Chordatiere
Die Chordatiere ([ˈkɔrda]-; Chordata) sind ein Stamm des Tierreichs. Zu den Chordatieren gehören der Unterstamm der Schädel- (Craniota) oder Wirbeltiere (Vertebrata), und damit auch die Säugetiere (Mammalia) einschließlich des Menschen, sowie zwei weniger bekannte, ausschließlich im Meer lebende Unterstämme, die Schädellosen (Cephalochordata oder Acrania) und die Manteltiere (Tunicata oder Urochordata).
| Chordatiere | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Beim durchscheinenden Sternflecksalmler ist die Wirbelsäule zu sehen, die bei fortgeschrittenen Chordatieren die Chorda dorsalis ersetzt. | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Chordata | ||||||||||||
| Dallas, 1875 |
Zum Stamm der Chordatiere zählen über 56.000 Arten (54.711 Wirbeltierarten,[1] 30 Arten Schädellose und 1.600 Manteltierarten), von denen mehr als die Hälfte – hauptsächlich Knochenfischarten – im Wasser leben.
Alle Chordatiere zeigen eine Reihe gemeinsamer abgeleiteter Merkmale (Synapomorphien). Diese können bei adulten Wirbel- und Manteltieren mehr oder weniger stark abgewandelt sein, sind an den Larven bzw. Embryos aber noch deutlich zu erkennen. Gemeinsam ist den Chordaten die namensgebende Chorda dorsalis (ein stabförmiger Stützapparat im Rücken), das Neuralrohr (ein oberhalb der Chorda liegender Nervenstrang), der Kiemendarm (der zum Filterapparat erweiterte Vorderdarm), das bauchseitig gelegene Herz, das das Blut nach vorne zum Kopf pumpt, und der unterhalb der Chorda liegende Darm, der auf der Bauchseite nach außen mündet und dahinter Platz für den postanalen Schwanz schafft.
Begriff
Die Entwicklung des Begriffs Chordata (Rückensaitentiere, Chordatiere) ging in drei Schritten durch alle drei großen europäischen Wissenschaftssprachen des 19. Jahrhunderts. Der Anfang befand sich in einem Buch des sehr einflussreichen deutschen Zoologen Ernst Haeckel aus dem Jahr 1874.[2]
„Achte Ahnenstufe: Chordathiere (Chordonia). Die Ascidien-Larve als Schattenbild der Chordonier. Ausbildung des Achsenstabes oder der Chorda. Mantelthiere und Wirbelthiere als divergente Zweige der Chordonier.“
Ernst Haeckel hatte das eingedeutschte Wort „Chordathiere“ erfunden und ihm zugleich ein latinisiertes Synonym zur Seite gestellt. Dieses Fremdwort lautete aber nicht „Chordata“, sondern „Chordonia“. Ein paar Seiten später erläuterte Haeckel, was er unter „Chordathieren“/„Chordonia“ genau verstand.
„Wir wollen diese Wurmform, welche vor allem durch den Besitz des Achsenstabes oder der Chorda charakterisiert war, einstweilen als Chordathier (Chordonium) bezeichnen. Als zwei divergierende Linien haben sich aus diesen Chordoniern einerseits die Ascidien, andererseits die Wirbelthiere entwickelt.“
Aus dem Zitat ging hervor, dass Haeckel mit dem Ausdruck „Chordathiere“/„Chordonia“ keine rezenten Organismen meinte, sondern die ausgestorbenen Vorfahren der späteren Wirbeltiere (Vertebrata) und Manteltiere (Tunicata). Seiner Ansicht nach gab es keine lebenden Chordathiere/Chordonia. Dabei zählte Haeckel auch Lanzettfischchen zu den Wirbeltieren,[5] die sich ebenfalls erst aus Chordathieren/Chordonia entwickelt hatten.[6] Haeckels feiner begrifflicher Unterschied zwischen ausgestorbenen Chordathieren/Chordonia und späteren Chordathieren/Chordonia-Nachfahren (Wirbeltiere und Manteltiere) ging jedoch bereits 1875 verloren. In jenem Jahr veröffentlichte sein ehemaliger Schweizer Student Hermann Fol eine kurze Abhandlung zur Herkunft der Keimzellen.
„L’origine première du testicule et de l’ovaire dans les deux feuillets primitifs de l’embryon est maintenant constatée avec certitude chez des exemples tirés de deux embranchements du règne animal, les Coelentérés et les Mollusques; elle est rendue fort probable pour des exemples tirés des deux divisions de l’embranchement des Chordés, à savoir les Tuniciers et les Vertébrés.“
Mit Fols französischer Abhandlung wurde das erste Mal „Chordés“ als zusammenfassende Gruppenbezeichnung für Manteltiere („Tuniciers“) und Wirbeltiere („Vertébrés“) gebraucht. Hermann Fol wich mit dem Wort also von der Auffassung seines früheren Professors ab: „Chordés, à savoir les Tuniciers et les Vertébrés.“ Aber nicht nur die begriffsinhaltliche Verschiebung, sondern auch die Wortform „Chordés“ beeinflusste die weitere Begriffsgeschichte. Denn die französische Wortneuschöpfung hätte sich sowohl auf „Chordathiere“ als auch genauso auf „Chordonia“ beziehen können. Bis 1875 war eine französische Wortform von „Chordathiere“ jedoch noch nicht eingeführt worden. Und bloß wenige Monate zuvor war erstmals und in zwei Sätzen „Chordonia“ in einem Referat vor der Association Française pour l’avancement des Sciences gebraucht worden,[8] der Hermann Fol allerdings nicht angehörte.[9] Gemäß Hermann Fols Abhandlung bezeichnete „Chordés“ nun jene Gruppe von Tieren, die Manteltiere und Wirbeltiere umfasste. Sein Text gelangte sehr bald in die Hände des britischen Zoologen William Sweetland Dallas, der umgehend eine Übersetzung anfertigte, die noch im gleichen Jahr erschien.[10]
„The primary origin of the testis and the ovary in the two primitive lamellae of the embryo is now ascertained positively in cases taken from two great divisions of the animal kingdom, the Coelenterata and the Mollusca; it is rendered very probable by examples taken from the two divisions of the section of the Chordata – namely, the Tunicata and the Vertebrata.“
William Sweetland Dallas hatte entschieden, dass das französische „Chordés“ von Haeckels deutschem „Chordathieren“ abgeleitet worden war. Damit hatte er allerdings gleichzeitig die Möglichkeit verworfen, hier Haeckels internationalisierbares „Chordonia“ zu gebrauchen. Also latinisierte der britische Zoologe „Chordathiere“ einfach nochmals – nun zum Wort „Chordata“. Und innerhalb des Übersetzungstextes verwendete er „Chordata“ zwangsläufig im Sinn von Hermann Fol: Im Jahr 1875 hatte William Sweetland Dallas den Begriff Chordata (Rückensaitentiere, Chordatiere) mit heutiger Wortform und Wortbedeutung geprägt.
| Autor | Jahr | Wortform | Wortbedeutung |
|---|---|---|---|
| Ernst Haeckel | 1874 | „Chordathiere“ / „Chordonia“ | Vorfahren der Manteltiere und Wirbeltiere (einschließlich Lanzettfischchen) |
| Hermann Fol | 1875 | „Chordés“ | Gruppe der Manteltiere und Wirbeltiere (einschließlich Lanzettfischchen) |
| William Sweetland Dallas | 1875 | „Chordata“ | Gruppe der Manteltiere und Wirbeltiere (einschließlich Lanzettfischchen) |
Merkmale
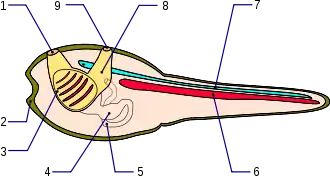
Chorda dorsalis
Die Chorda dorsalis (Latein: Rückensaite), im englischen auch Notochord genannt, ist das namensgebende und Hauptmerkmal der Chordatiere. Sie funktioniert als Endoskelett und ist ein langgestreckter, flexibler Stab, der als Ausstülpung des Urdarms (Archenteron) entstanden ist. Sie liegt deshalb über dem Darm und unter dem Neuralrohr. Sie hat bei allen Chordatieren die gleiche Ontogenese, die durch das Brachyury-Gen gesteuert wird. Es ist nicht feststellbar, ob sich die Chorda ursprünglich nur über den Schwanzbereich oder über den ganzen Körper erstreckte. Seitlich der Chorda finden sich Längsmuskeln, die bei ursprünglichen Chordaten den Körper und den Ruderschwanz wellenartig bewegen können und der Fortbewegung dienen. Während ihrer Ontogenese durchläuft die Chorda eine Münzenstapel-Stadium genannte Phase, in der sie aus scheibenartig zusammengepressten Zellen besteht. Die Chorda der Lanzettfische bleibt in diesem Stadium und wird nur durch Muskelfasern ergänzt. Bei den Larven der Manteltiere und bei den Wirbeltieren entstehen Freiräume zwischen den Zellen, die bei den Larven der Manteltiere ineinander übergehen und einen durchgehenden Kanal bilden, der von Epithelzellen umgeben ist. Bei den Larven der Manteltiere erstreckt sich die Chorda nur über den Schwanzbereich. Die Chorda der Wirbeltiere wird im Laufe der Ontogenese von der Wirbelsäule verdrängt. Ein Überrest der Chorda dorsalis sind die Nuclei pulposi in den Bandscheiben der höheren Wirbeltiere.
Bei den Froschlurchen und verschiedenen Gruppen urtümlicher Landwirbeltiere, u. a. den Temnospondyliern und Reptiliomorphen wie den Anthracosauriern, kann die Bildung der Wirbelkörper im Verlauf der Individualentwicklung sehr stark verzögert sein,[12] so dass bei manchen fossilen Formen, besonders solchen, die auch andere neotene Merkmale aufweisen, noch im Erwachsenenstadium eine Chorda dorsalis als Strang vorliegt und manchmal fossil nachgewiesen werden kann.[13] Häufig weisen die als Fossilien erhaltenen Wirbelkörper dieser Gruppen eine körperlängsachsenparallele Perforation, den Chordalkanal, auf, der die frühere Lage des Chordastrangs anzeigt.

Neuralrohr
Das Neuralrohr entsteht durch die Einstülpung eines länglichen Bereichs des Ektoderms, also einer außen liegende Zellschicht, und befindet sich deshalb zwischen Außenhaut und Chorda dorsalis. Es ist zunächst an beiden Seiten offen und mit der Außenwelt über einen vorderen (rostralen) und kaudalen Neuroporus verbunden. Bei Embryos verbindet der Canalis neurentericus Urdarm und Neuralrohr. Bei den Larven der Manteltiere erstreckt sich das Neuralrohr nur über den vorderen Körperteil.
Herz
Das Herz liegt ventral (auf der Bauchseite) und pumpt das Blut nach vorne zum Kiemendarm. Es fließt dann über dorsal (am Rücken) gelegene Gefäße wieder nach hinten. Bei den Kiemenlochtieren (Hemichordata) und den Urmündern (Protostomia) ist es genau umgekehrt.
Kiemendarm
Der Kiemendarm ist ein zum Filtrieren von Plankton und Detritus umgewandelter Vorderdarm. Mit der Außenwelt steht er durch Öffnungen – den Kiemenspalten – in Verbindung. Hieraus entstehen bei Wirbeltieren die Kiemen, bei Landwirbeltieren zeigt sich der Kiemendarm nur noch embryonal.
Im Unterschied zu den Eichelwürmern, die ihre auf der Körperoberfläche eingeschleimte Nahrung über Kiemenporen aufnehmen und dann durch den Kiemendarm in den Darm saugen, nehmen Chordatiere ihre Nahrung über den Mund auf, filtrieren die Nahrungspartikel im siebartigen Kiemendarm aus, umhüllen sie mit Schleim und geben sie über das Endostyl, einen Streifen von Wimpern- und Drüsenzellen, in den Verdauungstrakt ab. Das mit der Nahrung aufgenommene Wasser wird über den Peribranchialraum und die Kiemenspalten ausgestoßen. Bei den Wirbeltieren – einzige Ausnahme sind hier die den ursprünglichen Zustand bewahrenden Larven der Neunaugen – dienen die Kiemenspalten nur noch zur Atmung. Das Endostyl, das schon bei ursprünglichen Chordaten Iod enthaltende Hormone wie Thyroxin in das Blut abgibt, wurde zur Schilddrüse.
Evolution und äußere Systematik
Die Chordatiere gehören zu den Neumündern (Deuterostomia), denjenigen zweiseitig symmetrisch aufgebauten Tieren (Bilateria), bei denen in der Embryonalentwicklung des Darmes der Urmund (Blastoporus) zum After wird und der Mund sekundär aus dem Urdarm (Archenteron) durchbricht. Die Chordatiere sind innerhalb der Neumünder die Schwestergruppe der Ambulacraria, einem Taxon, das die Stachelhäuter (Echinodermata) und die wurmartigen Kiemenlochtiere (Hemichordata) vereint (siehe nachfolgendes Kladogramm; † = ausgestorben).
| Neumünder |
| |||||||||||||||


Für die Entstehung der Chordatiere aus primitiven Neumündern gibt es zwei Hypothesen:
Tornaria-Hypothese
Gemäß der Tornaria-Hypothese entstanden die Chordatiere aus modifizierten Tornaria-Larven der Kiemenlochtiere (Hemichordata). Die Chorda dorsalis ist nach dieser Theorie homolog zum rückseitigen Nerv der Eichelwürmer (Enteropneusta). Die Chordatiere sind normale Bilateria, deren Anatomie um Chorda, Neuralrohr und Endostyl ergänzt wurde. Ihre Rückenseite entspricht der Rückenseite der Hemichordaten. Die Unterschiede in der dorsoventralen Organisation der Chordatiere und der anderer Bilateria werden so aber nicht erklärt.
Dorso-ventrale Umkehr
Die Hypothese der dorso-ventralen Umkehr hat zur Grundlage, dass die Anordnung der Organe der Chordatiere oft umgekehrt zu der Organanordnung bei Nicht-Chordatieren ist. Sie nimmt eine Drehung des Chordatenkörpers in eine Rückenlage an. So ist vor allem eine der wichtigsten Apomorphien der Chordatiere, die dorsale Lage des Hauptnervenstrangs, zu erklären. Das Nervensystem der meisten anderen Tiere, z. B. der Gliederfüßer und der Ringelwürmer befindet sich ventral (Bauchmark).
Innere Systematik
Die verwandtschaftlichen Beziehungen zwischen den einzelnen Kladen der Chordatiere (Schädellose, Wirbeltiere, Manteltiere) waren lange Zeit umstritten.[14] Man unterscheidet zwei Hypothesen, die ältere Notochordata-Urochordata-Hypothese und die neuere Olfactores-Cephalochordata-Hypothese. Anatomische und molekularbiologische Untersuchungen unterstützen die Olfactores-Cephalochordata-Hypothese,[15][16][17] weshalb sie gegenwärtig als die Wahrscheinlichere angesehen wird. Demnach sind die heutigen Manteltiere stark abgeleitet, während lebende Schädellose viele ursprüngliche Merkmale der Chordatiere beibehalten haben.
Notochordata-Urochordata-Hypothese


Bei der Notochordata-Urochordata-Hypothese sind Schädellose und Wirbeltiere Schwestergruppen. Sie stehen gemeinsam den urtümlicheren Manteltieren gegenüber.[18][19] Dies wird in folgendem Kladogramm dargestellt:
| Chordatiere |
| ||||||||||||
Apomorphien der Notochordata:
- Eine Chorda dorsalis, die sich über den gesamten Körper erstreckt
- Die Segmentierung des Körpers (segmentierte Rumpfmuskulatur und Rückenmarknerven) durch Abschnürung des Mesoderms und des Coeloms aus dorsalen und lateralen Abschnitten der Darmtaschen
- Differenzierter Kopf und Schwanz bei ausgewachsenen Tieren
- Ähnliche Morphologie des Neuralrohrs
- Blutgefäßsystem ist geschlossen
- Ein vertikaler Flossensaum im Bereich des Rückens und des Schwanzes, zwei Metapleuren (laterale Bauchfalten)
- Direkte Entwicklung (mehr oder weniger)
Nach dieser Hypothese hatten die ursprünglichsten Chordaten eine direkte Entwicklung, waren auch als Adulti freischwimmend und die Adulti der Manteltiere entwickelten sich erst sekundär (später) in ihrer Geschichte zu sessilen (festsitzenden) Formen.
Die Merkmale der Notochordata könnten jedoch auch plesiomorphe Merkmale der Chordatiere sein, die bei den Manteltieren sekundär reduziert wurden. Da sie während ihrer Entwicklung über die Hälfte ihrer Hox-Gene verloren haben, ist es wahrscheinlich, dass sie stark vereinfachte Tiere sind. Wirbeltiere und Schädellose entsprächen so eher den ursprünglichen Chordaten, während die Manteltiere durch ihre sekundäre Vereinfachung stark abgeleitet wären. Gegen die Notochordata-Urochordata-Hypothese spricht auch, dass die Schädellosen sehr einheitlich segmentiert sind und in der Ontogenese der Wirbeltiere kein Stadium auftritt, das dem entspricht.
Olfactores-Cephalochordata-Hypothese

Die alternative Olfactores-Cephalochordata-Hypothese sieht die Manteltiere als stark abgeleitete Verwandte der Wirbeltiere. Die Schädellosen sind hier basal.
| Chordatiere |
| ||||||||||||
Apomorphien der Olfactores:
- Die Zellen werden durch Tight Junctions verbunden
- Morphologie der Chorda dorsalis, die auf der Grundlage des Turgordrucks (in den Vakuolen) funktioniert
- Bei der Ontogenese der Muskeln und des Pharynx sind besondere Pax-Gene aktiv
- Neuromasten, das sind Sinneszellen, die bei Wirbeltieren z. B. im Seitenlinienorgan zu finden sind
- Keine Myoepithelien
- Pigmentzellen aus dem Mantel der Manteltiere sind Zellen aus der Neuralleiste der Wirbeltiere sehr ähnlich
Die ursprünglichsten Chordaten könnten, wie die Manteltiere, einen Entwicklungszyklus mit zwei Phasen gehabt haben: eine freischwimmende Larve und ein sessiles Adulttier. In diesem Fall hätten sich die Olfactores (Manteltiere und Wirbeltiere) aus den Schwimmlarven der ursprünglichsten Chordaten entwickelt.
Fossilüberlieferung

Erste fossile Nachweise für Chordatiere gibt es mit †Haikouichthys und †Myllokunmingia schon aus dem Unterkambrium von Yunnan (Maotianshan-Schiefer) und mit †Metaspriggina und †Pikaia aus dem mittelkambrischen Burgess-Schiefer vor ungefähr 505 Millionen Jahren.
Zu den Chordatieren könnten auch die †Vetulicolia gehören, seltsame unterkambrische Tiere, die aus einem voluminösen Vorderteil und einem aus sieben Segmenten bestehenden Hinterteil bestehen. Zwischen Vorder- und Hinterteil sind sie eingeschnürt.
Literatur
- Hynek Burda, Gero Hilken, und Jan Zrzavý: Systematische Zoologie. UTB, Stuttgart, 2008, ISBN 978-3-8252-3119-4
Weblinks
- Lundberg, John G. 1995. Chordata. Version 1. Januar 1995 (under construction). in The Tree of Life Web Project
- Chordata Überblick über die Chordaten und ihre Charakteristika
Einzelnachweise
- Joseph S. Nelson: Fishes of the World. Seiten 17, 28 und 21, John Wiley & Sons, 2006, ISBN 0-471-25031-7
- Claus Nielsen: The authorship of higher chordate taxa. In: Zoologica Scripta. Band 41, 2012, S. 435 doi:10.1111/j.1463-6409.2012.00536.x
- Ernst Haeckel: Anthropogenie oder Entwicklungsgeschichte des Menschen. Verlag Engelmann, Leipzig 1874, S. 398
- Ernst Haeckel: Anthropogenie oder Entwicklungsgeschichte des Menschen. Verlag Engelmann, Leipzig 1874, S. 410
- Ernst Haeckel: Generelle Morphologie der Organismen. Zweiter Band. Verlag Georg Reimer, Berlin 1866, S. X und Tafel VII
- „Zuerst, in früherer archolithischer Zeit, war aller Wahrscheinlichkeit nach das Vertebraten-Phylum bloß durch Leptocardier repräsentiert, von denen uns der einzige lebende Amphioxus noch Kunde gibt. Aus diesen entwickelten sich (innerhalb oder vor der Silur-Zeit) die echten Fische.“ – Ernst Haeckel: Generelle Morphologie der Organismen. Zweiter Band. Verlag Georg Reimer, Berlin 1866, S. CXVIII
- Hermann Fol: Note sur l’origine première des produits sexuels. In: Bibliothèque universelle et Revue suisse – Archives des Sciences Physique et Naturelles. Nr. 209, 1875, S. 111
- „… transmis par les Chordonia aux vertébrés … dans la généalogie des Chordonia …“ – Alfred Mathieu Giard: Note sur quelques points de l’embryogénie des ascidies. In: Association Française pour l’avancement des Science. Compte Rendu de la 3me Session. Lille, 1874. Secrétariat de l’association Française, Paris 1875, S. 453–454
- vergleiche Liste des Membres. In: Association Française pour l’avancement des science. Compte Rendu de la 3me Session. Lille, 1874. Secrétariat de l’association Française, Paris 1875, S. XXV-LVI
- „Translated by W.S. Dallas, F.L.S.“ – Hermann Fol: On the primary Origin of the Sexual Products. In: Annals and Magazine of Natural History. Band 16, 1875, S. 157
- Hermann Fol: On the primary Origin of the Sexual Products. In: Annals and Magazine of Natural History. Band 16, 1875, S. 161
- Robert L. Carroll, Andrew Kuntz, and Kimberley Albright: Vertebral development and amphibian evolution. Evolution and Development 1(1), 1999, S. 36–48.
- Ralf Werneburg: Timeless design – colored pattern of skin in Early Permian branchiosaurids (Temnospondyli: Dissorophoidea). Journal of Vertebrate Paleontology 27 (4), 2007, S. 1047–1050.
- Hynek Burda: Systematische Zoologie. S. 241/242, Eugen Ulmer Stuttgart, 2008, ISBN 978-3-8252-3119-4.
- Delsuc, F., Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature (2006), 439(7079):965–968 doi:10.1038/nature04336 https://hal.archives-ouvertes.fr/halsde-00315436/file/Delsuc-Nature06_HAL.pdf
- Dunn, C.W., Broad phylogenetic sampling improves resolution of the animal tree of life. Nature (2008), 452(7188):745–749 doi:10.1038/nature06614
- Gupta, Radhey S., Molecular signatures that are distinctive characteristics of the vertebrates and chordates and supporting a grouping of vertebrates with the tunicates. Molecular Phylogenetics and Evolution (2016), 94(part A):383–391 doi:10.1016/j.ympev.2015.09.019
- Ax, P., Das System der Metazoa: ein Lehrbuch der phylogenetischen Systematik (2001)
- Stach, T., Chordate phylogeny and evolution: a not so simple three‐taxon problem. Journal of Zoology (2008), 276(2):117–141 doi:10.1111/j.1469-7998.2008.00497.x