Kognitive Neurowissenschaft
Die kognitive Neurowissenschaft (englisch cognitive neuroscience) beschäftigt sich mit den neuronalen Mechanismen, die mit kognitiven Fähigkeiten im Zusammenhang stehen.
Das Feld der kognitiven Neurowissenschaft weist Überschneidungen mit der Kognitionswissenschaft und der kognitiven Psychologie auf. Die Kognitionswissenschaft beschäftigt sich mit bewussten und unbewussten Vorgängen wie z. B. Wahrnehmung, Denken, Urteilen, Gedächtnis, Lernen und Sprache. Ihr Gegenstandsbereich ist allerdings nicht nur die Kognition, sondern auch Emotion, Motivation und Volition[1]. Während die kognitive Psychologie versucht den (menschlichen) Geist zu verstehen (z. B. die Entstehung von Erinnerungen, Gedanken etc.), beschäftigt sich die kognitive Neurowissenschaft mit den zugrundeliegenden neurobiologischen Prozessen. Also untersuchen die kognitive Psychologie und die kognitive Neurowissenschaft verschiedene Aspekte derselben Sache (z. B. Reaktionszeit, funktionelle Bildgebung). Sie beeinflussen sich gegenseitig, da ein genaueres Verständnis der mentalen Prozesse hilfreich für das Verständnis der zugrundeliegenden Hirnstrukturen ist und umgekehrt.
Die kognitive Neurowissenschaft ist eine recht junge Forschungsrichtung, deren Etablierung zu zahlreichen neuen Erkenntnissen und somit zu einem großen Sprung in der Untersuchung des menschlichen Gehirns führte.
Geschichte der kognitiven Neurowissenschaft
Historischer Hintergrund
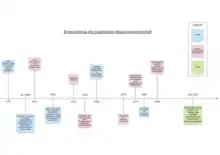
Gegenwärtig entstammen Forscher der kognitiven Neurowissenschaft gewöhnlich einem experimentalpsychologischen, kognitionspsychologischen, biopsychologischen, neurobiologischen, neurologischen, physikalischen oder mathematischen Hintergrund. Folglich sind die verwendeten Methoden vielfältig und umfassen psychophysikalische Experimente und funktionelle Bildgebung, aber auch Methoden der Neurophysiologie und auch der Neuroinformatik und der Computational Neuroscience. Die kognitive Neurowissenschaft in ihrem heutigen Verständnis hat eine lange Entstehungsgeschichte, welche durch verschiedene philosophische und wissenschaftliche Denkansätze geprägt wurde.
Wissenschaftlicher Hintergrund
Untersuchungen zu den Funktionen des Gehirns ließen sich ebenfalls bereits in der Antike finden, wie beispielsweise bei Galen (ca. 199–129 v. Chr.), der Gehirnverletzungen bei Gladiatoren untersuchte. Viele der antiken Vorstellungen blieben lange unangefochten. Die ersten Ansätze, die der heutigen Auffassung einer funktionellen Gliederung des menschlichen Gehirns entsprechen, entwickelten sich erst im frühen 19. Jahrhundert mit dem Aufkommen der Phrenologie nach Gall und Spurzheim. Viele andere Annahmen der Phrenologie sind zwar mittlerweile widerlegt, die Ansätze zur funktionalen Spezialisierung im Gehirn bleiben jedoch bis heute bestehen.[2]
In den folgenden Jahren wurden Untersuchungen des Gehirns essenzieller Bestandteil in der neurowissenschaftlichen Forschung, beispielsweise bei Forschern wie Broca und Wernicke. Dies führte zur Entdeckung der Funktionen verschiedener Gehirnareale. Besonderer Fokus lag hierbei auf der Untersuchung von Patienten mit Hirnschäden (Hirnläsionen), da hieraus Theorien zur normalen Funktionsweise des Gehirns abgeleitet werden konnten. Die Untersuchung von Hirnläsionen wird als kognitive Neuropsychologie bezeichnet und ist auch heute noch wichtiger Bestandteil der kognitiven Neurowissenschaft.[2]
Auch Experimente von Wilder Penfield und Kollegen lieferten weitreichende neue Erkenntnisse in der kognitiven Neurowissenschaft. Anfang des 20. Jahrhunderts führte das Forscherteam zahlreiche Operationen an menschlichen Gehirnen durch, während die Patienten bei Bewusstsein waren. Diese Eingriffe waren schmerzlos, weil im Gehirn keine Schmerzrezeptoren vorhanden sind. Dabei wurde herausgefunden, dass eine elektrische Stimulation spezifischer Gehirnregionen z. B. zu visuellen oder akustischen Erscheinungen oder motorischen Phänomenen führte. Daraus konnte geschlossen werden, dass bestimmte Gehirnregionen für bestimmte Funktionen in der menschlichen Wahrnehmung oder des Verhaltens zentral sind. Somit wurde erstmals eine experimentelle Bestätigung der Annahmen zur funktionalen Spezialisierung im Gehirn geliefert.[2] Neuere Forschungsergebnisse deuten jedoch darauf hin, dass verschiedene Regionen des Gehirns nur teilweise auf verschiedene Funktionen spezialisiert sind.
Ein weiterer wichtiger Schritt auf dem Weg zum heutigen Verständnis der kognitiven Neurowissenschaft war die Entwicklung der Annahme, dass die Funktionsweise des Gehirns der eines Computers ähnelt (komputationaler Ansatz). Eine frühe Theorie ist das Informationsverarbeitungsparadigma, welches ab den 1950er Jahren an Popularität gewann. Zum Beispiel argumentierte Broadbent, dass Informationen nach einem Stufenparadigma zuerst wahrgenommen werden, dann Aufmerksamkeit erhalten und im Kurzzeitgedächtnis gespeichert werden. Ein expliziteres komputationales Modell, das sich ab den 1980ern entwickelte, war das der neuronalen Netze. Dieses geht davon aus, dass die Informationsverarbeitung über die Interaktion miteinander verbundener neuronaler Knoten stattfindet.[2]
Die Geburt der modernen kognitiven Neurowissenschaft
Auch die Entwicklung neuer Bildgebungstechnologien ab den 1970er Jahren leistete einen bedeutenden Beitrag zur modernen kognitiven Neurowissenschaft. Besonders hervorzuheben ist hierbei die Erfindung des fMRT (funktionelle Magnetresonanztomographie). Das fMRT lieferte die Möglichkeit, die für die kognitiven Neurowissenschaften relevanten Teile des Gehirns zu beobachten, ohne das Gehirn zu schädigen. Schlussendlich führte also diese Kombination der Bildgebungstechnologien mit den Theorien der Kognitionspsychologie zu unserem heutigen Verständnis der kognitiven Neurowissenschaft.[2]
Die Disziplin der kognitiven Neurowissenschaft kann heute als Zusammenschluss der experimentellen Psychologie und der Neurowissenschaften betrachtet werden. Als Begründer dieser zusammengeschlossenen Disziplin können George A. Miller und Michael Gazzaniga gesehen werden. Als ein Vorreiter ist auch Alexander Romanowitsch Lurija anzusehen, der schon viel früher die Verschmelzung der vorgenannten Felder antizipierte und die Neurowissenschaft mit der Psychologie verband.
Philosophischer Hintergrund
Der Ausgangspunkt der kognitiven Neurowissenschaft lässt sich in der ursprünglich philosophischen Diskussion um das Leib-Seele-Problem finden, also der Frage, wie physische Masse (das Gehirn), psychische Erlebnisse erzeugen kann (z. B. Sinneswahrnehmungen). Für dieses Problem haben sich drei Lösungsansätze entwickelt: der Dualismus, die Dual-Aspect-Theory und der Reduktionismus. Die Grundannahme im Dualismus ist, dass Gehirn und Geist zwei vollständig unterschiedliche Phänomene sind. Gemäß der Dual-Aspect-Theory hingegen handelt es sich bei Gehirn und Geist um unterschiedliche Beschreibungen derselben Sache. Eine Analogie dazu aus der Physik ist das Elektron, welches als Welle und als Partikel beschrieben werden kann. Im Reduktionismus wird angenommen, dass geistige Konzepte wie Emotionen, Erinnerungen oder Aufmerksamkeit schlussendlich durch rein biologische Erklärungen (z. B. Neurotransmitter) ersetzt werden.[2]
Zukunft und weitere Forschungsbereiche der kognitiven Neurowissenschaft
Aktuelle Forschung widmet sich vor allem dem Verständnis des Gehirns als Netzwerk. Statt einem einzelnen Netzwerk könnten jedoch viele verschiedene Netzwerke bestehen, die je nach Bedarf aktiv sind oder nicht. Somit hätten nicht nur einzelne Hirnregionen spezielle Funktionen, sondern ganze Netzwerke im Gehirn übernehmen diese. Dieser Netzwerkansatz wird aktuell durch Versuche, das menschliche Konnektom abzubilden, veranschaulicht. Das Konnektom ist eine umfangreiche Karte aller Nervenverbindungen im Gehirn auf Makroebene (bis auf den Millimeter genau). Indem die Gehirne tausender Menschen mittels Magnetresonanztomographie untersucht werden, soll es beispielsweise möglich werden, Abweichungen im Konnektom, die mit Erkrankungen zusammenhängen, festzustellen.[1]
Als zentrale Herausforderung für die kognitive Neurowissenschaft kann das Erforschen der Beziehung zwischen Hirnstruktur und Funktionen (z. B. Verhalten) werden. Zusammengefasst gibt es dafür gegenwärtig drei Szenarien: Erstens könnte sich eine direkte eins-zu-eins Zuordnung von Hirnregionen zu Funktionen zeigen. Dies ist aus heutiger Sicht jedoch unwahrscheinlich. Zweitens könnten Hirnregionen zwar spezialisiert sein, jedoch müssten mehrere Regionen für eine Funktion miteinander interagieren. Drittens könnten Hirnregionen kaum spezialisiert sein und Funktionen entstünden durch die Interaktion mehrerer Netzwerke, welche wiederum zum Teil spezialisiert wären.[1]
Gegenwärtige Risiken im Kontext der Kognitiven Neurowissenschaft
Eine reduktionistische Perspektive auf die Kognitiven Neurowissenschaften (auch als Neuroessentialissmus bezeichnet) birgt Risiken für die gesamte Disziplin der Psychologie. Sie würde bedeuten, dass jegliches menschliches Erleben und Verhalten ausschließlich mithilfe von Gehirn-Prozessen zu erklären wäre. Traditionelle, tendenziell eher sozialwissenschaftlich orientierte psychologische Disziplinen, wie beispielsweise die Persönlichkeitspsychologie und die Sozialpsychologie, würden dann von der Kognitiven Neurowissenschaft verdrängt. Experten argumentieren jedoch, dass ein umfassendes Verständnis psychischer Prozesse allein auf Gehirnbasis nicht möglich sei, sondern multiple Aspekte und deren Interaktionen auf verschiedenen Ebenen berücksichtigt werden müssten, so zum Beispiel kognitive und emotionale Faktoren, sowie Kontexteinflüsse (vgl. Modell der Analyseebenen der Psychologie, Emergenz). Einige Risiken der wachsenden Rolle der KN in der Psychologie sollen daher im Folgenden beispielhaft angesprochen werden. Ein weiteres Problem der neurowissenschaftlichen Forschung in der Psychologie sind fehlerhafte logische Schlüsse, also solche, die über die auf Basis der gegenwärtigen Datenlage und der genutzten Methoden erlaubten Interpretationen hinausgehen. Dazu gehören neben dem bereits genannten Neuroessentialismus z. B. folgende Aspekte[3]:
- Ableitung von Kausalität aus Korrelationen: Korrelative Daten zeigen lediglich auf, dass ein Zusammenhang zwischen zwei Variablen besteht. Dabei kann nicht geschlussfolgert werden, welche der beiden Variablen die Ursache bzw. Wirkung ist.
- Neurorealismus: Neurorealismus fußt auf der Annahme, dass Neuroimaging-Daten verlässlich und objektiv sind, ohne dass dabei die Komplexität der Datenerhebung und -auswertung hinterfragt wird. Auf diese Weise werden neurowissenschaftliche Daten in der allgemeinen Öffentlichkeit gelegentlich unkritisch als Validierung oder Falsifizierung jeglicher Phänomene verwendet.
- Neuroredundanz: Neuroredundanz beschreibt die Tatsache, dass Neuroimaging-Daten zum Teil nicht in der Lage sind, einen Informationszugewinn gegenüber einfacher und weniger aufwändiger Erhebungsmethoden (z. B. Selbstberichtfragebögen) zu generieren.
- Reverse Inference Problem: Das Problem der Reverse Inference in der Kognitiven Neurowissenschaft beinhaltet die nicht zutreffende Schlussfolgerung, dass eine Gehirnaktivierung in der Bildgebung zwangsläufig einen darunterliegenden psychologischen Zustand oder kognitiven Prozess widerspiegeln muss.
Diese fehlerhaften logischen Schlüsse können dazu führen, dass neurowissenschaftliche Daten nicht ausreichend hinterfragt und überbewertet werden. Dies kann einen Effekt namens „Neuroseduction“ nach sich ziehen. Darunter versteht man das Phänomen, dass Personen eher dazu neigen, sich von fraglichen Schlussfolgerungen aus Studiendaten überzeugen zu lassen, wenn neurowissenschaftliche Daten und Erklärungen einbezogen werden.[3]
Risiken der Forschungsentwicklung
Die Reproduzierbarkeit von Studien wird in allen Bereichen der Psychologie diskutiert. Für die kognitiven Neurowissenschaften ist sie jedoch besonders relevant, da aus mehreren Studien und Analysen hervorgeht, dass besonders Neuroimaging-Studien einige methodische Schwächen aufweisen.[3] Hier werden einige Schwächen aufgezählt:
- Viele Studien der Kognitiven Neurowissenschaft sind sehr aufwändig und teuer, weswegen sie häufig nur eine geringe Anzahl an Probanden haben. Die statistische Power von Neuroimaging-Studien liegt einer Abschätzung zufolge folglich im Durchschnitt bei nur 8 % und ist somit deutlich geringer als die von Cohen angestrebten 80 %, was zur Folge hat, dass diese Neuroimaging-Studien weniger wahrscheinlich echte Effekte entdecken.
- The Winner’s Curse: In Studien mit zu geringer statistischer Power sind positive Ergebnisse wahrscheinlicher statistischer Zufall als in Studien mit adäquater statistischer Power.
- Die häufige Vielzahl an Analysen pro Studie einer Inflation von Fehlern 1. Art.
- Es wurde gezeigt, dass sich die Reliabilität von beispielsweise fMRT-Studien häufig weit unter den Ansprüchen anderer psychologischer Maße wie Fragebögen oder Interviews befindet.
Es wird vorgeschlagen, die Zusammenarbeit von Neuroimaging-Laboren, die die gleichen methodischen Protokolle benutzen, zu fördern. Damit können die statistische Power erhöht, der Winner’s Curse vermieden und das Risiko von falschen positiven Befunden minimiert werden.
In forschungsökonomischer Hinsicht gibt es weiterhin Risiken, die durch den Gewinn an der Bedeutung der Neurowissenschaft innerhalb der Psychologie entstehen können. Einerseits ist hier die bevorzugte Finanzierung von Forschungsprojekten mit neurowissenschaftlicher Fragestellung zu nennen. Die Prioritäten staatlicher Fördermittelgeber, wie z. B. des National Institute of Mental Health (NIMH), scheinen sich mehr in die Richtung von Projekten mit neurowissenschaftlichem Schwerpunkt zu verschieben.[3] Die Formulierung der strategischen Ziele des NIMH hat sich z. B. zunehmend an biologischen und bildgebenden Fragestellungen orientiert.[3] Dadurch könnten Psychologen bei ihrer Forschung unter Druck gesetzt werden, explizit in neurowissenschaftliche Fragestellungen nachzugehen oder neurowissenschaftliche Maße einzusetzen, selbst, wenn diese unökonomisch sein sollten.[3] Andererseits werden auch Auswirkungen auf die Personalpolitik von Universitäten diskutiert. So soll z. B. auf der Webseite der APA der Anteil der Stellenanzeigen in den Neurowissenschaften, als Bereich der Psychologie, innerhalb von drei Jahren von rund 40 % (2011) über 33 % (2012) auf 50 % (2013) angestiegen sein.[3]
Risiken im Zusammenspiel mit der klinisch-psychologischen Anwendung
Aufgrund neurowissenschaftlicher Erkenntnisse und damit verbundener Schlussfehler werden psychische Störungen konzeptuell gegenwärtig vermehrt als Erkrankungen des Gehirns verstanden.[4] Der Ansatz des konstitutiven Reduktionismus lässt einerseits zu, dass psychische Störungen auf der niedrigsten Analyseebene als Gehirnerkrankungen betrachtet werden, da jegliche psychologische Phänomene von neuronalen Faktoren mediiert werden. Jedoch gehen psychische Erkrankungen mit Dysfunktionen auf höheren Analyseebenen einher und werden auch durch diese definiert. Beispielsweise sind die ICD-10-Diagnosekriterien für F-Diagnosen bisher rein „psychologisch“ und beinhalten keine neuronalen, biologischen oder chemischen Aspekte.
Dass psychische Störungen auch mit Veränderungen in der Gehirnfunktion einhergehen, bedeutet nicht, dass diese Veränderungen kausal für die Entwicklung einer psychischen Störung sind. Die Verwendung des Begriffs „Gehirnerkrankung“ birgt daher die Gefahr, psychischen Störungen eine rein biologische Ätiologie zu unterstellen. Man geht in der klinisch-psychologischen und psychiatrischen Forschung jedoch davon aus, dass alle Störungsbilder Resultat einer multifaktoriellen Ätiopathogenese, bestehend aus einer Trias genetischer, biologischer und psychosozialer Faktoren, sind.
Anknüpfend an das veränderte Verständnis psychischer Störungen hat auch die Behandlung ebendieser einen Wandel erlebt. In der Psychotherapie wurden sogenannte „brain-based“ (übersetzt: gehirnbasierte) Psychotherapien entwickelt, welche auf Erkenntnissen der Kognitiven Neurowissenschaft aufbauen. Befürworter dieser Therapieformen argumentieren damit, dass neurowissenschaftliche Befunde maßgeblich zur Gestaltung effektiver psychotherapeutischer Interventionen beitragen. Fraglich ist dabei aber, ob diese Befunde tatsächlich eine Informationsbasis generieren, die über die Informationen aus sichtbarem Verhalten, Affekt und Kognition hinausgehen. Hierbei handelt es sich um ein praktisches Beispiel der Neuroredundanz. Darüber hinaus stellt die empirische Überprüfung des Nutzens von „brain-based“ Psychotherapien eine Schwierigkeit dar, die aus dem fehlenden Wissen über die Verknüpfung zwischen Gehirn und Verhalten hervorgeht (Erklärungslücke;[3]).
Neuroanatomische Grundlagen
Zentral für die Kognitive Neurowissenschaft sind physiologische Prozesse des Zentralen und des Peripheren Nervensystems sowie ihrer Kommunikationsprozesse mit dem Körper.
Dabei werden im Rahmen verschiedener Analyselevel sowohl Prozesse auf der neuronalen Ebene, als auch hinsichtlich der Interaktion von Hirnarealen bis hin zu kognitiven Phänomenen wie Erleben und Verhalten untersucht. Somit konnten auch wesentliche Erkenntnisse über die neuronale sowie die Gehirnphysiologie gewonnen werden.
Neuronale Ebene
Milliarden von Neuronen bilden das komplexe Nervensystem. Signale werden über elektrochemische Prozesse von Neuron zu Neuron übertragen. Wird ein Aktionspotential am Axonhügel, welcher zwischen dem Zellkörper und dem Axon eines Neurons liegt, ausgelöst, verschieben sich die Spannungsunterschiede am Axon, wodurch das Signal weitergetragen werden kann. Am Ende des Neurons befindet sich die Synapse, welche entweder auf chemischem Wege, über die Ausschüttung von unterschiedlichen Neurotransmittern, oder auf direktem (elektrischen) Weg das elektrische Potential (die Information), an das folgende Neuron oder einen Muskel überträgt. Die Information wird über die Frequenz der Feuerrate codiert. Neurone sind auf spezifische Frequenzen spezialisiert und solche mit ähnlicher funktioneller Spezialisierung liegen gruppiert beieinander. Aus diesem Grund sind auch bestimmte Hirnregionen für bestimmte Informationen spezialisiert. Neuronen bilden im Gehirn graue oder weiße Substanz, wobei graue Substanz aus neuronalen Zellkörpern und weiße Substanz aus Axonen und Gliazellen besteht. Die Großhirnrinde und der Subcortex bestehen aus Strukturen grauer Substanz, dazwischen befindet sich eine Masse aus weißer Substanz. Bahnen aus Nervenfaser verbinden verschiedene Gehirnregionen, sowohl innerhalb der Hirnhälften (Assoziationsbahnen), als auch zwischen den Hirnhälften (Kommissuren mit dem Corpus Callosum als zentralem System) oder zwischen corticalen und subcorticalen Strukturen.[5]
Funktionelle Ebene
Die Basalganglien sind bilateral in der Tiefe des Cortex angelegt. Sie sind funktional relevant für die Regulation von Bewegungen sowie die Intensität motorischen Verhaltens. Das Belohnungslernen und die Bildung von Gewohnheiten gilt als ebenso in den Basalganglien zu verorten. Läsionen in diesem Bereich führen zu Hypokinetik oder Hyperkinetik. Bekannte Erkrankungen, die in Verbindung mit den Basalganglien stehen, sind z. B. Chorea Huntington oder Morbus Parkinson.
Das limbische System setzt den Organismus auf Grundlage der aktuellen Bedürfnisse, der gegenwärtigen Situation und früherer Erfahrungen mit der Umwelt in Beziehung. Verschiedene Strukturen tun dies auf unterschiedliche Art und Weise. Die Amygdala gilt als zentral für die Detektion von Angst und damit auch für die Furchtkonditionierung. Der Gyrus cinguli wird mit der Entstehung und Verarbeitung emotionaler und kognitiver Konflikte assoziiert. Der Hippocampus wird als von großer Bedeutung für das Lernen und die Gedächtniskonsolidierung gesehen, ebenso wie der Mammilarkörper. Im Riechkolben werden olfaktorische Stimuli detektiert. Diese Reize können die Stimmung und Erinnerung des Menschen beeinflussen. Läsionen im limbischen System äußern sich je nach betroffener Region mannigfaltig.
Das Zwischenhirn (Dienzephalon) lässt sich grob in die Strukturen Thalamus und Hypothalamus einteilen. Der Thalamus verschaltet alle Sinneseindrücke (außer olfaktorische Reize) und projiziert in fast alle Regionen des Cortex. Der Hypothalamus spielt eine zentrale Rolle zur Regulation von Vitalfunktionen wie Körpertemperatur, Hunger, Durst, sexueller Aktivität und endokriner Funktionen, z. B. Wachstum.
Das Mittelhirn wird mit Orientierung, auditiver Verarbeitung und motorischem Verhalten in Verbindung gebracht. Das Mittelhirndach (Tectum) enthält zwei besonders bedeutsame Strukturen für die Kognitive Neurowissenschaft: Die Colliculi superiores scheinen sensorische Informationen zu integrieren und ermöglichen so eine schnelle Orientierung des Organismus hin zu auffälligen Stimuli. Die Colliculi inferiores hingegen gelten als zentral für die auditive Verarbeitung. Die Substantia nigra steht in enger Verbindung mit den Basalganglien und wird folglich als entscheidende Struktur für Bewegungsmuster verstanden.
Das Hinterhirn (Metenzephalon) besteht aus dem Kleinhirn (Cerebellum) und der Brücke (Pons). Das Kleinhirn gilt als zentral für die Integration motorischer Befehle und sensorischen Feedbacks. Die Brücke erhält visuelle Informationen, um Augen und Körper zu bewegen. Das Markhirn (Medulla oblongata) reguliert vitale Funktionen, wie die Atmung oder den Schlafzyklus.[5]
Kernmethoden der kognitiven Neurowissenschaft
Elektrophysiologische Verfahren
Im Rahmen der Kognitiven Neurowissenschaft werden häufig elektrophysiologische Verfahren, bei denen mithilfe der Messung elektrischer Potenziale die biochemischen und biophysikalischen Vorgänge des Organismus untersucht werden, genutzt. Im Rahmen der kognitiven Neurowissenschaften bieten diese Verfahren die Möglichkeit, neuronale Aktivität in direkter Weise zu messen. Zu den Verfahren gehören die Einzelzellableitung sowie die Elektroenzephalographie (EEG).
Einzelzellableitungen
Bei Einzelzellableitungen wird eine Elektrode entweder direkt in eine Zelle eingeführt (intrazelluläre Messung) oder außerhalb der Zellmembran platziert (extrazelluläre Messung). So kann gemessen werden, wie viele Aktionspotenziale ein Neuron als Reaktion auf einen bestimmten Stimulus auslöst. Wenn die Aktivität mehrerer nahe beieinander liegender Neuronen durch eine Elektrode aufgezeichnet wird, spricht man von Mehrzellableitungen. Durch spezielle Algorithmen, die das kombinierte Signal zergliedern, erhält man auch hier Rückschlüsse auf die Beiträge einzelner Neuronen. Die Methode ist hochinvasiv und wird daher fast ausschließlich bei Versuchstieren angewandt.
Zu der Frage, wie viele Neuronen für die Repräsentation einer einzelnen Information im Gehirn zuständig sind, wird in der Kognitiven Neurowissenschaft der Ansatz der „sparsamen verteilten Repräsentation“ (engl. „sparse distributed representation“) aktuell hauptsächlich vertreten. Dieser Ansatz besagt zum einen, dass die Informationen über einen Stimulus auf mehrere Neuronen verteilt sind, statt von einem einzigen Neuron kodiert zu werden (Großmutterneuron). Zum anderen wird davon ausgegangen, dass nur ein Teil der Neuronen einer Neuronengruppe Informationen über einen Stimulus führt, um Energie zu sparen.[2]
Elektroenzephalographie (EEG)
Bei der Elektroenzephalographie werden die elektrischen Signale des Gehirns noninvasiv mittels mehrerer Elektroden gemessen, die auf der Schädeloberfläche angebracht werden.
Während eine Referenzelektrode (z. B. an der Nase oder am Schläfenbein) benötigt wird, können die Schädelelektroden selbst an vielen verschiedenen Stellen angebracht werden. Um ihre Position zu beschreiben, wird häufig das 10-20 System von Jasper (1958) verwendet.
Es ist wichtig zu beachten, dass sich ein EEG kaum zur räumlichen Ortung eines elektrischen Signals eignet, da der Ursprung des Signals nicht unbedingt in unmittelbarer Nähe zur Elektrode liegen muss. Ein EEG besitzt hingegen eine sehr gute zeitliche Auflösung und ist also sehr gut zur Messung der zeitlichen Zusammenhänge zwischen kognitiven Ereignissen und neuraler Aktivität geeignet.
Als EEG-Oszillationen bezeichnet man wellenförmige Schwingungen im EEG-Signal. Verschiedene Schwingungsmuster werden als charakteristisch für die verschieden Schlaf- und Wachphasen angesehen. Außerdem werden sie mit verschiedenen kognitiven Prozessen im Wachzustand in Verbindung gebracht, z. B. scheinen Alpha-Wellen in Verbindung mit erhöhter Aufmerksamkeit zu stehen. Die häufigste Nutzung von EEG ist die Methode der ereigniskorrelierten Potentiale (EKP, englisch event-related potentials (ERP)). Die EEG-Wellenform reflektiert neuronale Aktivität aus allen Bereichen des Gehirns. Einige davon sind spezifisch für die laufende Aufgabe (Lesen, Rechnen…), aber die meisten entstehen durch spontane Aktivierung oder andere Neurone, die nicht direkt etwas mit der Aufgabe zu tun haben. Das eigentliche Signal dagegen ist sehr schwach, sodass man nur schwer Rückschlüsse daraus ziehen kann. Jedoch ist es möglich, durch Mittelung über mehrere Durchgänge das Erkennen von Signalen zu erhöhen.
Die graphische Darstellung erfolgt typischerweise in einem Liniendiagramm, mit „Zeit“ auf der x-Achse und „Elektrodenpotential“ auf der y-Achse. Es besteht aus einer Reihe positiver und negativer Peaks („Ausschläge“ = Extremwerte), welche nach ihrer Valenz gefolgt von der ungefähren Zeit des Peaks in Millisekunden (z. B. P300, N400) benannt werden.[2]
Mentale Chronometrie
Die Mentale Chronometrie ist die Untersuchung des zeitlichen Verlaufs der Informationsverarbeitung im menschlichen Nervensystem. Als Grundidee sollen sich Änderungen in der Art oder Effizienz der Informationsverarbeitung in der Zeit bemerkbar machen, die für die Erledigung einer Aufgabe benötigt wird. Eine mathematische Aufgabe umfasst beispielsweise in der Regel eine Reihe von Schritten, einschließlich der visuellen Erkennung der Ziffern, der Durchführung von Berechnungen und des Generierens einer Antwort. Diese können im abgebildeten Signal betrachtet werden.
Frühe Peaks könnten so das Erkennen und Enkodieren bedeuten, spätere z. B. Vergleiche mit den relevanten Informationen. Man kann beobachten, wie sich die Amplitude der Peaks verändert, wenn man die Aufgabenstellung variiert.[2]
Gesichtsverarbeitung
Eine ERP-Komponente, die relativ selektiv für die Verarbeitung jeglicher Gesichter ist, ist N170. N250 dagegen ist selektiv für berühmte oder vertraut Gesichter und reagiert auf Präsentationen von verschiedenen Bildern der gleichen Person. Damit kodiert sie besondere Merkmale eines spezifischen Gesichts.[2]
Endogene und exogene ERP-Komponenten
ERP-Komponenten werden klassischerweise als exogen oder endogen bezeichnet. Exogene Komponenten hängen von den physikalischen Eigenschaften eines Stimulus (Größe, Intensität, …) ab und sind evozierte Potentiale. Endogene Komponenten hängen dagegen von den Eigenschaften der Aufgabe ab. Exogene Komponenten zeigen sich meist zeitiger im EEG als endogene Komponenten.[2]
Weiterentwicklung: Magnetenzephalographie (MEG)
Das Aufnehmen magnetischer statt elektrischer Signale ist die Kerneigenschaft des MEGs. Die verwendete Vorrichtung benötigt extreme Kühlung (durch flüssiges Helium) und Isolation des Systems in einem magnetisch abgeschirmten Raum. Dementsprechend sind die Kosten für MEG sehr viel höher als die für EEG. Die sehr viel bessere räumliche Auflösung ist dagegen ein großer Vorteil des MEG.[2]
Bildgebung
Bei der Bildgebung (auch Bildgebende Verfahren oder Bildgebende Diagnostik) unterscheidet man in den kognitiven Neurowissenschaften zwischen strukturellen und funktionellen Methoden.
Arten der Bildgebung
Strukturelle Bildgebung basiert grundlegend auf der Erkenntnis, dass verschiedene Gewebe unterschiedliche physikalische Eigenschaften aufweisen, die für eine statische Abbildung der Gewebestrukturen genutzt werden können. Die gängigsten strukturellen Methoden sind die Computertomographie (CT) und die Magnetresonanztomographie (MRT).
Unter funktioneller Bildgebung versteht man im Gegensatz dazu die Messung temporärer Veränderungen im Gehirn, die mit kognitiven Prozessen assoziiert. Die häufigsten funktionellen Methoden sind die Positronen-Emissions-Tomographie (PET) und die funktionelle Magnetresonanztomographie (fMRT).
Experimentelle Designs von Bildgebungsstudien
Zum Vergleich der Hirnaktivitäten bei verschiedenen Bedingungen eines Experiments, um beispielsweise bestimmte funktionelle Spezialisierungen einzelner Areale zu erkennen, eignen sich, je nach Datenlage, unterschiedliche Methoden.[2]
Eine dieser Methoden ist die “Cognitive Subtraction”. Hierbei wird die Hirnaktivität bei einer Aufgabe, welche die zu untersuchende kognitive Komponente erfordert, mit der Hirnaktivität einer Kontrollbedingung verglichen, welche diese kognitive Komponente nicht enthält. Problematisch ist hierbei jedoch die Annahme der “Pure Insertion”, also die Annahme, dass das Hinzufügen einer Komponente die anderen in der Aufgabe enthaltenen Komponenten nicht beeinflusst. In der Forschung zeigte sich aber, dass die Aufgabe durch die resultierenden Wechselwirkungen verändert und die Bildgebungsdaten mehrdeutig werden. Um das Problem der Interaktionen zu reduzieren, können statt der “Cognitive Subtraction” mittels “Cognitive Conjunction” mehr als zwei Bedingungen, denen eine kognitive Komponente gemein ist, miteinander verglichen werden.[2]
Mittels “funktioneller Integration” kann herausgefunden werden, wie verschiedene Hirnregionen miteinander kommunizieren. Dabei wird untersucht, inwiefern die Aktivität in verschiedenen Hirnregionen voneinander abhängig ist und die Korrelation dieser Aktivitäten betrachtet.[2]
In Bezug auf die Gruppierung der Stimuli wird weiterhin zwischen Blockdesign und Event-Related Design unterschieden. Im Blockdesign werden die Stimuli gruppiert, die zur gleichen Experimentalbedingung gehören. Bei manchen Experimenten ist eine Gruppierung der Stimuli vor der Durchführung jedoch noch nicht möglich, weil sie sich für jeden Probanden subjektiv unterscheiden und individuell klassifiziert werden. So auch bei der Untersuchung des Zungenspitzenphänomens. In solchen Fällen empfiehlt sich ein Event-Related Design.[2]
Analyse von Bildgebungsdaten
Aufgrund potenzieller individueller Unterschiede und Aufnahmemängel (z. B. Belichtung, technische Schwierigkeiten) ist die fehlerfreie statistische Analyse und Interpretation funktioneller Bildgebungsdaten eine größere Herausforderung. Eine mehrstufige Aufbereitung der gewonnenen Daten ist notwendig, um eine Mittelung der personenbezogenen Daten und eine statistische Analyse vornehmen zu können.
Dazu können folgende Schritte zählen:
- die Korrektur für Kopfbewegungen während der Messung
- die Koregistrierung
- die stereotaktische Normalisierung
- das Glätten (Smoothing)[2]
Dabei umfasst die Koregistrierung den Abgleich funktioneller Aufnahmen mit höher aufgelösten strukturellen Aufnahmen. Bei der stereotaktischen (räumlichen) Normalisierung wird das Gehirn in Voxel (Datenpunkten in einem dreidimensionalen Gitter) unterteilt. Die ermittelten XYZ-Koordinaten werden den entsprechenden Voxel-Koordinaten eines Standardreferenzgehirns zugeordnet, den sogenannten Talairach-Koordinaten, um den standardisierten Vergleich der Aktivierungsmuster mehrerer Personen innerhalb eines einheitlichen dreidimensionalen Referenzraum zu ermöglichen. Beim anschließenden Glätten wird ein Teil des ursprünglichen Aktivierungsniveaus eines bestimmten Voxels auf benachbarte Voxel verteilt, um das Signal-Rausch-Verhältnis zu verbessern. Im letzten Schritt erfolgt die statistische Datenanalyse getrennt für jedes Voxel und die damit verbundene Signifikanzkorrektur für multiple Vergleiche (siehe auch Alphafehler-Kumulierung und Falscherkennungsrate).[2]
Interpretation von Bildgebungsdaten
Ein Areal wird bei funktioneller Bildgebung als aktiviert interpretiert, wenn es sich statistisch signifikant von der Kontrollbedingung unterscheidet. Ist ein Areal während einer bestimmten Aufgabe aktiv, lässt sich aber nicht kausal auf die kognitive Notwendigkeit dieses Areals für diese Aufgabe rückschließen (Reverse Inference Problem). Alternative Erklärungen sind beispielsweise interindividuelle kognitive Strategien, eine allgemeinere, übergeordnete Rolle des Areals (z. B. Aufmerksamkeit), inhibitorische neuronale Aktivität oder Zufall. Dementsprechend können Läsionsstudien nicht vollständig von Bildgebungsstudien ersetzt werden. Ob mittels Bildgebung zwischen exzitatorischen und inhibitorischen Prozessen unterschieden werden kann, ist bisher unklar.[2] So liegt auch die Möglichkeit, mittels Bildgebung „Gedanken lesen zu können“, noch in weiter Ferne.[6]
Läsionsstudien
Eine weitere Kernmethode der kognitiven Neurowissenschaft ist das Betrachten von Patienten mit Gehirnläsionen (z. B. in Folge von neurochirurgischen Eingriffen, Schlaganfällen, Traumata, Tumoren, viralen Infektionen oder neurodegenerativen Erkrankungen einerseits oder künstlich erzeugten und reversiblen Läsionen andererseits). Dabei wird untersucht, welche Funktionen im Gehirn erhalten bleiben und welche ausfallen bzw. eingeschränkt werden, wenn ein bestimmter Bereich des Gehirns gestört ist. Anschließend soll daraus auf die Funktion des Bereiches geschlussfolgert werden. Untersuchungen dieser Art gehören zum Fachgebiet der Neuropsychologie, die auch klinische Teilbereiche umfasst.[2]
Die Neuropsychologie hat sich in zwei verschiedene Richtungen entwickelt:
Die klassische Neuropsychologie versucht die Funktion einer Gehirnregion abzuleiten. Hierfür wird das Muster eingeschränkter und funktionstüchtiger Fähigkeiten von Patienten mit einer Läsion in dieser Region untersucht. Das ermittelte Fähigkeitsmuster ist dabei mit spezifischen Gehirnregionen assoziiert. Die bevorzugte Methode sind Gruppenstudien.[2]
Die kognitive Neuropsychologie versucht dagegen, „Bausteine der Kognition“ aus dem Muster eingeschränkter und funktionstüchtiger Fähigkeiten abzuleiten, die unabhängig von bestimmten Gehirnregionen betrachtet werden können. Die bevorzugte Methode sind dabei Einzelfallstudien.[2]
Doppelte Dissoziation
Aus der Beobachtung, dass Patienten mit einer Läsion in Bereich X eine Aufgabe A, aber nicht Aufgabe B lösen können und Patienten mit einer Läsion in Bereich Y Aufgabe B, aber nicht Aufgabe A lösen können, kann man ableiten, dass sich diese beiden Gehirnbereiche (X und Y) in ihrer Funktionalität unterscheiden. Diese Idee der doppelten Dissoziation wird sich in Läsionsstudien zunutze gemacht, um die Funktionalität unterschiedlicher Gehirnbereiche abgrenzen zu können.[2]
Einzelfallstudien
Einzelfallstudien ermöglichen die spezifische Untersuchung hirngeschädigter Patienten. Sie spielen eine wichtige Rolle, um Komponenten kognitiver Systeme festzustellen. Einzelfallstudien sind als gültige Forschungsmethode anerkannt und liefern Daten, mit denen Theorien getestet, verändert und weiterentwickelt werden können. Neue Theorien können jedoch auf Grundlage von Beobachtungen eines Einzelfalls nicht abgeleitet werden, da Verallgemeinerungen nur eingeschränkt möglich sind. Es gilt deshalb abzuschätzen, in welchem Umfang Ergebnisse generalisierbar sind.[2]
Gruppenstudien
Gruppenstudien können verschiedene Arten von Fragen behandeln, die sich vom Einzelfallansatz unterscheiden. Dazu werden hauptsächlich Mittelwerte berechnet, um Verallgemeinerungen auf allgemeine kognitive Mechanismen zuzulassen. Da Läsionen in der Regel groß und selten auf die interessierende Region beschränkt sind, bietet die Untersuchung mehrerer Patienten den Vorteil, lokalisieren zu können, welche Region für eine bestimmte Aufgabe wesentlich ist.[2]
Experimentelle Läsionen an Tieren
Die Untersuchung von Tieren anhand experimenteller Läsionen fällt unter den Begriff der Behavioral Neuroscience (Verhaltensneurowissenschaft). Indem chirurgisch künstliche Läsionen verursacht werden, ergibt sich die Möglichkeit, den Zustand vor und nach der operativen Läsion zu vergleichen.[2]
Methoden, die zur Verursachung von Läsionen verwendet werden, sind:
- Aspiration, das „Absaugen“ einer bestimmten Region
- Transsektion, das Durchtrennen von Bündeln weißer Substanz
- Neurochemische Läsion anhand von Toxinen
- Reversible, temporäre Läsionen, z. B. durch pharmakologische Manipulation oder Kühlung
Es ist zu beachten, dass sich bei der Untersuchung an Tiermodellen einige Schwierigkeiten ergeben, z. B. eine artgerechte Tierhaltung. Experimentelle Läsionsstudien sind aus ethischen Gründen am Menschen nicht erlaubt. Stattdessen verwendet man magnetische oder elektrische Methoden, die die Gehirnfunktionen nur kurzzeitig beeinflussen.[2]
Magnetische Methoden (TMS)
Die Transkranielle Magnetstimulation (TMS) stellt eine weitere Läsionsmethode in der kognitiven Neurowissenschaft dar. Hierbei wird das Gehirn mit Hilfe einer Magnetspule stimuliert, wodurch kognitive Funktionen temporär unterbrochen werden können. Diese Unterbrechung wird als reversible oder „virtuelle“ Läsion bezeichnet.[2]
Praktische Aspekte
Es muss für jede Fragestellung der richtige Zeitpunkt und Ort für die TMS-Impulse gefunden werden. Der Zeitpunkt kann aus theoretischen Überlegungen abgeleitet oder experimentell variiert werden. Alternativ kann eine ganze Reihe von Impulsen eingesetzt werden, dann spricht man von repetitiver transkranieller Magnetstimulation (rTMS). Studien zur Wahrnehmung verwenden beispielsweise eher einzelne Impulse, Studien zu höheren kognitiven Funktionen (wie z. B. Gedächtnis und Sprache) eher rTMS.[2]
Um kritische Regionen zu identifizieren, werden Orientierungspunkte am Schädel (z. B. am Inion, einer Erhebung am Hinterkopf) genutzt. Es kann ein präziser Punkt oder ein Gitter von z. B. 6 Punkten in einem 2 x 3 cm großen Bereich überprüft werden. Entsprechend werden jeweils die angrenzenden Gehirnregionen als Kontrollregionen verwendet, um die Wirkung der Stimulation zu überprüfen. Wenn Anlass zur Vermutung besteht, dass die kognitive Funktion lateralisiert ist (also nur von einer Gehirnhälfte übernommen wird), kann als Kontrollwert die gleiche Stelle in der anderen Hemisphäre ebenfalls stimuliert werden.[2]
Es ist sinnvoll, nicht-kritische Regionen oder Zeitfenster als Kontrollbedingungen zu nutzen, da bei der TMS auch periphere Effekte auftreten (z. B. lautes Pulsgeräusch, Zucken von Gesichtsnerven und -muskeln) und ohne geeignete Kontrolle die Messergebnisse verzerren können.[2]
Elektrische Methoden (TES)
Die Stimulation des Gehirn mit elektrischem Strom bezeichnet man als transkranielle Elektrostimulation (TES). Es existieren unterschiedliche Methoden, von denen einige invasiver sind als andere.
tDCS
Die transkranielle Gleichstromstimulation (engl. transcranial direct current stimulation, tDCS) nutzt nur einen sehr schwachen Stromimpuls im Vergleich beispielsweise zur Elektrokonvulsionstherapie (engl. electroconvulsive therapy, ECT). Bei der tDCS wird ein stimulierendes Elektroden-Pad über der interessierenden Region, sowie ein Kontroll-Pad über einer entsprechenden nichtinteressierenden Region platziert. Nach der Stimulation führt der zu Untersuchende einen kognitive Aufgabe durch. Die Ergebnisse werden im Anschluss mit denen einer Scheinstimulation verglichen.[2]
Während im ECT der elektrische Impuls von der Anode (positiv geladen) zur Kathode (negativ geladen), also kathodisch, geleitet wird, findet die Stimulation im tDCS entweder anodisch oder kathodisch statt. Kathodische tDCS verringert die neuronale Erregung der stimulierten Hirnregion. Das heißt, die spontane Feuerrate der Neuronen wird reduziert. Dabei wird das exzitatorische Glutamatsystem beeinflusst und die Leistung in einer Aufgabe verschlechtert sich tendenziell. Die anodische tDCS hingegen verbessert die neuronale Erregbarkeit eher, da das inhibitorische GABA-System beeinflusst wird, sodass die spontane Feuergeschwindigkeit der Neuronen steigt und sich die Leistung tendenziell verbessert.[2]
tACS
Die transkranielle Wechselstromstimulation (engl. transcranial alternating current stimulation, tACS) ist eine Methode zur Stimulation von Neuronen[1]. Der Aufbau gleicht dem des tDCS[1], es wird jedoch kein elektrischer Gleichstrom angelegt, sondern ein sinusförmiger Strom, der mit der natürlichen rhythmischen elektrischen Aktivität des Gehirns interagiert[1]. Die Eigenschwingung des Gehirns wird damit verstärkt. Die Methode des tACS wird in Studien sowohl zur Behandlung von Gehirnerkrankungen als auch zur Verbesserung der Gehirnfunktion bei gesunden Menschen getestet.
tRNS
Die transkranielle Rauschstimulation (engl. transcranial random noise stimulation, tRNS) ist eine Modifikation des tACS und die bisher am wenigsten erforschte Methode[1]. Beim tRNS wird im Vergleich zum tACS Wechselstrom mit zufälligen (anstatt festen) Interstimulus-Intervallen (Zeitabstände zwischen den Stimulationen) und Amplituden verwendet. Ein möglicher diskutierter Nutzen der tRNS-Methode liegt in der Verbesserung von kognitiven Funktionen[1]. Der Grundgedanke dahinter ist, dass bestimmte Oszillatorische Prozesse im Gehirn mit gesunden kognitiven und motorischen Funktionen in Verbindung gebracht werden. Liegt eine gestörte oszillatorische Aktivität vor, beeinträchtigt diese möglicherweise kognitive und motorische Funktionen. Durch das tRNS soll diese Aktivität wiederhergestellt werden.
Zentrale Untersuchungsbereiche der Kognitiven Neurowissenschaft
Im Folgenden werden einige zentrale Forschungsbereiche der Kognitiven Neurowissenschaft vorgestellt.
Gehirnentwicklung
Die kognitiven Neurowissenschaften beschäftigen sich zentral mit den Prozessen der Gehirnentwicklung, bei denen zwischen struktureller und funktioneller Gehirnentwicklung unterschieden wird. Die Gehirnentwicklung ist ein Prozess, der während der Schwangerschaft beginnt und ein Leben lang andauert.[1]
Strukturelle Gehirnentwicklung
Unter struktureller Gehirnentwicklung versteht man die Bildung und Reifung des Gehirns. Dabei ist die Nature-Nurture-Debatte eine grundlegende Debatte der kognitiven Neurowissenschaften. Sie behandelt die Frage, in welchem Ausmaß Kognition und Verhalten von genetischen Einflüssen oder Umwelteinflüssen bestimmt werden.[1]
Gottlieb (1992)[7] unterscheidet die Idee der prädeterminierten Entwicklung und der probabilistischen Entwicklung. Der Ansatz der prädeterminierten Entwicklung geht davon aus, dass die Gehirnstruktur allein durch die Gene bestimmt wird. Bei der Geburt wären damit alle Weichen der Entwicklung schon gestellt. Unser Erleben und die Erfahrungen, die wir machen, würden demnach von der Struktur unseres Gehirns beeinflusst. In der wissenschaftlichen Gemeinschaft ist der probabilistische Ansatz mittlerweile weiter verbreitet. Dieser geht neben genetischen Einflüssen von einer starken Beeinflussung der Gehirnstruktur durch die Umwelt aus. Demnach verändern unterschiedliche Umwelteinflüsse die Genexpression und umgekehrt. Über die gesamte Lebensspanne hinweg nehmen sämtliche alltägliche Erfahrungen Einfluss auf die Gehirnentwicklung. Das bedeutet, dass identische Erbanlagen, wie sie bei monozygoten (eineiigen) Zwillingen vorliegen, auch in ähnlicher Umwelt nicht zur gleichen Gehirnentwicklung führen, da einzelne Erfahrungen sich unterscheiden und die Genexpression beeinflussen[1].
Diese Veränderbarkeit des Gehirns nennt man neuronale Plastizität. Es werden pränatale und postnatale Phasen der Gehirnentwicklung unterschieden.[1]
Pränatale Gehirnentwicklung
Pränatale Gehirnentwicklung bezieht sich auf die Gehirnentwicklung während der Schwangerschaft. Das Nervensystem entwickelt sich aus dem Neuralrohr, das aus der Neuralplatte hervorgeht und in Ausbuchtungen und Windungen organisiert ist. Aus diesen ergeben sich verschiedene Teile des Gehirns. An der Mulde des Neuralrohrs befinden sich Gewebebereiche mit erhöhter Zellproliferation, sogenannte Proliferationszonen. Dort werden durch schnelle Zellteilung der Vorläuferzellen Neurone und Gliazellen gebildet. Diese bewegen sich durch Migration an die Stellen, an denen sie am reifen Gehirn benötigt werden. Radiale Gliazellen dienen dabei als Gerüst, an denen Neuronen entlang wandern können. Außerdem werden ältere Zellen passiv durch die neueren Zellen verdrängt und an die Hirnoberfläche gedrückt.[1]
Postnatale Gehirnentwicklung
Die meisten Neurone werden vor der Geburt gebildet. Nach der Geburt findet Gehirnwachstum durch Bildung von Synapsen (Synaptogenese), Dendriten und Axonbündeln sowie Myelinisierung statt. Feinabstimmung und Vernetzung bestehender Strukturen führt zu einer effizienteren Funktionsweise. Überflüssige Synapsen werden eliminiert.[1]
Protomap- und Protocortex – Theorien der Gehirnentwicklung
Es existieren zwei zentrale Theorien, welche die Frage untersuchen, wie und wann es zur regionalen Organisation der Gehirnstrukturen kommt.
Die Protomap-Theorie (Rakic, 1988)[8] bezieht sich auf die pränatale Gehirnentwicklung. Sie besagt, dass durch genetische Instruktionen (Transkriptionsfaktoren) und deren Dosierung bestimmt wird, welche Charakteristiken Neurone aufweisen und welche Funktion sie zukünftig übernehmen. Vereinfacht führt ein Signal über einer bestimmten Schwelle zu anderen Charakteristiken als unterhalb dieser Schwelle.
Demgegenüber steht die Protocortex-Theorie (O’Leary, 1989)[9], welche sich auf die postnatale Gehirnentwicklung bezieht. Verschiedene Regionen des Cortex gelten hier als anfänglich äquivalent und werden erst durch Projektionen vom Thalamus spezialisiert. Diese Projektionen wiederum werden durch postnatale sensorische Erfahrungen beeinflusst. Daraus resultiert die Annahme, dass zu Beginn der Gehirnentwicklung strukturelle Regionen des Cortex ausgetauscht werden können und an ihrer neuen Position ihre Funktion vom Thalamus erhalten. Demnach scheint es sowohl Gen- als auch Umwelteinflüsse auf die strukturelle Organisation des Gehirns zu geben. Die Protomap- und Protocortex-Theorie schließen sich dabei nicht aus.[1]

Funktionelle Gehirnentwicklung
Die funktionelle Gehirnentwicklung beschreibt, wie sich Gehirnfunktionen im Laufe der menschlichen Entwicklung verändern können. Zu diesen zählen zum Beispiel der Erwerb von Wissen und Fähigkeiten.[1]
Funktionelle Plastizität des Gehirns
Es wird angenommen, dass die Reifung und Entwicklung des Gehirns und seiner Bestandteile maßgeblich durch die Genetik und deren Zusammenspiel mit Umwelteinflüssen bestimmt wird – dies gilt insbesondere für die frühen Entwicklungsstadien. Im Laufe des Lebens können sich die Nervenzellen des Gehirns, und somit auch ganze, daraus bestehende, Gehirnregionen, durch Erfahrungen und Umwelteinflüsse anpassen. Dabei ist es zum einen möglich, dass Nervenzellen sich anatomisch verändern (zum Beispiel vermehrt Dendriten ausbilden), was als „neuronale Plastizität“ bezeichnet wird. Zum anderen können sich auch Funktionen von Neuronen oder ganzen Gehirnregionen verändern – das nennt man auch „funktionelle Plastizität“. Kommt es vorgeburtlich zu Fehlbildungen oder nach der Geburt zu Schädigungen von Gehirnregionen (zum Beispiel durch einen Unfall oder Schlaganfall), muss dieser Verlust von funktionierenden Nervenzellen ausgeglichen werden. Es wird jedoch angenommen, dass eine Neubildung von Nervenzellen (Neurogenese) nach der Geburt kaum noch erfolgt. Daher muss sich das Gehirn neu organisieren und die Funktion der geschädigten Gehirnregion unter Umständen einer anderen Gehirnregion zuweisen.[1] In zahlreichen Tierversuchen wurde untersucht, welche Folgen die Transplantation bestimmter Gehirnregionen in andere Bereiche des Gehirns hat. Aus diesen Untersuchungen gingen zwei wichtige Erkenntnisse hervor. Zum einen zeigte sich, dass in eine neue Region transplantierte Nervenzellen, die Funktion dieser Gehirnregion übernahmen und eintreffende Reize korrekt weiterverarbeiteten. Zum anderen zeigte sich, dass eine Schädigung von Nervenzellen in Gehirnregionen der Sinneswahrnehmung auch ohne das Transplantieren von Nervenzellen dazu führt, dass die verbleibenden funktionsfähigen Nervenzellen die ausgefallenen Funktionen übernahmen und somit zum Beispiel Reize aus einem anderen Sinnesorgan verarbeiteten.[1] Auch beim Menschen zeigen verschiedene Fallberichte, dass trotz teilweise schwerwiegender anatomischer Fehlbildungen des Gehirns, oder sogar fehlender Gehirnhälften, die jeweiligen Fähigkeiten der Personen kaum eingeschränkt sein können, da eine funktionelle Anpassung des Gehirns erfolgt sein kann.[1] Solche Fallberichte sollten nicht generalisiert werden, da es sich um Einzelfälle handelt. Diese hohe Anpassungsfähigkeit des Gehirns bringt jedoch auch Nachteile mit sich. In tierexperimentellen Studien zeigte sich beispielsweise die Verarbeitungsleistung von neuorganisierten Nervenzellen der Sinneswahrnehmung als qualitativ schlechter als die der ursprünglich dafür relevanten Nervenzellen. Außerdem ist davon auszugehen, dass nicht alle Nervenzellen universell austausch- und einsetzbar sind.[1] Die wichtigste und weitreichendste Einschränkung ist jedoch, dass die Neuorganisation von Gehirnfunktionen („funktionelle Plastizität“) nur zeitlich begrenzt möglich zu sein scheint und vor allem in sogenannten kritischen oder sensiblen Phasen stattzufinden scheint. Aktuell geht man davon aus, dass die funktionelle Plastizität in den frühen Lebensphasen am größten ist – dies wird auch als Kennard-Prinzip bezeichnet.[1]
Prägung und sensible Phasen
Ausgehend von Konrad Lorenz’ Konzept der kindlichen Prägung wurde in der kognitiven Neurowissenschaft zunächst angenommen, dass die menschliche Entwicklung in sogenannten kritischen Phasen abläuft. Diese bezeichnen eng umgrenzte Zeitfenster, in denen das Individuum relevanten Umweltreizen ausgesetzt sein muss, um bestimmte Lernerfahrungen zu machen und sich folglich weiterzuentwickeln.[2] Werden dieses Zeitfenster ohne die Konfrontation mit diesen Impulsen überschritten, kann die Lernerfahrung nicht mehr gemacht werden.[1]
Gegenwärtig wird in der kognitiven Neurowissenschaft eher vom abgeschwächten Konzept der „sensiblen Phasen“ ausgegangen. Die Basis bilden Studien, die belegen, dass Erlerntes unter bestimmten Bedingungen rückgängig gemacht werden kann bzw. änderbar ist und dass bestimmte Erfahrungen auch außerhalb des sensiblen Zeitfensters nachgeholt werden können. So wird gegenwärtig beispielsweise davon ausgegangen, dass jede Grundfähigkeit, die mit dem Spracherwerb verbunden ist (z. B. Hören, Motorik, Arbeitsgedächtnis), ihre eigene sensible Periode hat. Beispielsweise tritt die phonemische Prägung (z. B. die Unterscheidung zwischen „l“ und „r“) bereits im Säuglingsalter auf. Auch Akzente sind in der Kindheit beeinflussbarer und lassen sich mit zunehmendem Alter nur noch schwer verändern. Gegen diese Befunde sprach zunächst der Fall von Genie. Obwohl Genie während ihres frühen Kindesalters nicht sprechen durfte und nur über ein Vokabular von 20 Worten verfügte, entwickelte sie nach ihrer Befreiung gute Sprachfähigkeiten. Dieser Fall galt zunächst als Beleg gegen eine sensible Phase des Spracherwerb, weitere spätere Berichte sprechen von einem deutlich langsameren Spracherwerb Genies im Vergleich zu dem kleiner Kinder.[1]
Aktuelle Forschungsrichtungen untersuchen zur Beantwortung der offenen Fragen Personen, die zweisprachig aufgewachsen sind. Dabei kann verglichen werden, wie früh die jeweilige Person die zweite Sprache erlernt hat und wie sich dies in ihrer Gehirnaktivität zeigt. Eine Annahme, die die sensiblen Phasen erklärt, nimmt als Ursache “Reifungspläne” a. Eine Möglichkeit der Erklärung besteht dabei darin, dass sich die Plastizität der Neurone im Zuge der Reifung nach der sensiblen Phase verringert. Eine andere Möglichkeit besteht darin, dass bestimmte Neurone zum Lernen angelegt und für einen abgegrenzten Zeitraum bereit für ihre Stimulation sind.[1]
In der kognitiven Neurowissenschaft wird außerdem der Frage nachgegangen, inwiefern die menschliche Entwicklung durch angeborene Wissens- und Verhaltensstrukturen geprägt ist. Dass bestimmte Strukturen angeboren sind, zeigt sich beispielsweise daran, dass bestimmte Wahrnehmungspräferenzen und Fähigkeiten schon seit der Geburt vorhanden sind und sich im Verhalten ausdrücken, ohne dass die entsprechenden Lernerfahrungen bereits gemacht wurden (z. B. Präferenz von süßer Nahrung bei Neugeborenen und Kindern). Es wird davon ausgegangen, dass Lernbereitschaften hinsichtlich bestimmter Aspekte,[10] aber auch neuronale Systeme angeboren sind. Diese neuronalen Systeme können allerdings durch Mangel an Erfahrungen und Impulsen aus der Umwelt abgebaut werden.[2] Die Umwelt und Erfahrungen eines Individuums stellen für dessen Entwicklung eine zentrale Rolle dar. Grund dafür ist, dass die meisten Gene nicht alleinig für eine bestimmte Funktion verantwortlich sind, sondern mit der Umwelt in Wechselwirkung stehen. Diese Anlage-Umwelt-Wechselwirkungen werden vorrangig in der Verhaltensgenetik, zum Beispiel anhand von Zwillingsforschung und Adoptionsstudien, erforscht.
Genotyp-First- vs. Phänotyp-First-Ansatz
Zwei Ansätze werden verwendet, um die Entstehung von Unterschieden hinsichtlich der Gehirnentwicklung zwischen Individuen zu analysieren: der Genotyp-First-Ansatz und der Phänotyp-First-Ansatz.[1]
Der Genotyp-First-Ansatz untersucht, welche Unterschiede im Genotyp mit Unterschieden im Phänotyp assoziiert sind. Beispielsweise werden einzelne Gene, von denen bekannt ist, dass sie in mehreren Varianten (Polymorphismen) existieren, hinsichtlich deren Auswirkungen auf bestimmte Verhaltensweisen untersucht.[1]
Im Gegensatz dazu untersucht der Phänotyp-First-Ansatz, welche Unterschiede im Phänotyp mit Unterschieden im Genotyp assoziiert sind. Beispielsweise werden Merkmale oder Störungen, von denen bekannt ist, dass sie innerhalb der Population variieren, hinsichtlich der Teile des Genoms untersucht, welche am meisten zu der Variation dieser Merkmale oder Störungen beitragen. Dieser Ansatz wird vor allem bei genomweiten Assoziationsstudien (GWAS) verwendet. Diese basieren auf der Annahme, dass viele kleine Variationen im Genom zwischen Individuen bestehen. Diese werden als Einzelnukleotidpolymorphismen bezeichnet. Sie liefern Hinweise darauf, welche Teile des Genoms einen Hotspot enthalten, also in welchen Teilen des Genoms Individuen mit demselben Phänotyp Ähnlichkeiten im Genotyp aufweisen.[1]
Nature-Nurture-Debatte (Natur-Umwelt-Debatte)
Bei der Untersuchung der Gehirnentwicklung stellt sich die grundsätzliche Frage, inwieweit sie durch die Genetik und die Umwelt beeinflusst wird. Dieser Frage widmet sich die Nature-Nurture-Debatte. Dabei werden drei Mechanismen angenommen, durch welche sich Natur und Umwelt beeinflussen: Epigenetik, Gen-Umwelt-Korrelation und Gen-Umwelt-Interaktion.[1]
Während sich die Struktur des genetischen Kodes im Laufe des Lebens nicht verändert, ist die Funktionsweise sehr dynamisch. Verschiedene Gene können aktiv oder inaktiv sein. Ob ein Gen aktiv oder inaktiv ist, hängt auch von der Umwelt ab. Gegenwärtig geht man davon aus, dass Gene unter bestimmten Umständen mit einem chemischen Marker markiert werden, der ihre Expression entweder dämpft oder akzentuiert. Dieser Mechanismus, in welchem die Umwelt die Genexpression beeinflusst, wird als Epigenetik bezeichnet. Ein Beispiel für epigenetische Effekte bei Menschen ist der Einfluss von Vernachlässigung und Missbrauch in der frühkindlichen Entwicklung auf die spätere psychische Gesundheit. Es gibt Hinweise darauf, dass solche frühkindlichen Erfahrungen beispielsweise die Aktivierung von Genen beeinflussen, welche wiederum die physiologische Stressbewältigung beeinflussen.[1]
Genetische Einflüsse wirken sich auf die Exposition von Menschen gegenüber unterschiedlichen Umwelten aus. Menschen mit einem bestimmten Genotyp präferieren bestimmte Umwelten und suchen diese häufiger auf. Dieser Mechanismus wird als Gen-Umwelt-Korrelation bezeichnet.[1]
Die menschliche Entwicklung unterliegt in der Regel sowohl genetischen, als auch Umweltaspekten, deren Zusammenspiel als Gen-Umwelt-Interaktion bezeichnet wird. So beeinflussen die im Rahmen der Gen-Umwelt-Korrelation beschriebenen Prozesse (z. B. das Aufsuchen bestimmter Umweltbedingungen) wiederum die Ausprägung (epi-)genetischer Prozesse. Die Interaktion des Gens und der Umwelt führt somit zu einem größeren Effekt, als aufgrund der Summe der jeweiligen Einzeleffekte des Gens und der Umwelt zu erwarten wäre.[1]
Handlungssteuerung
Des Weiteren befasst sich die kognitive Neurowissenschaft auch mit Prozessen der Handlungsplanung und -durchführung. In einem hierarchisch aufgebauten System von Handlung und Bewegung interagieren verschiedene Ebenen der Wahrnehmung, Kognition und Motorik miteinander. Eine Handlung wird basierend auf Zielen und Intentionen geplant. Außerdem bedarf es perzeptueller, propriozeptorischer und motorischer Systeme, durch die der Mensch die Umwelt wahrnehmen und mit ihr interagieren kann. Das Gehirn berechnet Handlungsabläufe nicht jedes Mal von Grund auf neu. Generalisierte motorische Programme kodieren allgemeine Aspekte der Bewegungen, um eine schnellere Reaktion zu ermöglichen.[11] Der Prozess der Kombination aller handlungsrelevanten Informationen wird als Sensomotorische Integration bezeichnet. Das Ergebnis des Zusammenwirkens aller genannten Prozesse manifestiert sich in einer ausgeführten Handlung.[2]
Frontale Areale
Aufgrund der komplexen Vernetzungen von Wahrnehmung, Kognition und Motorik für die Handlungssteuerung gilt eine Vielzahl von Gehirnarealen als beteiligt. Als zentral für die Handlungssteuerung gelten die Frontallappen. Von posterioren Teilen hin zu den anterioren Bereichen werden ihre Funktionen immer unspezifischer. Anteriore Teile sind eher in die Kontrolle von Verhalten involviert, ohne dabei zwingend zu sichtbaren Handlungen zu führen.
Als posteriorer Teil der Frontallappen steuert der Primäre Motorcortex (M1) die Ausführung von Bewegungen. Er ist somatotopisch organisiert, wobei die rechte Körperhälfte über den linken Teil des Motorcortex gesteuert wird und umgekehrt. Im Gegensatz zu den Bewegungen der Gliedmaßen werden Augenbewegungen nicht vom M1 gesteuert, sondern vom frontalen Augenfeld (engl. frontal eye fields; FEF).
Der Prämotorcortex befindet sich anterior zum M1. Der laterale Prämotorcortex steht vor allem in Verbindung mit Bewegungen, die mit Objekten in der Umgebung zusammenhängen (z. B. nach der Fernbedienung greifen). Er erhält Informationen vom parietalen Cortex über den dorsalen Pfad des Sehens. Der mediale Prämotorcortex (oder auch supplementär-motorisches Areal; SMA) wird mit spontanen, gut gelernten Handlungen assoziiert – insbesondere mit Handlungen, die relativ unabhängig von der Umgebungswahrnehmung sind (also z. B. eine vertraute Melodie auf dem Klavier spielen). Hierfür werden weniger Informationen über die Position von Objekten (dorsaler Pfad) benötigt, stattdessen bezieht das SMA hauptsächlich Signale über die Position der Gliedmaßen.[2]
Präfrontaler Cortex
Der präfrontale Cortex ist in die Planung und in höhere kognitive Aspekte der Handlungskontrolle involviert. Er dient der Auswahl des Prämotorareals (SMA oder lateraler Prämotorcortex) und hält Ziele der Handlung aufrecht. Die Aktivierung des präfrontalen Cortex erfolgt unabhängig von der Bewegung selbst, er ist also lediglich während der Planung und Entscheidung aktiv.[2]
Das intraparietale Areal und frontale Hirnregionen
In einem Netzwerk aus parietalen Arealen und frontalen Regionen werden visuelle, kognitive und motorische Informationen zusammengeführt. Es ist davon auszugehen, dass das anteriore intraparietale Areal und mit ihm in Verbindung stehende frontale Arealen primär abstrakte Eigenschaften einer Handlung kodieren und wahrscheinlich für die Übertragung von Fähigkeiten vom Körper auf ein Werkzeug zuständig sind.
Defizite bezüglich der sensomotorischen Integrationsprozesse können zu verschiedenen Störungen von Handlungsabläufen führen. Die häufigste Form auftretender Schädigungen zieht eine ideomotorische Apraxie nach sich.
Bei der Integration sensorischer und motorischer Informationen sind verschiedene Gehirnareale aktiv. Die beteiligten Neurone kodieren jeweils verschiedene Informationen des Gesamtprozesses.
- Neurone, die die besonderen Aspekte einer Handlung kodieren: Der Mensch besitzt ein gespeichertes Repertoire verschiedener Handlungsabfolgen. Dazu gehören u. a. das Greifen oder das Halten eines Gegenstandes. Feine Fingerbewegungen werden neuronal anders kodiert als ein Griff mit der ganzen Hand.
- Die Kodierung sensorischer Informationen über verschiedene Sinnesmodalitäten hinweg: Es gibt Neurone, die sowohl auf die gefühlte Position einer Gliedmaße im Raum, als auch auf die visuell wahrgenommene Position reagieren. Das heißt, dass unsere visuelle Wahrnehmung immer relativ zur Körperstellung verarbeitet wird.
- Neurone, die handlungsrelevante Eigenschaften von Objekten kodieren: Diese Neurone reagieren v. a. auf die Form der Objekte, deren Größe und deren Ausrichtung im Raum. Lokalisiert sind diese Neurone im anterioren intraparietalen Areal, das vorrangig auf veränderbare Formen und 3D-Objekte reagiert.[2]
Die Rolle subkortikaler Strukturen
Subkortikale Strukturen sind wichtig für die Vorbereitung und Durchführung von Handlungen. Zwei Haupttypen subkortikaler Schleifen gelten als an der Erzeugung von Bewegungen beteiligt:
- Die Cerebellum-Schleife führt durch das Kleinhirn und ist für die Koordination von Bewegungen zuständig. Sie nutzt sensorische und motorische Informationen, um ein genaues Timing und die Akkuratheit der Bewegungen zu koordinieren.[12]
- Die Basalganglien-Schleife besteht aus fünf verschiedenen Schleifen, die zu unterschiedlichen Strukturen in den Basalganglien und dem Cortex projizieren und verschiedene Aspekte des Verhaltens regulieren. Sie bestehen aus exzitatorischen (erregenden) und inhibitorischen (hemmenden) Pfaden.[13] Basalganglien erzeugen keine Signale zur Bewegung, sondern verändern die Aktivität in frontalen motorischen Strukturen (z. B. SMA) und regulieren damit die Wahrscheinlichkeit und Art der Bewegung (z. B. Kraft). Die relevanteste Schleife, der Motorschaltkreis, verläuft durch dorsale Bereiche der Basalganglien bis in prämotorische Areale und das SMA. Er ist besonders wichtig für die Initiierung und Ausführung intern generierter Bewegungen, Handlungsfolgen und für prozedurales Lernen.[2]
Erkrankungen der Basalganglien
Liegen Fehlfunktionen in den Basalganglien vor, kann es zu Krankheiten kommen, die in hypokinetische oder hyperkinetische Ausprägungen unterteilt werden. Hypokinetische Krankheiten zeichnen sich durch ein reduziertes Vorkommen an spontaner Bewegung aus. Ein Beispiel hierfür ist Morbus Parkinson. Hyperkinetische Krankheiten zeichnen sich durch ein erhöhtes Aufkommen an spontaner Bewegung aus, zum Beispiel Huntington, Tourette oder Zwangsstörung (OCD).[2]
Das Supervisory Attentional System-Modell
Im Rahmen der Kognitiven Neurowissenschaft wurde das Supervisory Attentional System-Modell (SAS-Modell) von Norman und Shallice zur Erklärung der Planung zielgerichteter Handlungen entwickelt. Es beschreibt zwei Systeme, die miteinander arbeiten:
Das Contention Scheduling System wählt aus vielen möglichen Schemata eines zur Handlung aus. Die Auswahl eines Schemas hängt einerseits von der Umgebung (sensorischer Input) ab, die automatische Handlungsmuster aktivieren kann.
Andererseits wird das Contention Scheduling System von einem zweiten System beeinflusst, nämlich dem sogenannten Supervisory Attentional System welches die aktuellen und zukünftigen Ziele/Bedürfnisse der Person repräsentiert. Dieses ist besonders dann aktiv, wenn Situationen die Unterbrechung automatischer Handlungsabläufe und das Ausführen neuer, ungeübter Handlungssequenzen erfordern.
Das Contention Scheduling System verrechnet die Informationen des sensorischen Inputs und des Supervisory Attentional System und wählt das Schema mit der höchsten Aktivierung zur Handlung aus. Dieses sollte die aktuellen Bedürfnisse der Person befriedigen und zu den existierenden Umweltbedingungen passen.
Wenn Regionen des präfrontalen Cortex beschädigt werden, bedeutet dies nicht, dass Bewegungen und Handlungen zwangsläufig beeinträchtigt sind; sie werden nur schlecht organisiert und reflektieren nicht notwendigerweise die Ziele der Person. So können typische Handlungsfehler bei Präfrontalen-Cortex-Läsionen mithilfe des Supervisory Attentional System-Modells erklärt werden: Durch die Läsion kommt es zur Imbalance der Informationen, die ins Contention Scheduling System gelangen. Typische Handlungsfehler sind hier beispielsweise die Perseveration, das Utilisationsverhalten oder die frontale Apraxie.[2]
Handlungen und freier Wille
Im Libet-Experiment wurde gezeigt, dass das motorische Zentrum des Gehirns mit der Vorbereitung einer Bewegung bereits begonnen hat, bevor sich eine Person bewusst wird, dass sie sich für die Ausführung entschieden hat. Eine radikale Interpretation dieser Ergebnisse wäre, dass ein freier Wille nicht existiert. Libets Experimente führten zu kontroversen Diskussionen über den freien Willen.
Handlungsverständnis und -imitation
Es gibt gemäß der Kognitiven Neurowissenschaft zwei Möglichkeiten, beobachtete Handlungen Anderer zu reproduzieren:
- Mimikry findet lediglich über sensomotorische Integration statt. Ziele und Absichten der beobachteten handelnden Person werden nicht ergründet.
- Imitation ist eine komplexere Reproduktionsform. Für diese Art von Prozessen gelten die Spiegelneuron eals zentral. Sie bezeichnen eine Gruppe von Nervenzellen, die sowohl während der Durchführung, als auch der Beobachtung zielgerichteter Handlungen reagiert.[14][15] Spiegelneurone unterscheiden dabei nicht, ob diese vom eigenen Selbst oder anderen Personen durchgeführt werden. Sie scheinen bevorzugt auf präzise zielgerichtete Handlungen zu reagieren, entscheidend ist also die Zweckmäßigkeit der Handlung. Es wird angenommen, dass sich Spiegelneurone beim Menschen hauptsächlich im Broca-Areal (besonders Brodmann-Areal 44) befinden, welches sich bis ins prämotorische Gebiet erstreckt.[16]
Handlungen mit Gegenständen
Das Zusammenspiel verschiedener Sinne macht es möglich, Objekte gezielt und funktional einzusetzen. Die Information darüber, wo sich ein Objekt im Raum befindet, muss hierfür mit motorischen Informationen verknüpft werden, um die ausgeführte Handlung den räumlichen Gegebenheiten anzupassen. Hinzu kommt das Verständnis darüber, mit welchen Funktionen die Gegenstände assoziiert sind.
Ungerleider und Mishkin (1982) beschrieben erstmals, dass die visuelle Verarbeitung von Objekten zwei Pfade beinhaltet: den ventralen und den dorsalen Pfad.[17] Der ventrale Pfad oder auch „Was-Pfad“ erstreckt sich vom Okzipital- zum Temporallappen und ist für die explizite Wahrnehmung eines Objekts zuständig. Läsionen hier führen in der Regel zu einer visuellen Agnosie. Der dorsale Pfad wird dagegen als „Wo-Pfad“ oder manchmal auch als „Wie-Pfad“ bezeichnet und erstreckt sich vom Okzipital- zum Parietallappen. Er ist für die Wahrnehmung handlungsrelevanter Eigenschaften wie Größe oder Position von Objekten zuständig. Läsionen hier haben beispielsweise eine optische Ataxie zur Folge.
Auch bei Menschen ohne Läsionen lassen sich Dissoziationen zwischen der visuellen Wahrnehmung und der visuellen Kontrolle von Handlungen beobachten. Bei visuellen Illusionen wie der Ebbinghaus-Täuschung werden eigentlich gleich große Objekte als unterschiedlich groß wahrgenommen. Die Illusion beeinflusst also die visuelle Wahrnehmung, jedoch nicht die Handlung am Objekt.[2]
Werkzeug im kognitions-neurowissenschaftlichen Sinne
Werkzeuge im kognitions-neurowissenschaftlichen Sinne unterscheiden sich von anderen Objekten darin, dass sie mit bestimmten Handlungsabfolgen und dementsprechenden Funktionen assoziiert werden. Anhand von fMRT-Studien wurde herausgefunden, dass der linke Parietallappen (inklusive des anterioren intraparietalen Areals) und das Broca-Areal nur bei der Betrachtung von Werkzeugen im oben beschriebenen Sinne reagierten, nicht aber auf andere Objektklassen.[18]
Exekutive Funktionen
Ein weiteres wesentliches Forschungsthema der Kognitiven Neurowissenschaft ist menschliche Selbstregulation und -kontrolle, maßgeblich innerhalb der sogenannten Exekutiven Funktionen verordnet.
Exekutive Funktionen des Gehirns gelten als nicht an eine bestimmte Domäne (Gedächtnis, Sprache, Wahrnehmung usw.) gebunden, sondern übernehmen eine metakognitive, überwachende oder kontrollierende Rolle[2].
Neuroanatomische Grundlagen Exekutiver Funktionen
Die Exekutive Funktionen sind vornehmlich mit dem präfrontalen Cortex des Frontallappen assoziiert, wobei auch weitere Regionen eine Rolle spielen könnten. Generell wird dem präfrontalen Cortex Bedeutsamkeit bei der Kontrolle von (v. a. nicht automatisierten) Bewegungen, sowie von kognitiven Prozessen bezüglich mentaler Simulation zugesprochen. Darüber hinaus laufen Kontrollprozesse und Speicherkomponenten des Arbeitsgedächtnisses über den präfrontalen Cortex[2].
Der präfrontale Cortex wird anatomisch in seine laterale, mediale und orbitale Oberfläche unterteilt. Der laterale präfrontale Kortex wird vorwiegend mit sensorischen Inputs assoziiert. Er empfängt visuelle, somatosensorische und auditive Informationen sowie Inputs multimodaler Regionen, die über alle Sinne integriert werden. Im Gegensatz dazu gilt der mediale und orbitale präfrontale Kortex als enger mit medialen Temporallappenstrukturen verbunden, die als zentral für das Langzeitgedächtnis und die Verarbeitung von Emotionen angesehen werden[2].
Im Rahmen vielfacher Untersuchungen konnte zusätzlich eine Aktivierung des dorsalen Teils des Anterioren Cingulären Cortex bei der Ausführung exekutiver Funktionen beobachtet werden. Es wird diskutiert, dass er vorwiegend eine Rolle bei der Erkennung und Überwachung von Fehlern bei der Aufgabenbearbeitung haben könnte[2].
Erforschung von Exekutiven Funktionen
Die Erforschung von Exekutiven Funktionen im Rahmen der Kognitiven Neurowissenschaft passiert beispielsweise mittels Aufgaben zu
- Task Setting (Lösungsgenerierung bei vorgegebenem Start- und Zielpunkt, z. B. mittels der Turm-von-London-Aufgabe)
- Inhibition (Unterdrückung gewohnheitsmäßiger Reaktionen, wie z. B. während des Stroop-Tests gefordert)
- Task-Switching (Wechseln zwischen Lösungsstrategien und damit verbundenen Handlungen bei Veränderung der Aufgabenanforderungen, z. B. mittels des Wisconsin Card Sorting Tests)[19][20]
- Versagen beim Task-Switching, d. h. das Festhalten an alten Handlungs- und Denkmustern resultierend in der Unfähigkeit, zu neuen Lösungsmustern zu finden, wird auch als Perseveration bezeichnet
- Multi-Tasking (mentale Aufrechterhaltung mehrere Ziele gleichzeitig (aber nur Ausführung eines), z. B. anhand des Six Elements Tests[21])
- Testpersonen mit präfrontalen Läsionen können separate Aufgaben gut bewältigen, scheitern jedoch bei der Bewältigung mehrerer, gleichzeitig dargebotenen Aufgaben, die sie koordinieren und planen müssen
Theorien zu Exekutiven Funktionen der Kognitiven Neurowissenschaften
Im Rahmen der Kognitiven Neurowissenschaft sind viele Ansätze und Modelle entstanden, um die Struktur der exekutiven Funktionen zu erklären. Diese unterscheiden in dem Ausmaß, in welchem die Exekutivfunktionen in bausteinartige Prozesse unterteilt bzw. alseinheitlicher Bereich ausgelegt werden.
„Heiße“ vs. „kalte“ Kontrollprozesse
Als heißen Kontrollprozess bezeichnet man die Kontrolle von Kognition und Verhalten durch affektive, gewinn- bzw. belohnungsbezogene Stimuli (z. B. Geld beim Menschen, Futter bei Tieren). Diese Prozesse erfolgen automatisch und gelten als hauptsächlich mit dem orbitofrontalen Cortex, aber auch mit dem ventromedialen präfrontalen Cortex assoziiert.
Kalte Kontrollprozesse hingegen, die vorwiegend mit dem lateralen präfrontalen Cortex assoziiert werden, beziehen sich ausschließlich auf kognitive Stimuli (wie z. B. auf die sensorischen Ebenen Farbe und Form).[2]
Weiterführend bildet die „Hypothese der somatischen Marker“ von Damasio (1996)[22] bildet einen Erklärungsansatz für Patienten, die aufgrund einer Gehirnläsion des präfrontalen Cortex Schwierigkeiten in ihrer Verhaltensregulation zeigen, obwohl sie Tests bezüglich ihrer „kalten“ Prozesse bestanden haben. Laut dieser Hypothese verlinken somatische Marker frühere Situationen (welche im Cortex abgespeichert wurden) mit dem dazugehörigen Gefühl (gespeichert in entsprechenden Regionen, z. B. der Amygdala) und dem körperlichen Zustand (z. B. in der Insula). Die somatischen Marker werden vermutlich im ventromedialen Frontalcortex gespeichert und spielen eine zentrale Rolle in der Kontrolle des aktuellen Verhaltens.
The Multiple Demand Network
Eine weitere zentrale Frage der kognitiven Neurowissenschaft ist, ob der laterale präfrontale Cortex in weitere funktionale Untereinheiten zu gliedern ist. Eine mögliche Antwort auf diese Frage liefert Duncan mit der Theorie des Multiple Demand Networks. Das Multiple Demand Network umfasst laut Theorie Regionen des lateralen präfrontale Cortex, des anterioren cingulären Cortex, sowie des Parietallappens (insbesondere jene um den intraparietalen Sulcus). Außerdem ist es eng mit dem Konzept der fluiden Intelligenz assoziiert, da bei zugehörigen Tests (z. B. Raven’s Matrizen) ein sehr ähnliches neuronales Aktivierungsmuster im fMRT zu verzeichnen ist (Duncan, 2010[23]; Woolgar et al., 2010[24]). Laut aktuellem Forschungsstand nutzen alle Exekutivfunktionen dasselbe Netzwerk. Daher wird das Multiple Demand Network als undifferenzierte Einheit charakterisiert, die Theorie verneint also weitere funktionelle Untereinheiten im lateralen präfrontale Cortex[2].
Organisation: posterior nach anterior?
Ein weiterer Forschungsfokus in diesem Kontext ist die hierarchische Organisation des präfrontalen Cortex. Je nach Komplexität einer ausgeführten Aufgabe werden unterschiedliche Areale in diesem aktiviert. Betrachtet man den anterioren Teil des präfrontalen Cortex, so ist dieser bei Multitasking aktiv. Führt eine Person die gleichen Aufgaben nacheinander aus, so bleibt der anteriore Teil des präfrontalen Cortex inaktiv, während posteriore Areale des präfrontalen Cortex aktiv sind[2].
Auf Basis dieser Erkenntnisse stellen Koechlin und Summerfield (2007)[25] die Theorie der hierarchischen Organisation (posterior nach anterior) exekutiver Funktionen auf.
Hierbei sind laut der Theorie überwiegend posteriore Areale bei der Implementierung einfacher Einzelstimuli aktiv („bei grün die linke Taste drücken“). Bei komplexeren Stimuli („bei grün die linke Taste drücken, jedoch nur, wenn ein Vokal vorhanden ist“) werden überwiegend anteriore Areale des präfrontalen Cortex aktiviert.
Hemisphärische Unterschiede
Es wird kontrovers diskutiert ob funktionelle Unterschiede zwischen den Hemisphären bestehen.
In einem Modell von Stuss und Kollegen auf Basis von Läsionsstudien (1995)[26] gilt der linke laterale präfrontale Cortex als spezialisiert für Task-Setting, während der rechte laterale präfrontale Cortex auf Task Monitoring spezialisiert ist.
Eine alternative Sicht auf die Funktion des rechten (inferioren) lateralen präfrontalen Cortex ist, dass er funktionell auf Inhibition spezialisiert ist[2].
Die Rolle des anterioren cingulären Kortex (ACC)
Gegenwärtig wird angenommen, dass der ACC für die exekutiven Funktionen vor allem eine Rolle spielt, wenn es um die Erkennung von Fehlern geht. Es scheint einen kognitiven Mechanismus zu geben, der für die Wahrnehmung von Fehlern sensitiv ist und die Aufgabenleistungen neu kalibriert. Dies zeigt sich im Verhalten beispielsweise durch den typischen Befund, dass Versuchspersonen bei Laboraufgaben unmittelbar nach einem Fehler langsamer und genauer arbeiten. Es ist noch unklar, ob der ACC nur für die Erkennung des Fehlers, oder auch für das kompensatorische Verhalten danach wichtig ist.[1]
Weiterhin wird diskutiert, ob der ACC auch an anderen exekutiven Funktionen beteiligt ist. Solche möglichen Funktionen sind die Bewertung von Antwortkonflikten (wie beim Stroop-Test) und motivationale Prozesse, wie monetäre Belohnungen oder Verluste.[1]
Rolle der Kognitiven Neurowissenschaft für Emotionstheorien
Auch in Hinblick auf Emotionswahrnehmung und soziales Verhalten konnte die Kognitive Neurowissenschaft wesentliche Beiträge leisten. So bestanden schon früh Forschung zum Zusammenhang physiologischer Prozesse und Emotionswahrnehmung.
Hervorzuheben ist dabei die James-Lange Theorie, sie beschreibt die Entstehung von Emotionen. Laut Theorie folgt auf einen Stimulus erst eine körperliche Reaktion (z. B. Muskelanspannung, Veränderungen des Blutdrucks…) und dann das emotionale Erleben durch das Wahrnehmen dieser Reaktion. Jedoch ließ sich feststellen, dass emotionales Erleben auch ohne körperliche Reaktion möglich ist.[27] Der Mediziner Walter Cannon argumentierte außerdem, dass die Körperreaktionen nicht die Unterschiede im Emotionsspektrum erklären könnten. Aus diesen Erkenntnissen entstand die Cannon-Bard-Theorie. Laut dieser Theorie findet das emotionale Erleben im Gehirn vor der körperlichen Reaktion statt. Das neuronale Emotionszentrum sollte dabei der Hypothalamus sein.
Paul Ekman kategorisierte Emotionen in sogenannte Basisemotionen (Freude, Wut, Ekel, Furcht, Verachtung, Traurigkeit und Überraschung), deren Ursprung er vorwiegend in der Genetik des Menschen sah. Außerdem nimmt Ekman an, dass jede Basisemotion ein distinktes neuronales Korrelat besitzt, das ließ sich jedoch nicht bestätigen.
Neuronale Korrelate der Emotionsverarbeitung
Die Kognitiven Neurowissenschaften sehen für die neuronale Verarbeitung von sozialen und emotionalen Prozessen insbesondere folgende Hirnregionen als relevant an: die Amygdala, die Insula, der orbitofrontale Cortex, das anteriore Cingulum und das ventrale Striatum.[28]
Die Amygdala wird vor allem im Kontext des Erinnerns von emotionalen Erfahrungen diskutiert. Dieser Einfluss wird vor allem in Experimenten zu Furchtkonditionierung deutlich. Man geht davon aus, dass eine Aktivierung der Amygdala eine Rückmeldung darüber gibt, ob ein Stimulus oder Verhalten belohnt oder bestraft wird. In der Vergangenheit hatte die Amygdala den Ruf, das Furchtzentrum des Gehirns zu sein. Inzwischen geht man jedoch mehr davon aus, dass sie Teil eines größeren Netzwerkes ist, welches Furcht en- und dekodiert.
Hinsichtlich der Insula zeigen einige Befunde Zusammenhänge mit Aspekten der Schmerz- und Geschmackswahrnehmung. So kann es dazu kommen, dass bei einigen Insulaläsionen die Emotion Ekel in Gesichtern anderer Menschen weniger gut erkannt wird.
Der orbitofrontale Cortex gilt als zentral für die Bewertung wahrgenommener Stimuli im Kontext der aktuellen Situation. Dabei spielt er eine große Rolle beim Verlernen konditionierter Stimuli (Extinktion).
Das anteriore Cingulum gilt als maßgeblich für die Einschätzung, welches Verhalten eine Belohnung und welches eine Bestrafung nach sich zieht.
Das ventrale Striatum ist Teil des dopaminergen Systems und wird mit der Enkodierung und Antizipation von Belohnungen in Zusammenhang gebracht. Eine Abweichung von der Erwartung geht mit einer veränderten Aktivität des Striatums einher.
Emotionserkennung
Gesichter transportieren wichtige Informationen darüber, wie sich jemand fühlt und was jemand plant. Neben diesen sozialen Informationen muss natürlich auch die Identität des Gegenübers festgestellt werden. Es gibt zwei Modelle, die beide annehmen, dass es dafür zwei unterschiedliche Mechanismen gibt.
In der “Single-Route-Detection” (Bruce und Young, 1986)[29] gehen die Experten davon aus, dass es fest zugeordnete Routen für jeden einzelnen emotionalen Gesichtsausdruck gibt. Im neuroanatomischen Modell von Haxby (2000)[30] unterscheidet man zwischen zeitlich gleichbleibenden Repräsentationen von Gesichtern und zeitlich veränderbaren Repräsentationen des Gesichts. Zeitlich gleichbleibende Repräsentationen sind wichtig, um die Identität einer Person zu erkennen. Diese sind in der Fusiformen Face Area gespeichert. Zeitlich veränderbare Gesichtsausdrücke hingegen werden zum Wiedererkennen von Gesichtsausdrücken benötigt und sind im superioren temporalen Sulcus (STS) zu verorten.
Es gibt zudem ein übergeordnetes System zum Erkennen von Emotionen (Heberlein und Adolphs „Simulation theory“). Nach dieser Theorie aktivieren wir zum Verständnis einer Emotion eines Anderen denselben affektiven Pfad, um das entsprechende Gefühl in uns zu reproduzieren.
Zudem werden auch die Muskeln aktiviert, die die jeweilige Emotion als Ausdruck im Gesicht anzeigen. Nicht nur das Sehen, sondern auch die Aktivierung der Muskeln, die für die Emotion typisch sind, führt zum Erkennen der Emotion des Anderen.
Das Erkennen von Gesichtsausdrücken wird zudem verwendet, um das eigene Verhalten anzupassen. So kann bei dem sogenannten „Social Referencing“ eine emotionale Reaktion einer anderen Person (zum Beispiel die der eigenen Mutter) auf einen vorher neutralen Sinneseindruck, dazu führen, dass man sich annähert oder abwendet.
Empathie und die Theory of Mind (ToM)
Auch hinsichtlich Empathie, des affektiven, sowie Theory of Mind, des kognitiven Bestandteils des Nachempfindens von mentalen Zuständen anderer Personen, bestehen wichtige Erkenntnisse der Kognitiven Neurowissenschaft. Singer et al. (2004) konnten beispielsweise feststellen, dass man die Schmerzen, die eine geliebte andere Person hat, dahingehend teilt, dass nicht nur die Hirnregionen von antizipierten Schmerzen anderer aktiv sind, sondern genauso auch die eigenen Schmerzregionen.[31] Man empfindet den Schmerz anderer also in der Region für eigene Schmerzen nach. Über fMRT-Experimente und Läsionsstudien wurden insbesondere drei wichtige Hirnregionen festgestellt, die im Zusammenhang mit dem Nachdenken über mentale Zustände stehen: der mediale präfrontale Kortex, der temporale Pol sowie der Übergangsbereich vom Temporallappen zum Parietallappen.[32]
Spezielle weitere Forschungsbereiche der kognitiven Neurowissenschaft
Literatur
- Bryan Kolb, Ian Whishaw: Neuropsychologie. Spektrum Akademischer Verlag, Heidelberg 1996, ISBN 3-8274-0052-X
- Lutz Jäncke: Einführung in die Kognitiven Neurowissenschaften. Huber-Hogrefe-Verlag, Bern 2013, ISBN 978-3-456-85004-7
- Jamie Ward: The Student's Guide to Cognitive Neuroscience. 3. Auflage. Taylor & Francis Ltd., 2015, ISBN 978-1-84872-272-9.
Einzelnachweise
- Jamie Ward: The student's guide to cognitive neuroscience. Fourth edition Auflage. Milton Park, Abingdon, Oxon 2020, ISBN 978-1-138-49052-9.
- Jamie Ward: The Student's Guide to Cognitive Neuroscience. 3. Auflage. Taylor & Francis Ltd., 2015, ISBN 978-1-84872-272-9.
- APA PsycNet. Abgerufen am 29. Juni 2020.
- H. U. Wittchen, F. Jacobi, J. Rehm, A. Gustavsson, M. Svensson: The size and burden of mental disorders and other disorders of the brain in Europe 2010. In: European Neuropsychopharmacology. Band 21, Nr. 9, 1. September 2011, ISSN 0924-977X, S. 655–679, doi:10.1016/j.euroneuro.2011.07.018 (sciencedirect.com [abgerufen am 29. Juni 2020]).
- Ward, Jamie, 1972-: The student's guide to cognitive neuroscience. Third edition Auflage. London 2015, ISBN 978-1-84872-271-2.
- Bildgebung: Der Traum vom Gedankenlesen. Abgerufen am 25. Juni 2020.
- Gottlieb, G. (1992). Individual development and evolution. New York: Oxford University Press.
- Rakic, P. (1988). Specification of cerebral cortical areas. Science, 241, S. 170–176.
- O’Leary, D. D. M. (1989). Do cortical areas emerge from a protocortex? Trends in Neurosciences, 12, S. 400–406.
- Martin E.P. Seligman: Phobias and preparedness. In: Behavior Therapy. Band 2, Nr. 3, Juli 1971, ISSN 0005-7894, S. 307–320, doi:10.1016/s0005-7894(71)80064-3.
- Richard A. Schmidt: A schema theory of discrete motor skill learning. In: Psychological Review. Band 82, Nr. 4, Juli 1975, ISSN 1939-1471, S. 225–260, doi:10.1037/h0076770.
- Tatsuya Ohyama, William L. Nores, Matthew Murphy, Michael D. Mauk: What the cerebellum computes. In: Trends in Neurosciences. Band 26, Nr. 4, April 2003, S. 222–227, doi:10.1016/S0166-2236(03)00054-7.
- G.E. Alexander, M.D. Crutcher: Parallel processing within motor areas of cerebral cortex and basal ganglia in the monkey. In: 1990 IJCNN International Joint Conference on Neural Networks. Juni 1990, S. 711–716 vol.2, doi:10.1109/IJCNN.1990.137784 (ieee.org [abgerufen am 29. Juni 2020]).
- G. di Pellegrino, L. Fadiga, L. Fogassi, V. Gallese, G. Rizzolatti: Understanding motor events: a neurophysiological study. In: Experimental Brain Research. Band 91, Nr. 1, Oktober 1992, ISSN 0014-4819, S. 176–180, doi:10.1007/BF00230027.
- Giacomo Rizzolatti, Luciano Fadiga, Vittorio Gallese, Leonardo Fogassi: Premotor cortex and the recognition of motor actions. In: Cognitive Brain Research. Band 3, Nr. 2, März 1996, S. 131–141, doi:10.1016/0926-6410(95)00038-0.
- Giacomo Rizzolatti, Leonardo Fogassi, Vittorio Gallese: Motor and cognitive functions of the ventral premotor cortex. In: Current Opinion in Neurobiology. Band 12, Nr. 2, April 2002, S. 149–154, doi:10.1016/S0959-4388(02)00308-2.
- Jason W. Flindall, Claudia L.R. Gonzalez: Revisiting Ungerleider and Mishkin: Two cortical visual systems. In: Brain and Behaviour: Revisiting the Classic Studies. SAGE Publications Ltd, 1 Oliver’s Yard, 55 City Road London EC1Y 1SP 2017, ISBN 978-1-4462-9652-3, S. 45–54, doi:10.4135/9781529715064.n5 (sagepub.com [abgerufen am 29. Juni 2020]).
- Linda L. Chao, Alex Martin: Representation of Manipulable Man-Made Objects in the Dorsal Stream. In: NeuroImage. Band 12, Nr. 4, Oktober 2000, S. 478–484, doi:10.1006/nimg.2000.0635 (elsevier.com [abgerufen am 29. Juni 2020]).
- Brenda Milner: Effects of Different Brain Lesions on Card Sorting: The Role of the Frontal Lobes. In: Archives of Neurology. Band 9, Nr. 1, 1. Juli 1963, ISSN 0003-9942, S. 90, doi:10.1001/archneur.1963.00460070100010.
- Hazel E. Nelson: A Modified Card Sorting Test Sensitive to Frontal Lobe Defects. In: Cortex. Band 12, Nr. 4, Dezember 1976, S. 313–324, doi:10.1016/S0010-9452(76)80035-4.
- Tim Shallice, Paul W. Burgess: Deficits in strategy application following frontal lobe damage in man. In: Brain. Band 114, Nr. 2, 1991, ISSN 0006-8950, S. 727–741, doi:10.1093/brain/114.2.727.
- Damasio, A. R.: The somatic marker hypothesis and the possible functions of the prefrontal cortex. In: Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. Band 351, Nr. 1346, 29. Oktober 1996, ISSN 0962-8436, S. 1413–1420, doi:10.1098/rstb.1996.0125.
- John Duncan: The multiple-demand (MD) system of the primate brain: mental programs for intelligent behaviour. In: Trends in Cognitive Sciences. Band 14, Nr. 4, April 2010, ISSN 1364-6613, S. 172–179, doi:10.1016/j.tics.2010.01.004.
- A. Woolgar, A. Parr, R. Cusack, R. Thompson, I. Nimmo-Smith: Fluid intelligence loss linked to restricted regions of damage within frontal and parietal cortex. In: Proceedings of the National Academy of Sciences. Band 107, Nr. 33, 2. August 2010, ISSN 0027-8424, S. 14899–14902, doi:10.1073/pnas.1007928107.
- Etienne Koechlin, Christopher Summerfield: An information theoretical approach to prefrontal executive function. In: Trends in Cognitive Sciences. Band 11, Nr. 6, Juni 2007, ISSN 1364-6613, S. 229–235, doi:10.1016/j.tics.2007.04.005.
- Stuss, D. T., Shallice, T., Alexander, M. P., & Picton, T. W.: A Multidisciplinary Approach to Anterior Attentional Functions. In: Annals of the New York Academy of Sciences. Band 769, Structure and Functions of the Human Prefrontal Cortex, Dezember 1995, ISSN 0077-8923, S. 191–212, doi:10.1111/j.1749-6632.1995.tb38140.x.
- Fritsch, G., & Hitzig, E.: Ueber die elektrische Erregbarkeit des Grosshirns. 1870.
- Jamie Ward: The Student's Guide to Cognitive Neuroscience. 3. Auflage. Taylor & Francis Ltd., 2015, ISBN 978-1-84872-272-9, S. 382–391.
- Vicki Bruce: Influences of Familiarity on the Processing of Faces. In: Perception. Band 15, Nr. 4, August 1986, ISSN 0301-0066, S. 387–397, doi:10.1068/p150387.
- James V. Haxby, Elizabeth A. Hoffman, M. Ida Gobbini, James V. Haxby, Elizabeth A. Hoffman: The distributed human neural system for face perception. In: Trends in Cognitive Sciences. Band 4, Nr. 6, 1. Juni 2000, ISSN 1364-6613, S. 223–233, doi:10.1016/S1364-6613(00)01482-0, PMID 10827445.
- Jamie Ward: The Student's Guide to Cognitive Neuroscience. 3. Auflage. Taylor & Francis Ltd., 2015, ISBN 978-1-84872-272-9, S. 397.
- Jamie Ward: The Student's Guide to Cognitive Neuroscience. 3. Auflage. Taylor & Francis Ltd., 2015, ISBN 978-1-84872-272-9, S. 404.