Die mongolische Regierung beantragte darauf hin bei den US-amerikanischen Behörden eine einstweilige Verfügung zur Unterbindung der Auktion. Dem Antrag wurde stattgegeben; die Auktion fand dennoch statt und das Skelett wurde für mehr als 1 Million US-Dollar versteigert. Der damalige Präsident der Mongolei, Tsachiagiin Elbegdordsch wandte sich persönlich an das United States Attorney’s Office for the Southern District of New York und bat um rechtlichen Beistand in der Angelegenheit. Auf Antrag des Präsidenten wurde eine Expertenkommission damit beauftragt das Skelett genauer zu untersuchen. Diese bestätigte Minjins Verdacht und erklärte, dass das Skelett zweifellos aus der Nemegt-Formation in der Mongolei stammte.[30][32] Das Skelett wurde von der United States Immigration and Customs Enforcement (ICE) beschlagnahmt. Im Oktober des gleichen Jahres wurde ein in Florida ansässiger Fossilienhändler, der das Skelett, mit falschen Angaben auf den Einfuhrpapieren, in die USA gebracht hatte, verhaftet. Im Mai 2013 wurde das Tarbosaurus-Skelett formell an die Mongolei zurückgegeben.[32]
Die Ermittlungen durch das ICE gingen unterdessen weiter und bis Juli 2014 wurden mehr als 18 aus der Mongolei stammende Dinosaurierskelette aus den Lagerbeständen des angeklagten Händlers beschlagnahmt und an die Mongolei zurückgegeben. Darunter befanden sich zwei weitere Skelette von Tarbosaurus, zwei Saurolophus angustirostris und vier Gallimimus-Skelette, die sich als Leitformen eindeutig der Nemegt-Formation zuordnen lassen.[32] Im Dezember 2015 kam noch ein weiterer Schädel eines Tarbosaurus dazu, der bereits 2007 bei einer Auktion in Manhattan von einem privaten Sammler für 276.000 US-Dollar erworben worden war. In den Medien wurde kurz darauf berichtet, dass es sich bei dem Käufer um den Hollywood-Schauspieler Nicolas Cage handelte, der das Fossil in Unkenntnis seiner illegalen Herkunft erworben und bei der Auktion angeblich auch noch seinen Schauspielkollegen Leonardo DiCaprio überboten hatte.[33][34]
Im Dezember 2014 beantragte die Mongolei die Aufnahme von drei typischen Lokationen der Nemegt-Formation (Nemegt, Hermiin Tsav und Bügiin Tsav) in die Weltnaturerbeliste der UNESCO.[35] Parallel dazu erstellten Wissenschaftler einen geochemischen Fingerabdruck für Dinosaurierfossilien aus der Nemegt-Formation, der es ermöglichen sollte illegal geborgene und geschmuggelte Fossilien eindeutig der Nemegt-Formation zuweisen zu können. Den entscheidenden Hinweis zur Entwicklung dieser Methode lieferte Zofia Kielan-Jaworowskas Ehemann Zbigniew Jaworowski. Der Radiologe hatte bereits in den 1960er-Jahren festgestellt, dass die fossilen Knochen aus der Nemegt-Formation einen ungewöhnlich hohen Gehalt an Uran aufwiesen.[36]
Paläontologie
Flora
Fossile Hölzer zählen zu den häufigeren Funden aus der Nemegt-Formation. Einzelne Stämme erreichen eine Länge von 8 m und einen Durchmesser von bis zu 30 cm. Die Hölzer lassen sich meist den Araukariengewächsen zuordnen.[3][8] Neben den zahlreichen Belegen für Araukariengewächse liegt mit Nyssoidea mongolica von der Lokation Bügiin Tsav auch ein Nachweis für einen Vertreter der Tupelogewächse vor.[37]
Feinkörnige Sedimente von der Lokation Ulaan Bulag (= Ulan Bulak) zeigen eine reiche Flora von Wasserpflanzen mit Laichkräutern, der Brachsenkräuter-ähnlichen Bärlapppflanze Monilitheca und Samen von Wasserlinsengewächsen. Derselbe Fundort lieferte auch zahlreiche Abdrücke der Halme und Blätter von Sauergrasgewächsen.[38]
Die systematische Beprobung der Nemegt-Formation nach Mikrofossilien durch die Polnisch-Mongolischen Expeditionen von 1970 und 1971 erbrachte unter anderem auch eine reiche Ausbeute an spiralig skulpturierten Oogonien (Gyrogoniten) von Charophyten. Mindestens 34 Arten wurden unterschieden, wobei die meisten davon den Gattungen Mongolichara und Mesochara zugeordnet werden, die gleichzeitig auch die größte Anzahl an Einzelfunden stellen.[39]
Fungi
Eine Besonderheit ist der Fund mehrerer Fruchtkörper eines den Erdsternen ähnlichen Ständerpilzes von der Lokation Tsagaan Khushuu, die 2003 als Geastroidea lobata beschrieben wurden. Die fossilen Pilze sind auf einer Schichtfläche in feinkörnigen Sedimenten gemeinsam mit reichlichem Pflanzenstreu und den Früchten von Wasserlinsengewächsen erhalten. Unter dem Rasterelektronenmikroskop konnte auf der Schichtfläche ein ganzes Netz von Pilzhyphen festgestellt werden.[40]
Mycelien von Pilzen oder den pilzähnlichen Saprolegnia konnten auch in den Langknochen von mehreren Individuen des Ornithomimosauriers Gallimimus bullatus von unterschiedlichen Lokationen der Nemegt-Formation nachgewiesen werden. Die mycelbildenden Organismen besiedelten post mortem die Markhöhle der Knochen und drangen über die Osteone auch in das angrenzende Knochengewebe vor.[41]
Wirbellose
Neben den bereits als Leitformen der Nemegt-Formation genannten Gastropoden der Gattung Mesolanistes aus der Familie der Apfelschnecken ist mit der Gattung Hydrobia auch ein Vertreter der Wasserdeckelschnecken belegt. Süßwassermuscheln sind mit jeweils mehreren Arten der Gattungen Buginella und Pseudohyria vertreten.[3]
Phyllopoda sind mit mehreren Arten und Gattungen nachgewiesen.[3] Aufgrund der gezielten mikropaläontologischen Probenahmen durch die Polnisch-Mongolischen Paläontologischen Expeditionen ist umfangreiches Belegmaterial für Süßwasser-Ostrakoden vorhanden. In lokalen Massenansammlungen können die Ostrakoden-Schalen nahezu gesteinsbildenden Charakter annehmen. 34 Arten aus 22 Gattungen wurden 1978 beschrieben. Fünf Gattungen, Altanicypris, Mongolocypris, Talicypridea, Gobiella und Khandia waren zuvor unbekannt und belegen einen hohen Anteil an endemischen Formen. Mengenmäßig wird die Ostrakoden-Fauna der Nemegt-Formation jedoch von Vertretern der Gattung Cypridea dominiert.[42] Die Gattung war in der Unterkreide weltweit verbreitet, verschwand gegen Ende der Unterkreide aber in fast allen Regionen. Nur in den intrakontinentalen Becken Chinas und der Mongolei konnte sie sich bis zum Ende der Oberkreide halten.[42][43]
Fische
Fossilreste von Fischen sind keine Seltenheit in den Sedimenten der Nemegt-Formation, beschränken sich in der Regel jedoch auf einzelne Wirbel und Kieferfragmente. Einige dieser Funde konnten der Familie der Hiodontidae zugeordnet werden, die rezent nur noch mit einer Gattung und zwei Arten in Seen und Flüssen Nordamerikas bekannt ist.[44]
Amphibien
Altanulia alifanovi wurde 1993 auf Basis eines etwa 2 cm langen Oberkieferfragments von der Lokation Altan Uul II als Vertreter der Scheibenzüngler innerhalb der Anura beschrieben. Die Zuordnung des Fossils zu den Froschlurchen wurde von späteren Autoren allerdings in Zweifel gezogen.[45]
Schildkröten

Überreste von Schildkröten zählen zu den häufigsten Wirbeltierfossilien der Nemegt-Formation. Lagenweise Massenansammlungen von Schildkrötenpanzern wurden von mehreren Lokationen beschrieben.[37] In diesen Massenansammlungen sind die einzelnen Individuen teilweise so dicht gepackt, dass sich ihre Panzer gegenseitig überlagern oder regelrecht ineinander verkeilt sind.[46]
In Bezug auf die Individuenanzahl wird die Schildkrötenfauna der Nemegt-Formation von Mongolemys elegans dominiert.[46] Von dem, an ein Leben im Süßwasser angepassten, Vertreter der ausgestorbenen Gruppe der Lindholmemydidae sind Exemplare aller Entwicklungsstadien, vom Schlüpfling über noch nicht ausgewachsene Jungtiere bis zu Adulttieren, erhalten.[47] Mit Gravemys barsboldi ist neben Mongolemys elegans noch ein weiterer, allerdings wesentlich seltener auftretender Vertreter der Lindholmemydidae aus der Nemegt-Formation bekannt.[48]
Die artenreichste Schildkrötengruppe der Nemegt-Formation stellen die Weichschildkröten und deren Stammgruppenvertreter (Pan-Trionychidae). Kuhnemys breviplastra gilt als Stammgruppenvertreter der Trionychidae. Die Art ist auch aus der Baruungoyot-Formation bekannt. Nemegtemys conflata wurde als Stammgruppenvertreter der Klappen-Weichschildkröten (Cyclanorbinae) interpretiert und wäre damit einer der ältesten bekannten Nachweise für einen Vertreter der Kronengruppe der Weichschildkröten in der Alten Welt. Zwei weitere Taxa lassen sich noch nicht genauer zuordnen und werden vorläufig als „Trionyx“ gilbentuensis und „Trionyx“ gobiensis im (paläontologischen) Sammeltaxon „Trionyx“ geführt, was jedoch keineswegs eine nähere Verwandtschaft mit der rezenten Gattung Trionyx impliziert. Das ebenfalls aus der Nemegt-Formation beschriebene Taxon Amyda menneri gilt als Nomen dubium.[49]
Mongolochelys efremovi ist dagegen eine große, an das Landleben angepasste Schildkröte. Sie gilt als Vertreter der Sichuanchelyidae, einer Klade aus der Stammgruppe der Schildkröten, die sich sporadisch vom Mitteljura bis ins Paläozän Eurasiens nachweisen lässt.[50][51]
Schuppenechsen
Aus der Baruungoyot- und der Djadokhta-Formation sind Vertreter von insgesamt 15 Familien der Schuppenechsen bekannt. In der Nemegt-Formation sind entsprechende Funde wesentlich seltener und es lassen sich nur Vertreter von vier Familien, den Waranen (Varanidae), den Schienenechsen (Teiidae) und den beiden ausgestorbenen Familien der Mongolochamopidae und der Necrosauridae nachweisen.[52]
Crocodyliformes
Bis 1983 waren aus der Nemegt-Formation mit Shamosuchus ancestralis (= Paralligator ancestralis), Shamosuchus tersus und Shamosuchus ulanicus drei Taxa aus der Gruppe der Crocodyliformes beschrieben worden. Eine spätere phylogenetische Analyse identifizierte alle drei Formen als identisch mit der Art Paralligator gradilifrons, die bereits 1954 aus der etwas älteren Bayanshiree-Formation beschrieben worden war.[53]
Flugsaurier
Der einzige bislang bekannte Fossilbeleg eines Flugsauriers aus der Nemegt-Formation lässt sich möglicherweise einem sehr großen Vertreter der Azhdarchidae zuordnen. Eine genauere Bestimmung auf Gattungs- oder gar Artebene ist jedoch noch nicht möglich.[54]
Dinosaurier

Die Nemegt-Formation gilt, nach der Dinosaur-Park-Formation Kanadas, als lithostratigraphische Einheit mit einer der artenreichsten Dinosaurier-Faunen der Oberkreide.[13]
Die artenreichste Dinosaurier-Gruppe der Nemegt-Formation stellen die Oviraptosauria. Alle drei oberkretazischen Familien der Oviraptosauria, die Avimimidae, die Caenagnathidae und die Oviraptoridae treten auf. Die Avimimidae und die Caenagnathidae sind mit jeweils einer Gattung (Avimimus und Elmisaurus) vertreten.[55] Von den Oviraptoridae sind hingegen mindestens fünf verschiedene Gattungen aus der Nemegt-Formation bekannt. Neben Nemegtomaia, eine Gattung, die auch aus der Baruungoyot-Formation bekannt ist, sind dies Rinchenia, Nomingia (von einigen Autoren auch als möglicher Vertreter der Caenagnathidae interpretiert), Gobiraptor und eine weitere Gattung, die in der Literatur zunächst als „Guriliin Tsav Oviraptoride“ bezeichnet und später als Oksoko avarsan beschrieben wurde.[55][56][57]
Auch die mit den Oviraptosauria verwandten Ornithomimosauria treten in der Nemegt-Formation mit mehreren Gattungen in Erscheinung. Neben den relativ gut bekannten Taxa Gallimimus bullatus, Deinocheirus mirificus und Anserimimus planinychus existierte möglicherweise noch mindestens eine weitere Art dieser Gruppe im Ablagerungsraum der Nemegt-Formation.[58]
Von den Troodontidae, einer weiteren Gruppe der Maniraptoriformes, wurden ebenfalls drei Taxa aus der Nemegt-Formation beschrieben. Die drei Einzelfunde von Saurornithoides junior, Borogovia gracilicrus und Tochisaurus nemegtensis zeigen allerdings keine signifikant überlappenden Skelettelemente und es ist denkbar, dass alle drei Exemplare nur einer einzigen Art (Saurornithoides junior) zugehören.[59] Der Holotypus von Saurornithoides junior wurde 2009 aus der Gattung Saurornithoides ausgegliedert und als Zanabazar junior einer eigenen Gattung zugeordnet.[60] Die Dromaeosauridae, eine Schwesterngruppe der Troodontidae, sind mit Adasaurus mongoliensis vertreten.[61]
Relativ gut belegt ist hingegen das Auftreten von mindestens zwei Formen der Alvarezsauridae (Mononykus olecranus und Nemegtonykus citus) in der Nemegt-Formation. Beide Arten werden innerhalb der Alvarezsauridae der Gruppe der Parvicursorinae zugeordnet.[62] Die Therizinosauroidea sind mit Therizinosaurus cheloniformis vertreten.[61] Damit waren, einschließlich der weiter unten beschriebenen Vögel, alle bekannten Großgruppen der Maniraptoriformes mit mindestens einer Art im Ablagerungsraum der Nemegt-Formation vorhanden.

Abgesehen von den zahlreichen Arten der Maniraptoriformes sind die Theropoden noch mit mehreren Taxa aus der Gruppe der Tyrannosauroidea vertreten.[61] Tarbosaurus bataar, ein naher Verwandter des annähernd gleichzeitig im westlichen Nordamerika lebenden Tyrannosaurus rex,[63] ist in der Nemegt-Formation der größte und gleichzeitig auch am besten bekannte Vertreter dieser Gruppe.[61] Die etwas kleinere Gattung Alioramus ist mit zwei Einzelfunden aus der Nemegt-Formation bekannt, die jeweils eigenen Arten (Alioramus remotus und Alioramus altai) zugeordnet wurden. Es besteht jedoch die Möglichkeit, dass diese beiden Taxa lediglich unterschiedliche ontogenetische Stadien ein und derselben Art repräsentieren.[23] Während Tarbosaurus und Alioramus als Vertreter der Tyrannosauridae innerhalb der Tyrannosauroidea gewertet werden, ist abseits dieser Teilgruppe mit Bagaraatan ostromi ein weiterer Vertreter der Tyrannosauroidea aus der Nemegt-Formation belegt.[61] Unklar ist die Situation in Bezug auf Raptorex kriegsteini. Falls der Holotypus und bislang einzige Fossilbeleg von Raptorex kriegsteini tatsächlich nicht aus der Unterkreide der Jehol-Gruppe, sondern aus der Nemegt-Formation stammt,[44] wäre er der vierte Tyrannosauroide aus dieser Formation. In Summe stellen die Tyrannosauroidea in Bezug auf die Anzahl an Einzelindividuen mehr als ein Drittel aller Dinosaurierfunde aus der Nemegt-Formation und ein Großteil davon lässt sich Tarbosaurus bataar zuordnen.[13]

In Bezug auf Knochenfunde ist, nach Tarbosaurus bataar, der Hadrosaurier Saurolophus angustirostris die zweithäufigste Dinosaurierart der Nemegt-Formation. Mit Barsboldia sicinskii wurde noch eine zweite Hadrosaurierart aus der Formation beschrieben.[13] Von dieser liegt allerdings nur ein einzelnes Teilskelett vor und es ist nicht ganz auszuschließen, dass es sich bei diesem Taxon lediglich um ein sehr großes Exemplar von Saurolophus angustirostris handelt.[13][14]
Überreste großer Sauropoden machen etwa 9 % aller Dinosaurier-Knochenfunde aus der Nemegt-Formation aus.[13] Mit Opisthocoelicaudia skarzynskii und Nemegtosaurus mongoliensis wurden zwei Taxa beschrieben. Da sich das bekannte Skelettmaterial beider Taxa jedoch kaum überschneidet, lässt sich nicht zur Gänze ausschließen, dass Opisthocoelicaudia skarzynskii und Nemegtosaurus mongoliensis derselben Art angehörten und die tatsächliche Anzahl der Sauropoden-Arten der Nemegt-Formation wird nach wie vor kontrovers diskutiert.[18][64]

Die Ankylosauridae stellen mit etwa 7 % aller Dinosaurier-Knochenfunde die nächsthäufigste Gruppe.[13] Auch in diesem Fall ist die tatsächliche Anzahl von in der Nemegt-Formation vertretenen Arten unklar. Der ursprünglich als für die Nemegt-Formation charakteristisch beschriebene Ankylosaurier „Dyoplosaurus“ giganteus[3] wurde später als Tarchia gigantea in die 1977 mit Tarchia kielanae aus der Baruungoyot-Formation beschriebene Gattung Tarchia gestellt und der Fossilbeleg für Tarchia gigantea um einen weitgehend vollständigen Schädel samt Unterkiefer (PIN 3142/250) aus der Nemegt-Formation von Hermiin Tsav ergänzt. Gleichzeitig wurde bereits angedeutet, dass die beiden Taxa möglicherweise zur selben Art gehörten.[65] Spätere Autoren griffen diese Möglichkeit auf und Tarchia kielanae wurde lange als Juniorsynonym von Tarchia gigantea gewertet. 2014 wurde festgestellt, dass der ursprünglich als „Dyoplosaurus“ giganteus beschriebene Holotypus von Tarchia gigantea keine Merkmale zeigt, die eine eindeutige Unterscheidung von anderen Ankylosauriern zulassen würde. Das Taxon wurde dementsprechend verworfen und Tarchia kielanae wurde als alleinige Art der Gattung reaktiviert. PIN 3142/250 wurde gleichzeitig der ebenfalls 1977 aus der Baruungoyot-Formation beschriebenen Art Saichania chulsanensis zugeordnet.[66] Dieser Interpretation wurde 2017 jedoch teilweise widersprochen und PIN 3142/250 wurde unter der Bezeichnung Tarchia teresae erneut als zweite Art neben Tarchia kielanae in die Gattung Tarchia gestellt.[67] Unabhängig von den taxonomischen Schwierigkeiten sind aus der Nemegt-Formation zwei morphotypen ankylosaurider Schwanzkeulen bekannt, die möglicherweise zwei unterschiedliche Arten dieser Gruppe repräsentieren. Allerdings lässt sich keiner der beiden Typen eindeutig einem der infrage kommenden Taxa zuordnen.[66]

Funde von Vertretern der Pachycephalosauria sind in der Nemegt-Formation relativ selten.[13] Mit Prenocephale prenes und Homalocephale calathocercos sind mindestens zwei Arten aus unterschiedlichen Gattungen in der Formation belegt. Die wenigen Funde aus der Nemegt-Formation zählen allerdings zu den am besten erhaltenen und vollständigsten Belegen für die ganze Gruppe und waren mit ausschlaggebend für die Etablierung einer eigenen Unterordnung Pachycephalosauria innerhalb der Vogelbeckensaurier.[68]
Trotz des allgemein hohen Artenreichtums scheinen Vertreter der Neoceratopsia in der Nemegt-Formation vollständig zu fehlen, obwohl die Gruppe mit den Protoceratopsidae sowohl aus der Djadokhta- als auch aus der Baruungoyot-Formation bekannt ist.[69]
Vögel
Im Gegensatz zu den Dinosauriern sind Fossilfunde von Vögeln aus der Nemegt-Formation selten und bruchstückhaft. Dies gilt auch für die Baruungoyot- und die Djadokhta-Formation. Die wenigen Funde aus allen drei Formationen deuten jedoch darauf hin, dass sich die Avifauna der Nemegt-Formation von der der beiden älteren Formationen unterschied. Während in den beiden älteren Formationen Vertreter der Enantiornithes überwiegen, sind aus der Nemegt-Formation verhältnismäßig mehr Funde von Vertretern der Ornithuromorpha, zu denen auch die Modernen Vögel zählen, bekannt.[70]
Gurilynia nessovi ist der einzige bekannte Vertreter der Enantiornithes aus der Nemegt-Formation.[71] Fossile Eierschalen der Ootaxa Laevisoolithus und Subtiliolithus werden ebenfalls den Enantiornithes zugeschrieben.[70] Von Letzteren sind aus der Nemegt-Formation nur Fragmente erhalten, die in einem Fall in einer Anhäufung mit einem Durchmesser von etwa 30 cm, möglicherweise einem Gelege, angereichert sind.[72]
Die Hesperornithiformes, eine ausgestorbene Gruppe von zahntragenden, meist flugunfähigen und an eine schwimmende und tauchende Lebensweise angepassten Vögeln innerhalb der Ornithuromorpha, sind mit mindestens zwei Spezies, Judinornis nogontsavensis und Brodavis mongoliensis vertreten.[73] Die Modernen Vögel sind mit Teviornis gobiensis, einem Vertreter der Gänsevögel, belegt.[71]
Säugetiere
Sowohl von der Baruungoyot-Formation als auch von der etwas älteren Djadokhta-Formation sind zahlreiche Funde von Säugetieren aus der Gruppe der Multituberculata bekannt, die sich alle einer gemeinsamen Überfamilie Djadochtatherioidea zuordnen lassen.[74] Von der Nemegt-Formation liegen dagegen nur zwei Säugetierfunde vor.
Buginbaatar transaltaiensis wurde 1969 von der Lokation Khaichin Uul in der Umgebung von Bügiin Tsav beschrieben.[75] Im Gegensatz zu den Funden aus der Baruungoyot- und der Djadokhta-Formation lässt sich Buginbaatar jedoch nicht den Djadochtatherioidea zuordnen, sondern ist vermutlich nahe verwandt mit den Cimolomyidae, die sonst nur von der Oberkreide Nordamerikas bekannt sind.[74]
Der zweite Säugetierfund stammt von der Lokation Guriliin Tsav und umfasst den Schädel eines noch nicht benannten Vertreters der Metatheria. Das Fossil wird in der Literatur schlicht als „Schädel von Guriliin Tsav“ („Guriliin Tsav skull“) bezeichnet.[5]
Weblinks
Einzelnachweise
- M. J. Benton, M. A. Shishkin, D. A. Unwin & E. N. Kurochkin (Hrsg.): The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, 2000, ISBN 0-521-55476-4, S. xxii–xxviii, (Digitalisat).
- D. A. Eberth: Stratigraphy and paleoenvironmental evolution of the dinosaur-rich Baruungoyot-Nemegt succession (Upper Cretaceous), Nemegt Basin, southern Mongolia. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 29–50, (Abstract).
- R. Gradziński, Z. Kielan-Jaworowska & T. Maryańska: Upper Cretaceous Djadokhta, Barun Goyot and Nemegt formations of Mongolia, including remarks on previous subdivisions. In: Acta Geologica Polonica, Band 27, Nummer 3, 1977, S. 281–318, (Digitalisat).
- D. A. Eberth, D. Badamgarav & P. J. Currie: The Baruungoyot-Nemegt Transition (Upper Cretaceous) at the Nemegt Type Area, Nemegt Basin, South Central Mongolia. In: Journal of the Paleontological Society of Korea, Band 25, Nummer 1, 2009, S. 1–15, (Digitalisat).
- Z. Kielan-Jaworowska, R. L. Cifelli & Z.-X. Luo: Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. Columbia University Press, New York / Chichester, 2004, ISBN 0-231-11918-6, S. 71, (Leseprobe).
- P. R. Bell: Cranial Osteology and Ontogeny of Saurolophus angustirostris from the Late Cretaceous of Mongolia with Comments on Saurolophus osborni from Canada. In: Acta Palaeontologica Polonica, Band 56, Nummer 4, 2011, S. 703–722, (Digitalisat).
- J. Van Itterbeeck, D. J. Horne, P. Bultynck & N. Vandenberghe: Stratigraphy and palaeoenvironment of the dinosaur-bearing Upper Cretaceous Iren Dabasu Formation, Inner Mongolia, People’s Republic of China. In: Cretaceous Research, Band 26, 2005, S. 699–725, (Digitalisat).
- K. Owocki, B. Kremer, M. Cotte & H. Bocherens: Diet preferences and climate inferred from oxygen and carbon isotopes of tooth enamel of Tarbosaurus bataar (Nemegt Formation, Upper Cretaceous, Mongolia). In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 537, 2020, Artikel 109190, doi:10.1016/j.palaeo.2019.05.012.
- J. F. Hicks, D. Fastovsky, D. J. Nichols & M. Watabe: Magnetostratigraphic correlation of Late Cretaceous dinosaur-bearing localities in the Nemegt and Ulan Nuur Basins, Gobi Desert, Mongolia. In: Geological Society of America Abstracts with Programs, Band 33, Nummer 6, 2001, S. 1920, (Abstract).
- T. Jerzykiewicz: Okavango Oasis, Kalahari Desert: A Contemporary Analogue for the Late Cretaceous Vertebrate Habitat of the Gobi Basin, Mongolia. In: Geoscience Canada, Band 25, Nummer 1, 1998, S. 15–26, (Abstract).
- P. J. Currie, D. Badamgarav & E. B. Koppelhus: The First Late Cretaceous Footprints from the Nemegt Locality in the Gobi of Mongolia. In: Ichnos, Band 10, Nummer 1, 2003, S. 1–13, doi:10.1080/10420940390235071.
- S. Ishigaki, M. Watabe, K. Tsogtbaatar & M. Saneyoshi: Dinosaur footprints from the Upper Cretaceous of Mongolia. In: Geological Quarterly, Band 53, Nummer 4, 2009, S. 449–460, (Digitalisat).
- Ph. J. Currie: Dinosaurs of the Gobi: Following in the footsteps of the Polish-Mongolian Expeditions. In: Palaeontologia Polonica, Band 67, 2016, S. 83–100, (Digitalisat).
- J. Nakajima, Y. Kobayashi, Ch. Tsogtbaatar, T. Tanaka, R. Takasaki, K. Tsogtbaatar, P. J. Currie & A. R. Fiorillo: Dinosaur tracks at the Nemegt locality: Paleobiological and paleoenvironmental implications. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 147–159, (Abstract).
- F. Fanti, P. Bell, P. Currie & K. Tsogtbaatar: The Nemegt Basin — One of the best field laboratories for interpreting Late Cretaceous terrestrial ecosystems. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 1–4, (Digitalisat).
- E. N. Kurochkin & R. Barsbold: The Russian-Mongolian expeditions and research in vertebrate palaeontology. In: M. J. Benton, M. A. Shishkin, D. M. Unwin & E. N. Kurochkin (Hrsg.): The Age of Dinosaurs in Russia and Mongolia, Cambridge University Press, Cambridge, 2000, ISBN 0-521-55476-4, S. 235–255, (Digitalisat).
- Z. Kielan-Jaworowska & N. Dovchin: Narrative of the Polish-Mongolian Palaeontological Expeditions 1963–1965. In: Palaeontologia Polonica, Band 19, 1969, S. 7–30, (Digitalisat).
- P. J. Currie, J. A. Wilson, F. Fanti, B. Mainbayar & K. Tsogtbaatar: Rediscovery of the type localities of the Late Cretaceous Mongolian sauropods Nemegtosaurus mongoliensis and Opisthocoelicaudia skarzynskii: Stratigraphic and taxonomic implications. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 5–13, (Digitalisat).
- H. Osmólska & E. Roniewicz: Deinocheiridae, a new family of theropod dinosaurs. In: Palaeontologia Polonica, Band 21, 1970, S. 5–19, (Digitalisat):
- Z. Kielan-Jaworowska & R. Barsbold: Narrative of the Polish-Mongolian Palaeontological Expeditions 1967–1971. In: Palaeontologia Polonica, Band 27, 1972, S. 5–13, (Digitalisat).
- R. Gradziński, J. Kaźmierczak & J. Lefeld: Geographical and Geological Data from the Polish-Mongolian Palaeontological Expeditions. In: Palaeontologia Polonica, Band 19, 1969, S. 33–82, (Digitalisat).
- R. Barsbold: Saurornithoididae, a new Family of small Theropod Dinosaurs from Central Asia and North America. In: Palaeontologia Polonica, Band 30, 1974, S. 5–22, (Digitalisat).
- S. L. Brusatte, T. D. Carr & M. A. Norell: The Osteology of Alioramus, A Gracile and Long-snouted Tyrannosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Mongolia. In: Bulletin of the American Museum of Natural History, Nummer 366, 2012, 197 S., (abrufbar).
- M. Watabe, K. Tsogtbaatar, S. Suzuki & M. Saneyoshi: Geology of dinosaur-fossil-bearing localities (Jurassic and Cretaceous: Mesozoic) in the Gobi Desert: Results of the HMNS-MPC Joint Paleontological Expedition. In: Hayashibara Museum of Natural Sciences Research Bulletin, Band 3, 2020, S. 41–118, (Digitalisat).
- D. B. Weishampel, D. E. Fastovsky, M. Watanabe, D. Varricchio, F. Jackson, K. Tsogtbaatar & R. Barsbold: New Oviraptorid Embryos from Bugin-Tsav, Nemegt Formation (Upper Cretaceous), Mongolia, with Insights into Their Habitat and Growth. In: Journal of Vertebrate Paleontology, Band 28, Nummer 4, 2008, S. 1110–1119, (Digitalisat).
- T. Tsuihiji, M. Watabe, K. Tsogtbaatar, T. Tsubamoto, R. Barsbold, S. Suzuki, A. H. Lee, R. C. Ridgely, Y. Kawahara & L. M. Witmer: Cranial Osteology of a Juvenile Specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. In: Journal of Vertebrate Paleontology, Band 31, Nummer 3, 2011, S. 497–517, (Digitalisat).
- Y.-N. Lee, R. Barsbold, P. J. Currie, Y. Kobayashi, H.-J. Lee, P. Godefroit, F. Escuillié & T. Chinzorig: Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus. In: Nature, Band 515, 2014, S. 257–260, (Digitalisat).
- G. F. Funston, P. J. Currie, D. A. Eberth, M. J. Ryan, T. Chinzorig, D. Badamgarav & N. R. Longrich: The first oviraptorosaur (Dinosauria: Theropoda) bonebed: evidence of gregarious behaviour in a maniraptoran theropod. In: Scientific Reports, Band 6, 2016, Artikel 35782, doi:10.1038/srep35782.
- P. J. Currie & E. B. Koppelhus: Extracting data from poached and old quarries in the Nemegt Formation of Mongolia. In: 6th Annual Fossil Preparation & Collections Symposium, Program and Abstracts, 2013, S. 23–28, (Digitalisat).
- L. D. DuBoff & M. D. Murray: Art Law: Cases and Materials. 2. Ausgabe, Wolters Kluwer Law & Business, 2017, ISBN 978-1-4548-7663-2, S. 776ff, (Leseprobe).
- National Geographic: Episode 3: The United States v. One Tyrannosaurus Bataar. In: Podcast. National Geographic, 30. Juni 2020, abgerufen am 11. Oktober 2020..
- U.S. Attorney’s Office Southern District of New York: Manhattan U.S. Attorney Announces Return To Mongolia Of Fossils Of Over 18 Dinosaur Skeletons. In: Press Releases. United States Department of Justice, 10. Juli 2014, abgerufen am 13. Oktober 2020.
- U.S. Attorney’s Office Southern District of New York: United States Seeks To Forfeit And Return A Tyrannosaurus Bataar Skull Looted From The Gobi Desert In Mongolia. In: Press Releases. United States Department of Justice, 16. Dezember 2015, abgerufen am 17. Oktober 2020.
- BBC News Entertainment & Arts: Nicolas Cage returns stolen dinosaur skull to Mongolia. In: BBC News. 22. Dezember 2015, abgerufen am 17. Oktober 2020.
- UNESCO: Cretaceous Dinosaur Fossil Sites in the Mongolian Gobi. In: Tentative Lists. UNESCO World Heritage Centre, 19. Dezember 2014, abgerufen am 16. Oktober 2020..
- F. Fanti, P. R. Bell, M. Tighe, L. A. Milan & E. Dinelli: Geochemical fingerprinting as a tool for repatriating poached dinosaur fossils in Mongolia: A case study for the Nemegt Locality, Gobi Desert. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 51–64, (Manuskriptversion).
- Y. Khand, D. Badamgarav, Y. Ariunchimeg & R. Barsbold: Cretaceous System in Mongolia and its depositional environments. In: H. Okada & N. J. Mateer (Hrsg.): Cretaceous Environments of Asia, Elsevier, Amsterdam, 2000, ISBN 0-444-50276-9, S. 49–80, (Leseprobe).
- V. A. Krassilov & N. M. Makulbekov: Isoetalean megasporophylls with megaspores from the Upper Cretaceous of Mongolia. In: Review of Palaeobotany and Palynology, Band 94, 1996, S. 231–238, (Digitalisat).
- J. Karczewska & M. Ziembińska-Tworzydło: New Upper Cretaceous Charophyta from the Nernegt Basin, Gobi Desert. In: Palaeontologia Polonica, Band 42, 1981, S. 97–146, (Digitalisat).
- V. A. Krassilov & N. M. Makulbekov: The First Finding of Gasteromycetes in the Cretaceous of Mongolia. In: Paleontological Journal, Band 37, Nummer 4, 2003, S. 439–442, (Digitalisat).
- K. Owocki, B. Kremer, B. Wrzosek, A. Królikowska & J. Kaźmierczak: Fungal Ferromanganese Mineralisation in Cretaceous Dinosaur Bones from the Gobi Desert, Mongolia. In: PLOS ONE, Band 11, Nummer 2, 2016, e0146293, doi:10.1371/journal.pone.0146293.
- J. Szczechura: Fresh-water Ostracodes from the Nemegt Formation (Upper Cretaceous) of Mongolia. In: Palaeontologia Polonica, Band 38, 1978, S. 65–121, (Digitalisat).
- P. Carbonel, J.-P. Colin, D. L. Danielopol, H. Löffler & I. Neustrueva: Paleoecology of Limnic Ostracodes: A Review of some Major Topics. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 62, 1988, S. 413–461, (Digitalisat).
- M. G. Newbrey, D. B. Brinkman, D. A. Winkler, E. A. Freedman, A. G. Neuman, D. W. Fowler & H. N. Woodward: Teleost centrum and jaw elements from the Upper Cretaceous Nemegt Formation (Campanian-Maastrichtian) of Mongolia and a re-identification of the fish centrum found with the theropod Raptorex kreigsteini. In: G. Arratia, H.-P. Schultze & M. V. H. Wilson (Hrsg.): Mesozoic Fishes 5 – Global Diversity and Evolution, Verlag Dr. Friedrich Pfeil, München, 2013, ISBN 978-3-89937-159-8, S. 291–303, (Digitalisat).
- Z. Roček: Mesozoic Anurans. In: H. Heatwole & R. L. Carroll (Hrsg.): Amphibian Biology Volume 4, Palaeontology: The Evolutionary History of Amphibians, Surrey Beatty & Sons, Chipping Norton, 2000, ISBN 0-949324-87-6, S. 1295–1331, (Digitalisat).
- L. I. Khosatzky & M. Młynarski: Chelonians from the Upper Cretaceous of the Gobi Desert, Mongolia. In: Palaeontologia Polonica, Band 25, 1971, S. 131–144, (Digitalisat).
- E. A. Cadena, D. T. Ksepka & M. A. Norell: New material of Mongolemys elegans Khosatzky and Mlynarski, 1971 (Testudines: Lindholmemydidae), from the Late Cretaceous of Mongolia with comments on bone histology and phylogeny. In: American Museum Novitates, Nummer 3766, 2013, S. 1–27, (Digitalisat).
- I. Danilov: GravemysSukhanov and Narmandakh, 1983 (Testudinoidea: Lindholmemydidae) from the Late Cretaceous of Asia: new data. In: PaleoBios, Band 23, Nummer 3, 2003, S. 9–19, (Digitalisat).
- G. L. Georgalis & W. G. Joyce: A Review of the Fossil Record of Old World Turtles of the Clade Pan-Trionychidae. In: Bulletin of the Peabody Museum of Natural History, Band 58, Nummer 1, 2017, S. 115–208, (Digitalisat).
- W. G. Joyce: A Review of the Fossil Record of Basal Mesozoic Turtles. In: Bulletin of the Peabody Museum of Natural History, Band 58, Nummer 1, 2017, S. 65–113, (Digitalisat).
- A. Pérez-García: Surviving the Cretaceous-Paleogene mass extinction event: A terrestrial stem turtle in the Cenozoic of Laurasia. In: Nature - Scientific Reports, Band 10, 2020, Artikel 1489, doi:10.1038/s41598-020-58511-8.
- V. R. Alifanov: The fossil record of Cretaceous lizards from Mongolia. In: M. J. Benton, M. A. Shishkin, D. M. Unwin & E. N. Kurochkin (Hrsg.): The Age of Dinosaurs in Russia and Mongolia, Cambridge University Press, Cambridge, 2000, ISBN 0-521-55476-4, S. 368–389, (Digitalisat).
- A. H. Turner: A Review of Shamosuchus and Paralligator (Crocodyliformes, Neosuchia) from the Cretaceous of Asia. In: PLOS ONE, Band 10, Nummer 2, 2015, e0118116, doi:10.1371/journal.pone.0118116.
- T. Tsuihiji, B. Andres, P. M. O'Connor, M. Watabe, K. Tsogtbaatar & B. Mainbayar: Gigantic pterosaurian remains from the Upper Cretaceous of Mongolia. In: Journal of Vertebrate Paleontology, Band 37, Nummer 5, 2017, Artikel e1361431, (Digitalisat).
- G. F. Funston, S. E. Mendonca, P. J. Currie & R. Barsbold: Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 201–220, (Digitalisat).
- S. Lee, Y.-N. Lee, A. Chinsamy, J. Lü, R. Barsbold & K. Tsogtbaatar: A new baby oviraptorid dinosaur (Dinosauria: Theropoda) from the Upper Cretaceous Nemegt Formation of Mongolia. In: PLOS ONE, Band 14, Nummer 2, 2019, e0210867 , doi:10.1371/journal.pone.0210867.
- G. F. Funston, Ch. Tsogtbaatar, K. Tsogtbaatar, Y. Kobayashi, C. Sullivan & P. J. Currie: A new two-fingered dinosaur sheds light on the radiation of Oviraptorosauria. In: Royal Society Open Science, Band 7, 2020, Artikel 201184, doi:10.1098/rsos.201184.
- Ch. Tsogtbaatar, Y. Kobayashi, K. Tsogtbaatar, P. J. Currie, R. Takasaki, T. Tanaka, M. Iijima & R. Barsbold: Ornithomimosaurs from the Nemegt Formation of Mongolia: manus morphological variation and diversity. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 494, 2018, S. 91–100, (Digitalisat).
- P. J. Currie: Theropods from the Cretaceous of Mongolia. In: M. J. Benton, M. A. Shishkin, D. M. Unwin & E. N. Kurochkin (Hrsg.): The Age of Dinosaurs in Russia and Mongolia, Cambridge University Press, Cambridge, 2000, ISBN 0-521-55476-4, S. 434–455, (Leseprobe).
- M. A. Norell, P. J. Makovicky, G. S. Bever, A. M. Balanoff, J. M. Clark, R. Barsbold & T. Rowe: A Review of the Mongolian Cretaceous Dinosaur Saurornithoides (Troodontidae: Theropoda). In: American Museum Novitates, Nummer 3654, 2009, 63 S., (Digitalisat).
- P. R. Bell, P. J. Currie & Y.-N. Lee: Tyrannosaur feeding traces on Deinocheirus (Theropoda:?Ornithomimosauria) remains from the Nemegt Formation (Late Cretaceous), Mongolia. In: Cretaceous Research, Band 37, 2012, S. 186–190, (Abstract).
- S. Lee, J.-Y. Park, Y.-N. Lee, S.-H. Kim, J. Lü, R. Barsbold & K. Tsogtbaatar: A new alvarezsaurid dinosaur from the nemegt formation of Mongolia. In: Nature - Scientific Reports, Band 9, 2019, Artikel 15493, doi:10.1038/s41598-019-52021-y.
- S. L. Brusatte & T. D. Carr: The phylogeny and evolutionary history of tyrannosauroid dinosaurs. In: Nature - Scientific Reports, Band 6, 2016, Artikel 20252, doi:10.1038/srep20252.
- A. O. Averianov & A. V. Lopatin: Sauropod diversity in the Upper Cretaceous Nemegt Formation of Mongolia — a possible new specimen of Nemegtosaurus. In: Acta Palaeontologica Polonica, Band 64, Nummer 2, 2019, S. 313–321, (Digitalisat).
- T. A. Tumanova: The Armored Dinosaurs of Mongolia. In: The Joint Soviet-Mongolian Paleontological Expedition Transaction, Band 32, 1987, S. 19, (Digitalisat der englischen Übersetzung).
- V. Arbour, P. J. Currie & D. Badamgarav: The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia. In: Zoological Journal of the Linnean Society, Band 172, 2014, S. 631–652, (Digitalisat).
- P. Penkalski & T. Tumanova: The cranial morphology and taxonomic status of Tarchia (Dinosauria: Ankylosauridae) from the Upper Cretaceous of Mongolia. In: Cretaceous Research, Band 70, 2017, S. 117–127, (Abstract).
- T. Maryańska & H. Osmólska: Pachycephalosauria, a new Suborder of Ornithischian Dinosaurs. In: Palaeontologia Polonica, Nummer 30, 1974, S. 45–102, (Digitalisat).
- H. Osmólska: Nemegt Formation. In: P. J. Currie & K. Padian (Hrsg.): Encyclopedia of Dinosaurs, Elsevier - Academic Press, San Diego / London / Boston / New York / Sydney / Tokyo / Toronto, 1997, ISBN 978-0-12-226810-6, S. 471f, (Leseprobe).
- E. N. Kurochkin, S. Chatterjee & K. E. Mikhailov: An Embryonic Enantiornithine Bird and Associated Eggs from the Cretaceous of Mongolia. In: Paleontological Journal, Band 47, Nummer 11, 2013, S. 1252–1269, (Digitalisat).
- J. A. Clarke & M. A. Norell: New Avialan Remains and a Review of the Known Avifauna from the Late Cretaceous Nemegt Formation of Mongolia. In: American Museum Novitates, Nummer 3447, 2004, S. 1–12, (Digitalisat).
- D. J. Varricchio & F. D. Jackson: Reproduction in Mesozoic birds and evolution of the modern avian reproductive mode. In: The Auk: Ornithological Advances, Band 133, 2016, S. 654–684, doi:10.1642/AUK-15-216.1.
- T. Tanaka, Y. Kabayashi, K. Kurihara, A. R. Fiorillo & M. Kano: The oldest Asian hesperornithiform from the Upper Cretaceous of Japan, and the phylogenetic reassessment of Hesperornithiformes. In: Journal of Systematic Palaeontology, Band 16, Nummer 8, 2017, S. 689–709, (Digitalisat).
- Z. Kielan-Jaworowska & J. H. Horum: Phylogeny and Systematics of Multituberculate Mammals. In: Palaeontology, Band 44, Teil 3, 2001, S. 389–429, (Digitalisat).
- Z. Kielan-Jaworowska & A. V. Sochava: The First Multituberculate from the Uppermost Cretaceous of the Gobi Desert (Mongolia). In: Acta Palaeontologica Polonica, Band 14, Nummer 3, 1969, S. 355–367, (Digitalisat).