Newark-Supergruppe
Die Newark-Supergruppe (englisch Newark Supergroup) ist eine lithostratigraphische Einheit vom Rang einer Supergruppe, die eine, beckenübergreifend kumuliert, maximal rund 15.000 Meter[2] mächtige Abfolge von mindestens der Mittel-Trias bis in den Unteren Jura umfasst. Sie bildet die überwiegend siliziklastisch-sedimentäre, teils mit Basalten wechselgelagerte Füllung zahlreicher größerer und kleinerer fossiler Sedimentbecken an der Ostküste der USA und in den Seeprovinzen Kanadas. Die Ablagerung dieser Sedimente bzw. die Bildung der entsprechenden Becken und der damit einhergehende basaltische Vulkanismus stehen in engem Zusammenhang mit der Frühphase der Öffnung des Atlantischen Ozeans. Als Fundhorizonte der historisch ersten Dinosauriertrittsiegel und -knochen, der ersten weitgehend vollständig erhaltenen Fischfossilien sowie der ersten mesozoischen Insekten in Nordamerika haben die unterjurassischen Schichten der Newark-Supergruppe eine herausragende wissenschaftshistorische Bedeutung.
_USGS_Bulletin_1572.png.webp)
Historisches
Der Name Newark Group wurde im Jahr 1856 von William Charles Redfield geprägt. Zuvor wurden die frühmesozoischen, oft rötlich gefärbten Siliziklastika der Ostküstenstaaten der USA* aufgrund vermuteter Altersgleichheit mit faziell ähnlich ausgebildeten Abfolgen in der Sedimentüberlieferung insbesondere Großbritanniens verschiedentlich bezeichnet. So zunächst als Old Red Sandstone und ab den 1830er Jahren oft als New Red Sandstone†. Redfield hielt den von ihm gewählten Namen für geeigneter, weil sich die paläontologischen Befunde dafür mehrten, dass diese Schichten jünger sind als der New Red Sandstone von Großbritannien.[3] Obwohl sich das im Nachhinein als nur teilweise richtig herausstellte, setzte sich der Name Newark – anfangs auch in den Kombinationen Newark System und Newark Series – im 20. Jahrhundert durch. Israel Russell favorisierte ihn in einer 1892 veröffentlichten Monographie wegen seines regionalen Bezuges, ohne innewohnende Mutmaßung über eine Korrelation mit einer bestimmten Gesteinseinheit Europas.[4] Als solcher erfüllte er schon damals die Benamungsregeln der modernen Lithostratigraphie. Russell (1892) zählte, unter anderem basierend auf James Dwight Dana (1875)[5], auch die Ausbisse in den kanadischen Seeprovinzen zum Verbreitungsgebiet des „Newark System“ hinzu.[4]
Nach der Etablierung der „Theory of New Global Tectonics“ an der Wende von den 1960er zu den 1970er Jahren reifte die Erkenntnis, dass die Genese der Becken und damit auch die Ablagerung der Sedimente der Newark-Gruppe mit dem Auseinanderbrechen der westlichen Pangaea und der Entstehung des Atlantischen Ozeans zusammenhängen.siehe z. B. [6]
Im Jahre 1978 schlug Paul Eric Olsen, der gerade am Peabody-Museum von Yale seinen Geologie-Bachelor erworben hatte, die Heraufstufung der Newark-Gruppe in den Rang einer Supergruppe vor, da inzwischen für einige der Beckenfüllungen eigene Gruppen errichtet worden waren (u. a. die Fundy-Gruppe und Dan-River-Gruppe; vgl. Beckennamen unten) und der Terminus „Newark-Gruppe“ mithin nicht mehr als Überbegriff für alle Beckenfüllungen geeignet war.[7] 1984 erfolgte dann durch Olsen die offizielle Beantragung dieser Heraufstufung und deren Annahme durch den United States Geological Survey.[8] Zahlreiche Publikationen in den folgenden Jahrzehnten sorgten dafür, dass der Name Olsen untrennbar mit der modernen, insbesondere sedimentologischen, stratigraphischen und paläontologischen Erforschung der Newark-Supergruppe verbunden ist.
Verbreitung
.jpg.webp)
Die Gesteine der Newark-Supergruppe beißen diskontinuierlich in einem rund 100 Kilometer breiten Streifen zwischen den Appalachen und der Küstenebene aus, der sich vom Norden South Carolinas bis zum nordwestlichen Ende von Nova Scotia (Chedabucto Bay) erstreckt. Sie verteilen sich dabei auf sieben größere und rund 20 kleinere und sehr kleine strukturelle Reste von Sedimentbecken, die, prinzipiell Südwest-Nordost, parallel zu den Strukturen der Appalachen streichend, in Form von Halbgräben in das prä-mesozoische Grundgebirge eingesenkt sind.[1][9] Die Namen der größeren Becken lauten wie folgt (von Südosten nach Nordwesten):[1][9][10]
- Wadesboro Basin (North Carolina mit geringen Anteilen in South Carolina)
- Sanford-Durham Basin (North Carolina)
- Dan River-Danville Basin (North Carolina, Virginia)
- Culpeper Basin (Virginia, Maryland)
- Gettysburg-Newark Basin (Maryland, Pennsylvania, New Jersey, New York)
- Hartford-Deerfield Basin (Connecticut, Massachusetts, traditionell auch Connecticut (River) Valley genannt, weil der Connecticut River das Becken auf nahezu seiner gesamten Länge durchfließt)
- Fundy Basin (Nova Scotia)
Ablagerungen, die aufgrund ihres Alters und ihres geologischen Kontextes enge genetische Beziehungen zur Newark-Supergruppe aufweisen, finden sich außerdem von Florida und Georgia (South Georgia Basin) bis Neufundland (Jeanne d’Arc-Flemish Pass Basin) im Untergrund der Küstenebene und des vorgelagerten Schelfs.[2][9] In Nova Scotia gehen „On-shore“-Ausbisse und „Off-shore“-Vorkommen sogar unmittelbar lateral ineinander über.[2]
Erdgeschichtlich-regionalgeologischer Rahmen
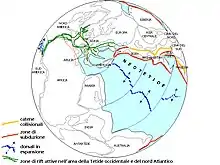
Die Kette von Becken, in denen die Sedimente der Newark-Supergruppe und ihre begrabenen Äquivalente zur Ablagerung kamen, entstand ab der Mitteltrias im Zuge einer Dehnung der Erdkruste im nordamerikanisch-nordafrikanischen Abschnitt des weitgehend abgetragenen variszischen Gebirgsgürtels.[11] Der nordamerikanische Teil des entsprechenden Systems aus Grabenbrüchen (engl. Central Atlantic Margin, abgekürzt CAM, genannt), das als mesozoisches Analogon zum Großen Afrikanischen Grabenbruchsystem gilt,[12] wird als Eastern North American Rift System (ENARS) bezeichnet. Die Dehnungstektonik ist der regionale Ausdruck des Auseinanderbrechens des Superkontinentes Pangaea und stellt die Initialphase der Öffnung des Zentralatlantiks dar.[9][13]
Mit diesem plattentektonischen Ereignis in engen Zusammenhang gebracht werden auch die Basalte, die im Bereich der Trias-Jura-Grenze in die sedimentäre Abfolge eingeschaltet sind. Die Basalte sind Teil der Zentralatlantischen Magmenprovinz (engl. Central Atlantic Magmatic Province, abgekürzt CAMP), die sich auch auf Nordwest- und Westafrika sowie den Nordosten Südamerikas erstreckt.[14]
Mit dem Übergang zur Ozeanbodenspreizung („rift to drift“) im eigentlichen Zentralatlantikbecken im weiteren Verlauf des Unterjura endet die kontinentale Krustendehnung und damit auch die Subsidenz und die Aufnahme von Sedimenten in den Becken der Newark-Supergruppe.[9][13] Die Riftbecken im Osten Nordamerikas erfahren in dieser Phase sogar eine leichte Inversion, bei der sich der Bewegungssinn der vormaligen Abschiebungen umkehrt und sich in einigen der Beckenfüllungen Falten ausbilden.[9] Als Ursache für diese Kompressionsphase werden das Einsetzen des „Ridge Push“ (siehe Ursachen der Plattentektonik) an der atlantischen Spreizungszone und/oder vom nordamerikanischen Kontinentalblock (Laurentia) ausgehende, der Plattenbewegung entgegengerichtete Trägheitskräfte vermutet.[9] Seither wurden sowohl die jeweils jüngsten Schichten der Beckenfüllungen, als auch die wahrscheinlich zwischen den heutigen Beckenresten (Depozentren) ursprünglich vorhandenen geringmächtigen Ablagerungen aufgrund ihrer mehr oder weniger permanenten Lage oberhalb der Erosionsbasis wieder abgetragen.[2][11][12]
Stratigraphie

Die interne lithostratigraphische Gliederung der Newark-Supergruppe war in den vergangenen Jahrzehnten mehrfachen Veränderungen unterworfen. Noch Ende der 1980er Jahre wurden für die Füllung jedes größeren Beckens eigene Formations- und Gruppennamen verwendet.[1][15] Insgesamt wurden 6 Gruppen und 57 Formationen unterschieden (die Formationen kleinerer Becken wurden dabei jeweils nicht in Gruppen zusammengefasst).[11] Im Zuge einer Revision durch Weems & Olsen (1997)[10] wurde die Anzahl der bis dahin ausgehaltenen Gruppen von neun auf drei reduziert und alle noch nicht einer Gruppe zugeordneten Formationen in eine dieser drei Gruppen gestellt. In einer nachfolgenden Revision (Weems et al., 2016)[2] wurde, bei Erhöhung der Anzahl der Gruppen auf vier, die Anzahl der Formationen auf 16 weitgehend beckenübergreifende Einheiten reduziert. Demnach gliedert sich die Newark-Supergruppe wie folgt (von jung nach alt):
- Portland-Gruppe (vormals Agawam-Gruppe**; jüngeres Hettangium – Sinemurium)
|
lateral mit beiden verzahnt: |
- Meriden-Gruppe (jüngstes Rhätium – älteres Hettangium)
- Chatham-Gruppe (Karnium – Rhätium)
- Passaic-Formation
- Lockatong-Formation
- Stockton-Formation
- Doswell-Formation
- Acadia-Gruppe (auf die kanadischen Seeprovinzen beschränkt; ? Oberperm, ? Anisium – ? Karnium)
|
mit unklarer Beziehung |
In North Carolina und weiter südlich sind die Beckenfüllungen auf die Obertrias beschränkt. Als ältestes Schichtglied der Supergruppe galt noch 1997 der untere Teil der Wolfville-Formation („Lower Economy beds“[13]; seinerzeit älteste Schichten der Chatham-Gruppe) des Fundy-Beckens, der bis mindestens in die obere Mittel-Trias hinabreicht.[2][10] Nachfolgend sind kleinräumig an der Fundy-Bay-Küste von New Brunswick aufgeschlossene, und ursprünglich ebenfalls als mitteltriassisch betrachtete Schichten der sogenannten Honeycomb-Point-Formation, die in der 1997er Revision keine Erwähnung finden, aufgrund von paläomagnetischen und palichnologischen Befunden als wahrscheinlich oberpermisch eingestuft und mit der klar permischen Ikakern-Formation des Argana-Beckens in Marokko korreliert worden.[13][16] Die Honeycomb-Point-Formation könnte aber noch vor dem Einsetzen der ENARS-Dehnungstektonik abgelagert worden sein.[17] Als jüngste Ablagerungen gelten allgemein die obersten Schichten des Longmeadow-Sandsteins (vormals obere Portland-Formation, Agawam-Gruppe) im Hartford-(Teil-)Becken.[10][11] Deren genaues Alter ist jedoch unsicher. In jüngeren Publikationen (2000 ff.) werden sie meist ins Sinemurium gestellt,[2][9][18] während Ende der 1980er Jahre noch ein spätunterjurassisches Alter (Toarcium) veranschlagt wurde.[11][15][19]
Lithologien


Sedimentgesteine – Fazies und Zyklen
Die Abfolge der Newark-Supergruppe umfasst mehrheitlich rötliche siliziklastische Gesteine, die als Sedimente seinerzeit entweder durch Flüsse (fluviatil) oder in Seen (lakustrin/limnisch) abgelagert wurden. Nördlich von North Carolina ist im triassischen Abschnitt der Beckenfüllungen in der Regel eine Dreiteilung der Abfolge feststellbar: Auf einen basalen, überwiegend fluviatilen Teil folgt ein Intervall, das von lakustrinen Tiefwasser-Ablagerungen mit oft grauen oder schwarzen Schichten dominiert wird, auf das ein Intervall mit überwiegend lakustrinen, rötlichen Ablagerungen folgt, wobei die Grenzen zwischen diesen Intervallen nicht beckenübergreifend gleich alt (isochron) sind.[11] In den unterjurassischen Abschnitten der Beckenfüllungen ist keine vergleichbar typische vertikale Faziesabfolge erkennbar.
Intervalle mit dominant lakustriner Sedimentation zeigen lateral einen konzentrischen Aufbau. Die feinkörnigsten Ablagerungen (Ton-Silt-Gesteine) finden sich in den relativ zentralen, wegen der Halbgrabengeometrie in Richtung der Beckenrandstörung versetzten Beckenbereichen mit der höchsten Subsidenz. Mit dieser eigentlichen lakustrinen Fazies verzahnen sich nach außen hin sandige fluviatile und deltaische Sedimente, die in Richtung der Beckenrandstörung weiter in Konglomerate übergehen.[11] Vertikal zeigen viele der mächtigen Abfolgen der eigentlichen lakustrinen Fazies einen zyklischen Aufbau, der Milanković-gesteuerte Klimaschwankungen widerspiegelt. Die im Aufschlussmaßstab beobachtbaren Van-Houten-Zyklen sind beckenweit korrelierbare lakustrine Transgressions-Regressions-Zyklen, die dem rund 20.000-jährigen Präzessionszyklus entsprechen. Die erst im Kartenmaßstab wahrnehmbaren, jeweils rund 20 von Van-Houten-Zyklen umfassenden McLaucghlin-Zyklen repräsentieren langfristigere aber extremere Klimaschwankungen im rund 400.000-jährigen Exzentrizitäts-Zyklus.[12][20][21] Innerhalb eines Van-Houten-Zyklus werden drei Abschnitte (engl. divisions) unterschieden:[11][20][21]
- transgressiver Abschnitt – geringmächtig, mit eher massig ausgebildeter Basis, zum Hangenden hin abnehmende Anzahl von Trockenrissen und Wurzelspuren, einhergehend mit deutlicher ausgebildeter Schichtung (Nicht untypisch sind karbonatische Bewegtwasserablagerungen der Uferzone. So enthält eine transgressive Sequenz in der Passaic-Formation in-situ-erhaltene, fusinitisierte, d. h. möglicherweise bei einem Waldbrand verkohlte, kleine Baumstümpfe, die von Stromatolithen überwachsen sind, die wiederum in oolithischem Sand eingebettet sind, und alles wird von schwarzen Siltsteinen mit an- und abschwellender Lamination, engl. „pinch and swell lamination“, überlagert.[22]); dokumentiert zunehmend feuchtes Klima
- Hochstand-Abschnitt – geringmächtig, oft graue oder schwarze, „schiefrig“-plattige, teils laminierte Silt-Tonsteine; dokumentiert einen Zeitraum mit sehr feuchtem Klima; dunkle, laminierte Schichten zeigen tiefe (bis zu 80 Meter) Seen mit geringer Wasserzirkulation und beckenweiter Erstreckung an (wahrscheinlich reichten die Seen deutlich über die Ränder der heutigen strukturellen Beckenreste hinaus, möglicherweise handelte es sich sogar um die „größten Seen aller Zeiten“[12])
- regressiver Abschnitt – mächtig, mit relativ deutlich ausgebildeter Schichtung an der Basis, zum Hangenden hin zunehmende Anzahl von Trockenrissen und Wurzelspuren, einhergehend mit massigerem Erscheinungsbild des Gesteins; dokumentiert zunehmend trockenes Klima; repräsentiert relativ flache, weniger ausgedehnte und regelmäßig zumindest teilweise trockenfallende Gewässer an
Schichten fluviatilen Ursprunges werden überwiegend als Zeugnisse verflochtener Flüsse gedeutet. Entsprechende Ablagerungen liegen meistens in Form rötlich-bräunlicher, kiesiger und/oder arkotischer Sandsteine vor. Nur einige fluviatile Sedimente zeigen Merkmale, die auf Ablagerung durch mäandrierende Flüssen hindeuten, wenngleich dies ein Artefakt sein könnte, das einer relativ schlechten Aufschluss-Situation geschuldet ist.[11] Bei den konglomeratischen Schichten der Beckenränder handelt es sich zumindest teilweise um Fanglomerate, das heißt um Ablagerungen von alluvialen Fächern, in denen grober, unsortierter Abtragungsschutt des Grundgebirges abgesetzt wurde, das aus dem seinerzeit stärker reliefierten Gelände jenseits der Grabenrandstörung eingeschwemmt wurde.[11] Speziell in der Doswell- und Lockatong-Formation (nach neuer Nomenklatur) mehrerer meist kleinerer Becken in North Carolina und Virginia treten, assoziiert mit sowohl fluviatilen als auch lakustrinen Ablagerungen, Steinkohleflöze mit Mächtigkeiten zwischen 5 Zentimetern und 2 Metern auf.[11][23]
Im Fundy-Becken sind zudem äolische Sedimente (Dünensande) relativ weit verbreitet.[11]

Vulkanite

Die Vulkanite der Newark-Supergruppe umfassen Tholeiitbasaltdecken und entsprechende pyroklastische Sedimente. Sie variieren in ihrer relativen chemischen Zusammensetzung und ihrem Mineralbestand nur geringfügig, wobei für jede der drei (nach neuer Nomenklatur) vulkanitdominierten Formationen eine bestimmte Zusammensetzung charakteristisch ist. Diese Zusammensetzungen entsprechen im Wesentlichen den „Chemotypen“, die von Weigand & Ragland (1970)[24] bei frühjurassischen Doleriten im Osten Nordamerikas und von Tollo & Gottfried (1989)[25] beim Hook-Mountain-Basalt (Hampden-Formation nach neuer Nomenklatur) des Newark-Beckens identifiziert wurden.[26] Demnach entspricht der Basalt der Talcott-Formation dem Typ titanreicher quarznormativer Tholeiit (HTQ; mehr als 1,0 Gew.-% TiO2), der chemisch relativ variable Basalt der Holyoke-Formation dem Typ eisenreicher quarznormativer Tholeiit (HFQ; 13–15 Gew.-% total Fe2O3) und im Culpeper-Becken und Newark-Becken auch dem Typ titanarmer quarznormativer Tholeiit (LTQ; 0,7–0,8 Gew.-% TiO2) und der Basalt der Hampden-Formation entspricht dem Typ eisen- und titanreicher quarznormativer Tholeiit (HFTQ; mehr als 15 Gew.-% total Fe2O3 bei mehr als 1,3 Gew.-% TiO2).[2][25][26][27]
Aus deutlichen Unterschieden der Spurenelement-Signaturen der Basalte der drei Formationen, insbesondere der Verteilung sogenannter inkompatibler Elemente (einschließlich SEE), das heißt Elemente, die sehr lange in der Schmelze verbleiben, weil ihre Ionenradien nicht zum Kristallgitter der bevorzugt auskristallisierenden Minerale passen, und deren Gehalt und Verteilung in der Schmelze folglich nicht durch Fraktionierungsvorgänge beeinflusst wird, ist geschlussfolgert worden, dass die Magmen der drei Formationen nicht aus ein und derselben Quelle stammen, sondern aus jeweils verschiedenen Quellen.[25]
Fossilführung

Die Newark-Supergruppe gilt heute als eine der fossilreichsten kontinentalen Schichtenfolgen der Welt.[22] Je nach sedimentärer Fazies bzw. Ablagerungsmilieu enthält sie vorwiegend Landwirbeltiere (Tetrapoden), verschiedenste Fische (weit überwiegend aber mehr oder weniger „primitive“ Strahlenflosser aus den Gruppen Palaeonisciformes bzw. Semionotiformes), terrestrische und aquatische Arthropoden, Süßwassermollusken, Pflanzen, fossile Trittsiegel von Landwirbeltieren und Spurenfossilien von Wirbellosen in verschiedenen Erhaltungszuständen. So enthalten die sandig-konglomeratischen fluviatil-alluvialen Randfazies zwar zahlreiche Landwirbeltiere, jedoch oft nur in schlechter bis moderater Erhaltung (d. h. mehr oder weniger stark abradierte Einzelknochen und Knochenfragmente, die nur selten auf Gattungs- oder gar Artebene bestimmbar oder diagnostizierbar sind). Pflanzenreste kommen dort nur in Form unidentifizierbarer Häcksel vor. Die Stillwasserfazies der zentraleren Beckenbereiche hingegen enthalten Pflanzen, Wirbellose, Fische und einige (aquatische und terrestrische) Landwirbeltiere in guter bis außergewöhnlich guter Erhaltung (Konservatlagerstätten). Trittsiegel finden sich in den feinkörnigeren fluviatilen Ablagerungen (Schwemmebene) und vor allem in den lakustrinen Ablagerungen ufernaher, periodisch trockengefallener Bereiche, wobei sie, einer allgemeinen „Überlieferungsregel“ folgend, kaum bis gar nicht mit fossilen Knochen assoziiert sind.[22]
Die Landwirbeltierfaunen weisen zum einen Beziehungen zu jeweils ungefähr gleichaltrigen Faunen des Colorado-Plateaus (Chinle-Formation, Kayenta-Formation) sowie europäisch-nordafrikanischen („British fissure fills“, Germanisches Becken, Argana-Becken) und chinesischen Faunen, zum anderen aber auch Beziehungen zu gondwanischen Faunen (speziell zu denen des Paraná-Karoo-Beckenkomplexes im südöstlichen Südamerika bzw. im südlichen Afrika) auf.
Da sie im nördlichen Teil ihres Verbreitungsgebietes auch fossilführende Schichten des Unterjura umfasst, bietet die Newark-Supergruppe einen Einblick in das Trias-Jura-Massenaussterben und die nachfolgende Erholungsphase.[28][29][16]
Mitteltrias
Mitteltriassische Wirbeltierfossilgesellschaften kommen nur im Fundy-Becken vor. Hierbei werden eine ältere in der Economy-Formation und eine jüngere in der Evangeline-Formation unterschieden. Die ältere beinhaltet verstreute Reste von auf Gattungsebene nicht eindeutig bestimmbaren „Urlurchen“, genauer trematosauroiden (vormals identifiziert als Aphaneramma oder Cosgriffius) und capitosauroiden Temnospondylen. Amnioten sind durch Procolophonoiden und verschiedene Vertreter der Archosauriformes vertreten, von denen jedoch nur Tanystropheus anhand eines einzelnen Halswirbels eindeutig identifizierbar ist.[16][22]
Eine ähnliche aber doch klar abgrenzbare Fauna enthält die Evangeline-Formation, mit dem metoposauroiden Temnospondylen Metoposaurus bakeri und den Procolophonoiden Acadiella psalidodon, Haligonia bolodon und Scoloparia glyphanodon sowie dem engen Trilophosaurus-Verwandten Teraterpeton hrynewichorum, „höheren“ (hyperodapedontinen) Rhynchosauriern, basalen terrestrischen Krokodillinien-Archosauriern aus der Gruppe der Aetosaurier und der Rauisuchier und mit Arctotraversodon plemmyridon auch einen sicher bestimmbaren Vertreter nicht-mammaler Cynodontier. Material, das vormals Dinosauriern zugeordnet wurde, gehört vermutlich zu anderen, bereits oben genannten Archosauriergruppen und nur in einigen Fällen zu Nicht-Dinosaurier-Ornithodiren.[16] Im Gegensatz zu Weems et al. (2016)[2] datieren Sues & Olsen (2015)[16] die Evangeline-Formation (bzw. die Evangeline-Subformation der Wolfville-Formation) nicht in die späte Mitteltrias (Ladinium), sondern in die frühe Obertrias (Karnium).
Obertrias

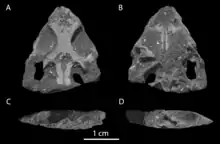
Landwirbeltierfaunen in fluviatil abgelagerten Schichten in der Obertrias der Newark-Supergruppe werden beherrscht durch Procolophonoiden (mit Hypsognathus fenneri als erd- und evolutionsgeschichtlich letztem Vertreter der Gruppe), Phytosaurier, Aetosaurier, Rauisuchier und hyperodapedontine Rhynchosaurier.[22] Dinosaurier bilden nur untergeordnete Faunen-Elemente und sind durch nicht näher bestimmbare fragmentarische Reste von (wahrscheinlich ausschließlich coelophysoiden) Theropoden repräsentiert.[30][31] Das vormals postulierte Vorkommen von „Prosauropoden“ ist für die Obertrias im gesamten Nordamerika aufgrund eines Mangels an diagnostischem Material nicht nachgewiesen.[32] Das ebenfalls vormals postulierte Auftreten von Ornithischiern („Fabrosauridae“, „Galtonia gibbidens “, „Pekinosaurus olseni “) wird mittlerweile sogar für die Obertrias ganz allgemein bezweifelt.[33] Umstritten sind auch die Verwandtschaftsbeziehungen des 1993 beschriebenen Fundes des Schädels eines kleinen juvenilen Reptils aus der New-Haven-Arkose (Passaic-Formation des Hartford-Deerfield-Beckens). Ursprünglich als nicht näher bestimmbarer Sphenodontier klassifiziert, wurde er 2018 als basaler Stammgruppen-Archosaurier unter dem Namen Colobops noviportensis neu beschrieben,[34] woraufhin die in dieser Arbeit getroffenen Interpretationen und die daraus gezogenen Schlüsse umgehend mit Verweis auf den Erhaltungszustand des Materials angezweifelt wurden.[35]
.jpg.webp)
Fossilführende, relativ klar als solche identifizierbare Sumpfablagerungen (definiert als dysoxische feinkörnige Ablagerungen in dicht mit Pflanzen bestandenen Teilen von Stillgewässern) sind auf die südlichen Becken der Newark-Supergruppe und auf die untere Obertrias (Karnium) beschränkt.[22] So kommt im unteren Karnium (Doswell-Formation) des kleinen Richmond-Beckens (Virginia) die älteste gut erhaltene Makroflora der Abfolge vor.[22] Sie wird von Farnen (u. a. Clathropteris), Schachtelhalmen (u. a. Neocalamites, Equisetites) und Palmfarnen (Macrotaeniopteris, Taeniopteris, Pterophyllum) dominiert. Mit ihr assoziiert ist eine Fischfauna, in der die Art Dictyopyge macrurus (Palaeonisciformes, Redfieldioidei) die bei weitem häufigste Spezies stellt. Diese Art ist endemisch in den kleinen Becken im Osten Virginias und dort „omnipräsent“ in allen fischführenden Schichten, nicht nur im unteren Karn. Darin unterscheiden sich die Fischfaunen dieser Becken von allen anderen obertriassischen Fischfaunen in der Newark-Supergruppe.[36] Jüngere (aber teils unsicher datierte) karnische Landwirbeltierfaunen in (mutmaßlichen) Sumpfablagerungen dieser Becken enthalten nicht-mammale Cynodontier (so den Traversodontiden Boreogomphodon jeffersoni[37], den Eucynodontier Microconodon tenuirostris und den Cynodontier unklarer Stellung Dromatherium sylvestre[38]), Dicynodontier (Placerias), „höhere“ Krokodil-Stammgruppenvertreter (Carnufex carolinensis[39] und den Sphenosuchier Dromicosuchus grallator), Phytosaurier (Rutiodon, dominant im Mittel-Karn), den nur von seinen Giftzähnen her bekannten Stammgruppen-Archosaurier Uatchitodon[40] sowie traditionell als Metoposauriden klassifizierte Temnospondylen.[22] Der aus möglichen Sumpfablagerungen des Newark-Beckens stammende, ebenfalls traditionell als Metoposauroide eingestufte Temnospondyle Calamops paludosus wird mittlerweile den Trematosauroiden zugerechnet.[41]
Aus einer flach-offenlakustrinen Fazies der Doswell-Formation des Taylorsville-Beckens (Virginia) wurde der krokodilähnliche, aber im Detail sehr eigentümlich gebaute basale Stammgruppen-Archosaurier Doswellia kaltenbachi beschrieben.[22]
Der weit überwiegende Teil der besser erhaltenen Fossilexemplare entstammt tief-lakustrin abgelagerten Schichten in den Van-Houten-Zyklen der zentraleren Beckenfazies (siehe oben). Reichhaltige allochthone Makrofloren mit Farnen, Schachtelhalmen, Palmfarnen und Koniferen kommen darin aber nicht generell vor, sondern vermutlich nur dort, wo die Ufer relativ nahe waren und eingetragenes Pflanzenmaterial mithin die tieferen Seeregionen leicht erreichen konnte, d. h. nur in den schmalen der größeren Becken wie dem Dan River-Danville-Becken. Ähnliches gilt anscheinend auch für das Vorkommen von Insekten. Im Ober-Karn oder Unter-Nor (Cow-Branch- bzw. Lockatong-Formation) des Danville-Beckens ist mit dem „Solite Quarry“ nahe Eden („Leaksville Junction“, North Carolina) eine einzelne obertriassische Fundstelle bekannt, in der vollständig und detailliert erhaltene aquatische und terrestrische Insekten häufig sind.[22][42] Bis 2007 waren in dieser Lokalität ca. 50 verschiedene Arten von Fluginsekten identifiziert worden, die sich auf 11 Großgruppen (Schaben, Käfer, Zweiflügler, Schnabelfliegen, Libellen, Heuschrecken, Steinfliegen, Pflanzenläuse, Zikaden, Wanzen und Fransenflügler) verteilen.[43] Bis 2010 waren während der Grabungen insgesamt 4000 Exemplare aufgesammelt worden.[42] Daneben ist aus dieser Lokalität die geologisch älteste Echte Webspinne beschrieben worden.[44]
Aus den höheren trophischen Niveaus sind in der Tiefwasserfazies in erster Linie Fische überliefert. Insgesamt sind zehntausende Exemplare in verschiedenen Grabungsstätten aufgesammelt worden.[22] Allgemein dominieren Vertreter der Typusgattung der Semionotiformes Semionotus, in der höheren Obertrias dominieren zudem entweder der Palaeoniscoide Turseodus oder der Quastenflosser Diplurus (= Osteopleurus) newarki.[22] Seltener aber immer noch relativ häufig sind Landwirbeltiere, vor allem vertreten durch die eigentümlichen Diapsiden Tanytrachelos ahynis (ein Tanystropheide) und Hypuronector limnaios[45] (ein Drepanosauride mit umstrittener Ökologie) sowie das diapside Gleitreptil Icarosaurus siefkeri und einige Überreste von Phytosauriern.[22] Eine bedeutende Fundstelle all dieser Reptilien ist der mittlerweile nicht mehr existente „Granton Quarry“ im Township North Bergen, New Jersey (Lockatong-Formation, Newark-Becken).[45][46]
Unterjura

Die einzige ergiebige unterjurassische Landwirbeltier-Fossillagerstätte im östlichen Nordamerika[22] ist Wasson Bluff bei Parrsboro im Fundy-Becken. In den dortigen feinsandigen bis brekziösen/konglomeratischen Beckenrandablagerungen und siltig-tonigen distaleren Äquivalenten (McCoy-Brook-Subformation der Shuttle-Meadow-Formation, unterstes Hettangium) dominieren Sphenodontier (Clevosaurus bairdi) und Protosuchier (Protosuchus micmac) sowie Semionotus und cf. Hybodus. Dinosaurier bilden nur ein untergeordnetes Element der Fauna und sind in erster Linie durch einen mutmaßlichen „Prosauropoden“ (cf. Ammosaurus bzw. „Fendusaurus eldoni “ vgl. [47]) vertreten. Ferner kommen spärliche Reste von kleinen Theropoden und Ornithischiern vor.[22][29][16] Amnioten der Synapsidenlinie sind durch nicht-mammale Cynodontier aus den Gruppen Tritheledontidae (Pachygenelus cf. monus) und Tritylodontidae (Oligokyphus) repräsentiert.[16]
Die 1818 gemachten Erstfunde von „Prosauropoden“-Resten (Anchisaurus polyzelus) in alluvialen/fluviatilen Sandsteinen des „Connecticut Valley“ (Portland-Gruppe des Hartford-Deerfield-Beckens) waren die historisch ersten, wenngleich, mehr als 30 Jahre vor der Errichtung des Taxons Dinosauria durch Richard Owen, zunächst missgedeuteten Funde von Dinosaurierknochen in Nordamerika.[48][49] Aus in etwa den gleichen Schichten und der gleichen Gegend stammen die Überreste eines coelophysoiden Theropoden (Podokesaurus holyokensis, Originalmaterial 1916 in einem Feuer vernichtet).[22][50]
Aus tonig-siltigen Schichten des Newark- sowie Hartford-Deerfield-Beckens, die dem sedimentologischen Befund nach in Schwemmebenen oder flachlakustrin abgelagert wurden, sind Überreste von Insekten bekannt, die überwiegend von Käfern stammen (meist isolierte Elytren). In zwei Lokalitäten im Turners-Falls-Sandstein (East-Berlin-Formation nach neuer Nomenklatur; frühes Hettang) des Deerfield-(Teil-)Beckens nahe Turners Falls in Massachusetts kommen in großer Dichte mutmaßliche Käferlarven vor, die 1850 die ersten mesozoischen Insektenfossilien waren, die in Nordamerika nachweislich aufgesammelt wurden. Sie wurden 1858 unter dem Namen Mormolucoides articulatus erstbeschrieben. Diese Larven starben seinerzeit vermutlich massenhaft während der Trockenzeit durch die zunehmende Versalzung der Flachwasserbereiche.[51]

Aus der tief-lakustrinen Fazies der unterjurassischen Schichten der Newark-Supergruppe wurden zehntausende Exemplare mehr oder weniger vollständiger Fische geborgen. Dafür enthält sie mit Ausnahme einiger weniger isolierter Zähne und Einzelknochen, die wahrscheinlich von Dinosauriern stammen, sogut wie keine Überreste von Landwirbeltieren. Die Fischfaunen werden von verschiedenen endemischen Arten von Semionotus dominiert und umfassen ferner Vertreter der Gattungen Redfieldius (Typusgattung der Palaeonisciformes-Subgruppe Redfieldioidei), Ptycholepis (Typusgattung der basalen Strahlenflosser-Gruppe Ptycholepidiformes) und den Quastenflosser Diplurus longicaudatus.[22] Auch diese Lagerstätten haben einen hohen wissenschaftshistorischen Wert, denn ihnen entstammen die gesichert historisch ältesten Funde artikulierter fossiler Fische in den USA.[49] Die in der jurassischen Tiefwasserfazies überlieferten Pflanzen sind durch isolierte aber gut erhaltene Triebe, Blätter und Zweige von Farnen, Palmfarnen (Otozamites) und Koniferen vertreten.[22]
Spurenfossilien
Die Datierung der Honeycomb-Point-Formation als oberpermisch basiert zwar auch auf palichnologischen Befunden, die betreffenden Trittsiegel, die Ähnlichkeit mit den permischen Formen Dimetropus und Hyloidichnus aufweisen, sind jedoch für eine genaue Bestimmung zu schlecht erhalten und überdies nicht unmittelbar zugänglich.[16]
Die fluviatilen und insbesondere die randlich-lakustrin abgelagerten obertriassischen und unterjurassischen Schichten der Newark-Supergruppe im Nordosten der USA sind schon seit dem 19. Jahrhundert weltbekannt für ihre Tetrapodenfährten und -trittsiegel. Hinsichtlich der Häufigkeitsverteilung der durch die Trittsiegel mutmaßlich repräsentierten Landwirbeltier-Taxa unterscheidet sich die Spurenfossilüberlieferung zum Teil deutlich von der Körperfossilüberlieferung.
So sind beispielsweise in der Obertrias die Spurengattungen Atreipus, Grallator, Anchisauripus und Evazoum, die basaleren Dinosauropmorpha (z. B. Silesauriden) und „echten“ Dinosauriern als Verursachern zugeordnet werden, gegenüber dem von Dinosauromorpha überlieferten Knochen- und Zahnmaterial stark überrepräsentiert. Die Spurengattungen Rhynchosauroides (Sphenodotiertrittsiegel/-fährten), Gwyneddichnium (Tanystropheidentrittsiegel/-fährten) sowie Procolophonichnus und Cynodontipus (Trittsiegel/-fährten bzw. Baue von Procolophonoiden) hingegen sind eher unterrepräsentiert. Temnospondylenspuren sind in der Trittsiegelüberlieferung überhaupt nicht vertreten. Die Häufigkeit der Phytosaurierspur Apatopus stimmt ungefähr mit der Häufigkeit überlieferter Phytosaurierknochen überein.[16][22]

Im Jura besteht ein ähnliches Ungleichgewicht. Dort sind Theropoden- und Ornithischierspuren (Grallator, Eubrontes, Amonoepus) stark überrepräsentiert, wohingegen die überwiegend als „Prosauropoden“-Spur gedeutete Ichnogattung Otozoum unterrepräsentiert ist. Ebenfalls unterrepräsentiert sind die seltenen Spurengattungen Rhynchosauroides und Ameghinichnus von denen letztgenannte tritheledontiden Cynodontiern zugeordnet wird. Wieder ist nur eine Krokodillinien-Archosaurierspur, Batrachopus, ungefähr genau so häufig wie das Knochenmaterial ihrer potenziellen Verursacher.[16][22]
Das Ungleichgewicht wird damit erklärt, dass zum einen Spuren- und Körperfossilgesellschaften jeweils verschiedene Tetrapodenfaunen repräsentieren und dass zudem die Körperfossilgesellschaften keine echten Faunen repräsentieren, sondern wahrscheinlich durch Transport selektiert und gemischt wurden.[22]
Eine sehr häufige Wirbellosengrabspur ist Scoyenia.[22]
Weblinks
- Early Mesozoic tetrapods of the Newark Supergroup of eastern North America – Seite von Hans-Dieter Sues auf der Unterpräsenz des Evolution of Terrestrial Ecosystems Program (ETE) der Internetpräsenz des Smithsonian National Museum of Natural History (USNM)
Einzelnachweise
- Gwendolyn W. Luttrell: Stratigraphic nomenclature of the Newark Supergroup of eastern North America. U.S. Geological Survey Bulletin 1572. Washington (DC) 1989, online.
- Robert E. Weems, Lawrence H. Tanner, Spencer G. Lucas: Synthesis and revision of the lithostratigraphic groups and formations in the Upper Permian?–Lower Jurassic Newark Supergroup of eastern North America. Stratigraphy. Bd. 13, Nr. 2, 2016, S. 111–153, online (alternativer Volltextzugriff: ResearchGate).
- William Charles Redfield: On the relations of the fossil fishes of the sandstone of Connecticut and other Atlantic states to the Liassic and Oolithic periods. American Journal of Science and Arts, 2nd series. Bd. 22, Nr. 66, 1856, S. 357–363, BHL.
- Israel C. Russel: Correlation papers: The Newark system. U.S. Geological Survey Bulletin 85. Washington (DC) 1892, online.
- James D. Dana: Manual of Geology – Treating of the Principles of the Science, with Special Reference to American Geological History. 2. Auflage. Ivison, Blakeman, Taylor & Co., New York 1875, archive.org, S. 403 ff..
- Elizabeth M. Truswell: Pre-Cenozoic palynolgy and continental movements. S. 13–26 in: M. W. McElhinny, D. A. Valencio (Hrsg.): Paleoreconstruction of the Continents. Geodynamic Series, Volume 2. American Geophysical Union/Geological Society of America, Washington (DC)/Boulder (CO) 1981, ISBN 0-87590-511-0, S. 19.
- Paul E. Olsen: On the use of the term Newark for Triassic and Early Jurassic rocks of eastern North America. Newsletters on Stratigraphy. Bd. 7, Nr. 2, 1978, S. 90–95, doi:10.1127/nos/7/1978/90.
- Albert J. Froelich, Paul E. Olsen: Newark Supergroup, a revision of the Newark Group of eastern North America. S. A55–A58 in: Stratigraphic Notes, 1983. U.S. Geological Survey Bulletin 1537-A. Washington (DC) 1984, online.
- Martha Oliver Withjack, Roy W. Schlische: A Review of Tectonic Events on the Passive Margin of Eastern North America. S. 203–235 in: Paul Post, Donald Olson, Kevin Lyons, Stephen Palmes, Peter Harrison, Norman Rosen (Hrsg.): 25th Annual Bob F. Perkins Research Conference: Petroleum Systems of Divergent Continental Margin Basins. Society for Sedimentary Geology (SEPM), Gulf Coast Section, Houston (TX) 2005, ResearchGate.
- Robert E. Weems, Paul E. Olsen: Synthesis and revision of groups within the Newark Supergroup, eastern North America. Geological Society of America Bulletin. Bd. 109, Nr. 2, 1997, S. 195–209, doi:10.1130/0016-7606(1997)109<0195:SAROGW>2.3.CO;2 (alternativer Volltextzugriff: ResearchGate).
- Warren Manspeizer, Jelle DeBoer, John K. Costain, Albert J. Froelich, Cahit Çoruh, Paul E. Olsen, Gregory J. McHone, John H. Puffer, David C. Prowell: Post-Paleozoic activity. S. 319–374 in: Robert D. Hatcher, Jr., William A. Thomas, George W. Viele (Hrsg.): The Geology of North America, Vol F-2: The Appalachian-Ouachita Orogen in the United States. Geological Society of America, Boulder (CO) 1989, ISBN 0-8137-5209-4.
- Paul E. Olsen: Fossil Great Lakes of the Newark Supergroup – 30 Years Later. S. 101–162 in: Alan I. Benimoff (Hrsg.): Field Trip Guidebook, New York State Geological Association, 82nd Annual Meeting, Staten Island, NY, September 24-26, 2010. ResearchGate.
- Paul E. Olsen: Stratigraphic record of the early Mesozoic breakup of Pangea in the Laurasia-Gondwana Rift System. Annual Review of Earth and Planetary Sciences. Bd. 25, Nr. 1, 1997, S. 337–401, doi:10.1146/annurev.earth.25.1.337 (alternativer Volltextzugriff: ResearchGate).
- Andrea Marzoli, Sara Callegaro, Jacopo Dal Corso, Joshua H. F. L. Davies, Massimo Chiaradia, Nassrrdine Youbi, Hervé Bertrand, Laurie Reisberg, Renaud Merle, Fred Jourdan: The Central Atlantic Magmatic Province (CAMP): A Review. S. 91–125 in: Lawrence H. Tanner (Hrsg.): The Late Triassic World – Earth in a Time of Transition. Topics in Geobiology, Bd. 46. Springer 2018, ISBN 978-3-319-68008-8.
- Newark Supergroup Correlation Chart. Tafel 1 (Extra-Beilage) in: Albert J. Froelich, Gilpin R. Robinson, Jr. (Hrsg.): Studies of the Early Mesozoic Basins of the Eastern United States. U.S. Geological Survey Bulletin 1776. Washington (DC) 1988, online.
- Hans-Dieter Sues, Paul E. Olsen: Stratigraphic and temporal context and faunal diversity of Permian-Jurassic continental tetrapod assemblages from the Fundy rift basin, eastern Canada. Atlantic Geology. Bd. 51, Nr. 1, 2015, S. 139–205, doi:10.4138/atlgeol.2015.006 (Open Access).
- Paul E. Olsen, Dennis V. Kent, Sarah J. Fowell, Roy W. Schlische, Martha O. Withjack, Peter M. LeTourneau: Implications of a comparison of the stratigraphy and depositional environments of the Argana (Morocco) and Fundy (Nova Scotia, Canada) Permian-Jurassic basins. S. 165–183 in: M. Oujidi, M. Et-Touhamin (Hrsg.): Le Permien et le Trias du Maroc. Actes de la Premièr Réunion su Groupe Marocain du Permien et du Trias. Hilal Impression, Oujda 2000, doi:10.7916/D86Q26MF.
- Heinz W. Kozur, Robert E. Weems: The biostratigraphic importance of conchostracans in the continental Triassic of the northern hemisphere. S. 315–417 in Spencer G. Lucas (Hrsg.): The Triassic Timescale. Geological Society, London, Special Publications, 334. 2010, doi:10.1144/SP334.1, S. 341.
- Nicholas G. McDonald, Peter M. LeTourneau: Paleoenvironmental reconstruction of a fluvial-deltaic-lacustrine sequence, Lower Jurassic Portland Formation, Suffield, Connecticut. S. 24–30 in: Albert J. Froelich, Gilpin R. Robinson, Jr. (Hrsg.): Studies of the Early Mesozoic Basins of the Eastern United States. U.S. Geological Survey Bulletin 1776. Washington (DC) 1988, online, S. 24.
- Paul E. Olsen, Dennis V. Kent, Bruce Cornet, William K. Witte, Roy W. Schlische: High-resolution stratigraphy of the Newark rift basin (early Mesozoic, eastern North America). Geological Society of America Bulletin. Bd. 108, Nr. 1, 1996, S. 40–77, doi:10.1130/0016-7606(1996)108<0040:HRSOTN>2.3.CO;2 (alternativer Volltextzugriff: SemanticScholar).
- Paul E. Olsen, Dennis V. Kent: Long-period Milankovitch cycles from the Late Triassic and Early Jurassic of eastern North America and their implications for the calibration of the Early Mesozoic time-scale and the long-term behaviour of the planets. Philosophical Transactions of the Royal Society of London, A. Bd. 357, Nr. 1757 (Discussion Meeting Issue ‘Astronomical (Milankovitch) calibration of the geological time–scale’), 1999, S. 1761–1786, doi:10.1098/rsta.1999.0400 (alternativer Volltextzugriff: ResearchGate).
- Paul E. Olsen: Paleontology and paleoecology of the Newark Supergroup (early Mesozoic, eastern North America). S. 185–230 in: Warren Manspeizer (Hrsg.): Triassic-Jurassic Rifting: Continental Breakup and the Origin of the Atlantic Ocean and Passive Margins. Part A. Developments in Geotectonics 22. Elsevier, Amsterdam/New York 1988, ISBN 0-444-42903-4.
- Eleanora I. Robbins, Gerald P. Wilkes, Daniel A. Textoris: Coal deposits of the Newark rift system. S. 649–682 in: Warren Manspeizer (Hrsg.): Triassic-Jurassic Rifting: Continental Breakup and the Origin of the Atlantic Ocean and Passive Margins. Developments in Geotectonics 22. Elsevier, Amsterdam/New York 1988, ISBN 0-444-42903-4.
- Peter W. Weigand, Paul C. Ragland: Geochemistry of Mesozoic dolerite dikes from eastern North America. Contributions to Mineralogy and Petrology. Bd. 29, Nr. 3, 1970, S. 195–214, doi:10.1007/BF00373305.
- Richard P. Tollo, David Gottfried: Early Jurassic quartz normative magmatism of the eastern North America province: Evidence for independent magmas and distinct sources. In: Continental Magmatism, Abstracts – International Association of Volcanology and Chemistry of the Earth's Interior, General Assembly, Santa Fe, New Mexico, USA, June 25 – July 1 1989. New Mexico Bureau of Mines and Mineral Resources Bulletin 131. Socorro (NM) 1989, S. 270, online (kompletter Band als PDF herunterladbar), HTML-Version (nur Abstract).
- Richard P. Tollo, David Gottfried: Petrochemistry of Jurassic basalt from eight cores, Newark Basin, New Jersey: Implications for the volcanic petrogenesis of the Newark Supergroup. S. 233–259 in: John H. Puffer, Paul C. Ragland (Hrsg.): Eastern North American Mesozoic Magmatism. Geological Society of America Special Paper 268. Boulder (CO) 1992, doi:10.1130/SPE268-p233; für eine im Zusammenhang mit dieser Arbeit zuvor publizierte Kurzbeschreibung der Geologie der Basalte des Newark-Beckens und die Ergebnisse der gesteinschemischen Analysen siehe Richard P. Tollo, David P. Hawkins, David Gottfried: Petrographic and geochemical data for Jurassic basalt from eight cores, Newark basin, New Jersey. Open-File Report 90-689. U.S. Geological Survey, U.S. Department of the Interior, Washington (DC) 1990, online.
- Harvey E. Belkin, Daniel F. Fiorito: Geochemical data for Jurassic basalts in the Early Mesozoic Newark Basin, New Jersey: the data of G.T. Faust. Open-File Report 93-10. U.S. Geological Survey, U.S. Department of the Interior, Washington (DC) 1993, online.
- Paul E. Olsen, Emma C. Rainforth: The “Age of Dinosaurs” in the Newark Basin, with Special Reference to the Lower Hudson Valley. S. 59–176 in: Alexander E. Gates (Hrsg.): 2001 New York State Geological Association Fieldtrip Guidebook: Geology of the Lower Hudson Valley. PDF (10,9 MB), S. 61.
- Paul E. Olsen, Jessica H. Whiteside, Timothy Fedak: Triassic-Jurassic faunal and floral transition in the Fundy Basin, Nova Scotia. North American Paleontological Convention (NAPC) 2005, Halifax, Nova Scotia. Exkursionsführer, PDF (46,7 MB), S. 32.
- Phillip Huber, Spencer G. Lucas, Adrian P. Hunt: Vertebrate biochronology of the Newark Supergroup Triassic, eastern North America. S. 179–186 in: Spencer G. Lucas, Michael Morales (Hrsg.): The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin, 3. Albuquerque 1993 (online), S. 180.
- Spencer G. Lucas: Late Triassic Terrestrial Tetrapods: Biostratigraphy, Biochronology and Biotic Events. S. 351–405 in: L.H. Tanner (Hrsg.): The Late Triassic World. Topics in Geobiology, 46. Springer, 2018, ISBN
- Sterling J. Nesbitt, Randall B. Irmis, William G. Parker: A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology. Bd. 5, Nr. 2, 2007, S. 209–243, doi:10.1017/S1477201907002040, S. 230.
- Paul E. Olsen, Dennis V. Kent, Jessica H. Whiteside: Implications of the Newark Supergroup-based astrochronology and geomagnetic polarity time scale (Newark-APTS) for the tempo and mode of the early diversification of the Dinosauria. Earth and Environmental Science Transactions of the Royal Society of Edinburgh. Bd. 101, Spezial-Nr. 3–4 (Late Triassic Terrestrial Biotas and the Rise of Dinosaurs), 2011, S. 201–229, doi:10.1017/S1755691011020032, S. 221 f.
- Adam C. Pritchard, Jacques A. Gauthier, Michael Hanson, Gabriel S. Bever, Bhart-Anjan S. Bhullar: A tiny Triassic saurian from Connecticut and the early evolution of the diapsid feeding apparatus. Nature Communications. Bd. 9, 2018, Art.-Nr. 1213, doi:10.1038/s41467-018-03508-1 (Open Access)
- Torsten M. Scheyer, Stephan N. F. Spiekman, Hans-Dieter Sues, Martín D. Ezcurra, Richard J. Butler, Marc E. H. Jones: Colobops: a juvenile rhynchocephalian reptile (Lepidosauromorpha), not a diminutive archosauromorph with an unusually strong bite. Royal Society Open Science. Bd. 7, 2020, Art.-Nr. 192179, doi:10.1098/rsos.192179 (Open Access)
- Bruce Cornet, Paul E. Olsen: Early to Middle Carnian (Triassic) Flora and Fauna of the Richmond and Taylorsville Basins, Virginia and Maryland, U.S.A. Virginia Museum of Natural History Guidebook, Nr. 1. Martinsville (VA), 1990.
- Hans-Dieter Sues, Paul E. Olsen, Peter A. Kroehler: Small tetrapods from the Upper Triassic of the Richmond basin (Newark Supergroup), Virginia. S. 161–170 in: Nicholas C. Fraser, Hans-Dieter Sues (Hrsg.): In the shadow of the dinosaurs: Early Mesozoic tetrapods. Cambridge University Press, 1994, ISBN 0-521-45242-2.
- Hans-Dieter Sues: On Microconodon, a Late Triassic cynodont from the Newark Supergroup of eastern North America. Bulletin of the Museum of Comparative Zoology. Bd. 156, Nr. 1, 2001, S. 37–48, BHL
- Lindsay E. Zanno, Susan Drymala, Sterling J. Nesbitt, Vincent P. Schneider: Early crocodylomorph increases top tier predator diversity during rise of dinosaurs. Scientific Reports. Bd. 5, 2015, Art.-Nr. 9276, doi:10.1038/srep09276 (Open Access)
- Jonathan S. Mitchell, Andrew B. Heckert, Hans-Dieter Sues: Grooves to tubes: evolution of the venom delivery system in a Late Triassic “reptile”. Naturwissenschaften. Bd. 97, 2010, S. 1117–1121, doi:10.1007/s00114-010-0729-0 (Open Access)
- Hans-Dieter Sues, Rainer R. Schoch: Anatomy and phylogenetic relationships of Calamops paludosus (Temnospondyli, Stereospondyli) from the Triassic of the Newark Basin, Pennsylvania. Journal of Vertebrate Paleontology. Bd. 33, Nr. 5, 2013, S. 1061–1070, doi:10.1080/02724634.2013.759120 (alternativer Volltextzugriff: ResearchGate).
- Cynthia M. Liutkus, James S. Beard, Nicholas C. Fraser, P. C. Ragland: Use of fine-scale stratigraphy and chemostratigraphy to evaluate conditions of deposition and preservation of a Triassic Lagerstätte, south-central Virginia. Journal of Paleolimnology. Bd. 44, 2010, S. 645–666, doi:10.1007/s10933-010-9445-1 (Open Access).
- Vladimir A. Blagoderov, David A. Grimaldi, Nicholas C. Fraser: How time flies for flies: diverse Diptera from the Triassic of Virginia and early radiation of the order. American Museum Novitates, 3572. American Museum of Natural History, New York 2007, online, S. 5.
- Paul A. Selden, John M. Anderson, Heidi M. Anderson, Nicholas C. Fraser: Fossil araneomorph spiders from the Triassic of South Africa and Virginia. The Journal of Arachnology. Bd. 27, Nr. 2, 1999, S. 401–414, JSTOR.
- Edwin H. Colbert, Paul E. Olsen: A new and unusual aquatic reptile from the Lockatong Formation of New Jersey (Late Triassic, Newark Supergroup). American Museum Novitates, 3334. American Museum of Natural History, New York 2001, online.
- Edwin H. Colbert: A phytosaur from North Bergen, New Jersey. American Museum Novitates, 2230. American Museum of Natural History, New York 1965, online.
- Adam Douglas Marsh: The Osteology of Sarahsaurus aurifontanalis and Geochemical Observations of the Dinosaurs from the Type Quarry of Sarahsaurus (Kayenta Formation), Coconino County, Arizona. Ms-Thesis, The University of Texas at Austin, 2013, online, S. 18.
- Peter Malcolm Galton: Prosauropod dinosaurs (Reptilia: Saurischia) of North America. Postilla, 169. Peabody Museum of Natural History, New Haven 1976, BHL, S. 4
- Arnaud Brignon: The earliest discoveries of articulated fossil fishes (Actinopterygii) in the United States: A historical perspective. American Journal of Science. Bd. 317, Nr. 2, 2017, S. 216–250, doi:10.2475/02.2017.03.
- David B. Weishampel: Another look at the dinosaurs of the East Coast of North America. S. 129–168 in: Colectivo Arqueológico-Paleontológico Salense (Hrsg.): Actas de las III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes, Burgos 2006, Yumpu, S. 136 f.
- Phillip Huber, Nicholas G. McDonald, Paul E. Olsen: Early Jurassic Insects from the Newark Supergroup, Northeastern United States. S. 206–223 in: Peter M. LeTourneau, Paul E. Olsen (Hrsg.): The Great Rift Valleys of Pangea in Eastern North America, Volume 2: Sedimentology, Stratigraphy, and Paleontology. Columbia University Press, New York 2003, ISBN 0-231-12676-X, ResearchGate.