Fischschädel
Der Schädel der Fische ist der älteste in der Stammesgeschichte der Chordatiere (Chordata).

Bauteile

Die Evolution des Fischschädels lässt sich – wie der Schädel der übrigen, stammesgeschichtlich jüngeren Schädel- oder Wirbeltiere – auf drei funktionell unterschiedliche Bauteile zurückführen.
Neurocranium
Das Neurocranium („Gehirnschädel“) umschließt schützend (ähnlich wie bei den Cephalopoden) das Nervenzentrum (Gehirn) und, so weit dies möglich ist, die hier angeordneten großen Sinnesorgane. Wie das Lanzettfischchen (Branchiostoma) zeigt, ist für eine Schwimm- oder Wühlform ohne diese Sinnesorgane noch kein Schädel nötig (die Chorda dorsalis als Versteifung bis in die Körperspitze genügt). Sobald die Sinnesorgane (Nase, Augen, Vestibularorgan) mit dem Gehirn dazwischen aber vorhanden sind, wird das Neurocranium zum Axocranium, dem festen Bug eines aktiven Schwimmtier-Körpers. Bei fossilen „Knochenfischen“ (Palaeonisciformes, aber auch schon lange vorher) besteht das Neurocranium aus einem einzigen Stück (wenn auch mit Knorpeleinbuchtungen, wahrscheinlich gegen Bruchgefahr), bei rezenten Knochenfischen aber aus 10 bis 14 „Knorpel- oder Ersatzknochen“ (weil sie im embryonalen Knorpelschädel durch Knorpel-Ersatz entstehen); bei den Tetrapoden nimmt ihre Zahl langsam wieder ab.
Splanchnocranium

Da nun diese „Schädeltiere“ von Anfang an Schlundspalten hatten (primär wegen der seihenden Nahrungsaufnahme – vgl. Tunicata; die zunehmende Größe bringt allerdings bald die Atemfunktion mit sich: Kiemenbildung), werden ab einer gewissen Größe auch für diese Kiemenspalten Skelettstützen nötig: das Splanchnocranium (von gr. σπλάγχνα pl „Eingeweide“). Diese Atemfunktion ist nicht notwendig an das Vorderende gebunden (Cephalopoden, Krebse), erweist sich hier aber „nachträglich“ als vorteilhaft – das Branchiocranium wird (von den „Agnatha“ [vgl. Myxinoidea und Petromyzontida, bei denen es weit hinter das Neurocranium reicht, während das Dermatocranium völlig reduziert ist] über die Chondrichthyes hin zu den Osteichthyes – schon bei den Acanthodii) immer enger an das Axocranium gebunden (allerdings in der weiteren Entwicklungsgeschichte hin zu luftatmenden Landwirbeltieren wieder überflüssig und bis aufs Zungenbein und weitere „Reste“ reduziert). Das Branchio- oder Splanchnocranium (die Kiemen- bzw. Pharyngealbogen mit Hyoid- und Kieferbogen) besteht bei Knochenfischen aus ca. 33 benannten Knochen, die meisten paarig und aneinander (in Amphiarthrosen) gelenkend.
Dermatocranium

Später (?) kommen – wahrscheinlich als „Reaktion“ auf gepanzerte Arthropoden wie die ordovicischen Eurypterida („Seeskorpione“) als Feinde – am Körpervorderende außen (zum Schutz von Weichteilen wie Muskeln, Nerven, Haut-Sinneskanälen) (dutzende) kleine Knochenplatten der Haut hinzu: das Dermatocranium, das als Besonderheit sehr harte Oberflächenstrukturen mitbringt, die sich dann im Bereich der Mundöffnung zu den Zähnen differenzieren konnten. Deckknochen treten ja auch in der ektodermalen Mund- und Rachenhöhle (Pharynx), d. h. innen am Branchiocranium, auf (siehe auch Kiemenreuse). Man hat jahrzehntelang gestritten, ob die Kopfseitenlinien-Sinneskanäle oder die Zähne den „Ursprung“ der Deckknochen darstellen (beides muss nicht sein). Auch die Deckknochen und Zähne gehören übrigens zum Mesoderm – nur der Zahnschmelz dürfte eine ektodermale Leistung sein. Das „Kalk“-Skelett überhaupt lässt sich wohl mit den Sklerit-Platten (dem Stereom) der Echinodermata „in Verbindung bringen“.
Es ist heute kaum möglich, die Abfolge des Auftretens der drei Bauteile festzulegen, die ja durch ihr Gewicht verschiedene phylogenetische Entwicklungstendenzen vorgeben. So scheinen schon bei den kleinen Conodonta vom „Schädel“ (?) nur die Zähne als Hartteile am lebenden Tier erhalten – alles Übrige ist der nektischen Lebensweise geopfert. Bei den Thelodonti ist ein dünner Haut-(Schuppen)panzer vorhanden. Bei den Ostracodermi u. a. hingegen verhindert die starke Panzerung das Leben im freien Wasser. Die gepanzerten Placodermi hatten zwar schon Kiefer, aber noch keine Zähne geeigneter Größe. Wahrscheinlich hatten sie auch schon eine Schwimmblase, um trotz schwerem Knochengewebe als erste Großräuber das freie Wasser durchschweifen zu können (Dunkleosteus war 8 m lang). Der „Panzer“ bewirkt, dass das Endocranium (innen – also Neuro- und Splanchnocranium) nun knorplig (zart) bleiben kann. Da auch die Schwimmblase Nachteile mit sich bringt, verzichteten die „Knorpelfische“ wieder auf sie und auf einen Teil des Skeletts (Knochengewebe und Dermatocranium) – nicht aber auf die Zähne.
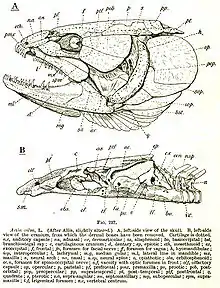
Syncranium
Die rezenten (jetzt lebenden) Knochenfische haben alle drei Schädel-Bauteile (und höchstens sekundär wenig Knochen, keine Zähne und keine Schwimmblase). Bei den Stören ist der Hautknochenpanzer noch recht selbständig, aber bei den höheren Formen (besonders Teleostei) kommt es zu einer Vereinheitlichung („Syncranium“). Auch nimmt die Anzahl der durch „Nähte“ (Suturen; über die Funktion von Nähten wissen wir sehr wenig), in Synarthrosen, miteinander verbundenen „Knochenindividuen“ ab (durch Verschmelzungen oder Ausfälle, wie man sagt; z. B. ist das Dentale der Teleosteer ein „Verschmelzungsprodukt“ aus Dentale, Mentomeckelianum, Spleniale, Surangulare (u. a.) bei Amia u. a., die aber auch nach Verkleinerung zugunsten des Dentale „ausgefallen“, d. h. verschwunden sein können).
Knochen des Fischschädels
Das Kopfskelett eines Stachelflossers, etwa des Barsches (Perca fluviatilis), besteht aus fast 160 Knochen (zum Vergleich: das des Menschen aus 31). Die meisten sind paarig (wir bezeichnen hier nur die unpaaren [u]). Im Schädeldach (also in Dorsal-Ansicht) sehen wir, von hinten nach vorne fortschreitend, folgende Deckknochen (d, mit Sinneskanal D) oder Knorpelknochen (a) bzw. Mischknochen (a+d verschmolzen): Supraoccipitale (u, a, vielleicht auch d; D), Supratemporale (D), d-Parietale, d-Epoticum, Intercalare (d), Frontale (D), Nasale (D) und (Mes-)Ethmoideum (u, a, d).
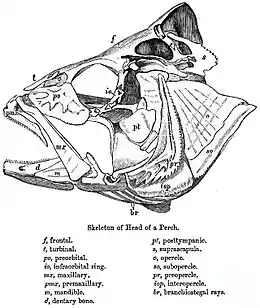

Die Seiten des Neurocranium machen folgende Verknöcherungen aus: a-Exoccipitale, Opisthoticum (D), a-Prooticum, a-Sphenoticum (mit d-Postfrontale vereint), a-Pteroticum (in den drei Otica liegen die drei Bogengänge des Vestibular-Organs), Basisphenoid (u), Pterosphenoid, (Orbitosphenoid, u, nur bei dickerer Augenhöhlenscheidewand), Ectetmoid (a mit d-Praefrontale). Das Auge wird geschützt durch Circumorbitalia (D; hier 5 oder 6 Suborbitalia – das erste heißt oft Praeorbitale oder Lacrimale). Zum Kiemendeckel (d) gehören Operculare, Suboperculare und Interoperculare. Die Ventralansicht zeigt die „Schädelbasis“: Basioccipitale (u), Parasphenoid (u, d) und Vomer (u, d).
Das Splanchnocranium besteht aus sieben „Visceralbögen“, deren Zurückführbarkeit auf „Kiemenbögen“ nicht für alle gesichert ist. Vier echte Kiemenbögen bestehen jeweils (von oben nach unten) aus Pharyngo-, Epi-, Cerato- und Hypobranchiale und je einem Basibranchiale (u); der „5. Bogen“ besteht nur aus dem „Cerato-Element“ (sagt man. Manche Elemente bleiben zeitlebens knorpelig). Diese Knorpelknochen (a; s. Pharyngealia) tragen innen Zahnplatten (d); a und d sind ganz verschieden weit verwachsen (das „5. Ceratobranchiale“ ist immer Mischknochen). Vor dem Kiemenkorb liegt ventral das Hyoid (daran angebändert die sieben d-Branchiostegal-Radien; s. Branchiostegalapparat), bestehend aus Epi-, Cerato- und zwei Hypohyale(n) und dem Basihyale (u, mit d-Glossohyale), dorsal das (a) Hyomandibulare, das außen vom Praeoperculare (D) bedeckt ist. Mit dem Quadratum ist das Hyoid durch das stäbchenförmige Inter- oder Stylohyale gelenkig verbunden, das Hyomandibulare durch das („eingekeilte“) Symplecticum. Der einstmalige „Kieferbogen“ (jetzt Suspensorium genannt) besteht aus: Palatinum (bei Percoidei nur d), zwei (d-) Pterygoiden (Ecto- und Ento-), Metapterygoid (a mit d), Quadratum (a, mit d?). Funktionell wichtig ist die meist breite Knorpelzone des Suspensoriums. Drei (bei Amia calva aber noch zehn!) Knochen: Dentale (D, mit a-Anteil), Articulare (a) und Angulare, bilden den Unterkiefer, in dem die elastische, knorpelige Mandibula stets erhalten ist. Der „Oberkiefer“ der Teleostei aber besteht nur mehr aus je zwei Deckknochen (ursprünglich der Nasenregion): Praemaxillare (bei Stachelflossern allein am Mundrand, bezahnt, mit (u) Rostralknorpel) und Maxillare (bei primitiveren Fischen noch bezahnt). Zuletzt zwei Sehnenverknöcherungen: das Urohyale (u, ventral am Hyoid) und das Sesam-Articulare (Ansatz der Kaumuskel-Sehne am Unterkiefer).

Beim Fisch kann man auch den Schultergürtel funktionell zum Schädel rechnen (z. B. da er die Kiemenhöhle hinten stützt), er besteht aus (d-) Supracleithrale, Cleithrale und den beiden a-Knochen Coracoideum und Scapulare; er gelenkt durch das Posttemporale am Hinterhaupt (vgl. die umgekehrte Semantik bei Dunkleosteus [Abb.]).
Funktionelles
(Hier nur ein sehr grober Überblick – die Vielfalt des Fischschädels ist unüberschaubar; vgl. Gregory 1933.) Der Schädel gelenkt an der Wirbelsäule, bei primitiveren Formen noch an der Chorda, die in ihn bis zum Basisphenoid hineinreichen kann. Die Gelenkung erfolgt am Basioccipitale, oft auch an den Exoccipitalia und ist meist wenig beweglich (Bugwirkung!). Größere Beweglichkeit ist gegeben z. B. durch Schädelhebung zum Zuschnappen (z. B. beim Zander; bei Uranoscopus, exzessiv bei manchen Tiefseefischen wie Malacosteinae); „Niederbeißen“ bei Clariidae; durch seitliche Kopfbewegungen berühmt geworden ist der Salamanderfisch Lepidogalaxias (von dem deshalb sogar angenommen wurde, dass er seine Augen nicht mehr bewegen kann. Wirbeltiere haben bewegliche Augen – dass es auch ohne geht, zeigen wieder die Cephalopoden). Zweck der Schädelhebung ist beim freischwimmenden Fisch, Fortbewegungs- und Schnapprichtung zur Deckung zu bringen (weil der Unterkiefer ja vorwiegend nach unten aufgeht). Eine andere Möglichkeit war es, nur den Vorderschädel zu heben, so dass das koordinierende Vestibularorgan (im „Otoccipitalblock“) in Ruhe bleiben konnte (intracraniale Kinetik, nur bei Sarcopterygii, rezent Latimeria; Nachteil: Bugwirkung geschwächt!). Es gab auch Fische, die nur die Nasenkapsel heben konnten (rezent bei Lepidosiren und Cobitidae, aber noch unklar). Suspensorium und Maxillarapparat (s. Fischmaul) höherer Fisch-Ordnungen schaffen hier ebenfalls Abhilfe.
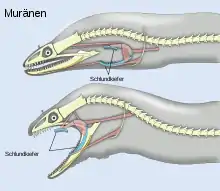
Abgesehen von der Rumpfmuskulatur setzen am Schädel (oder Teilen) ca. 60 Muskeln an (die meisten natürlich wieder paarig); einige davon in Portionen unterteilt, z. B. der Kaumuskel in bis zu 12. Das Suspensorium ermöglicht die laterale Erweiterung des Pharyngealraumes (für Atmung, Saugschnappen und (Großbrocken-)Schlucken); dazu gelenkt das Hyomandibulare am Pro-, Sphen- und Pteroticum und das Palatinum am Mes- und Ectethmoid (bei Karpfen und Welsen aber das Ectopterygoid). Die Bewegungen des Suspensorium nehmen oft das Lacrimale mit und dienen so der Ventilation des Geruchsorgans. Der am Quadratum gelenkende Unterkiefer (Sattelgelenk! Diarthrose) muss die Quererweiterung in der Symphyse mitmachen und dreht sich dabei der Länge nach mit den Zähnen nach außen. Lange Zähne können dabei auch umklappbar sein (sie richten sich dank elastischen Ligamenten nach der Beutepassage wieder auf; z. B. bei Merlucciidae). Es gibt sogar Fische mit intramandibularem Gelenk: zur Erhöhung der Beißkraft (Scaridae; „Streptognathie“). Die Bewegungen von Hyoid und Kiemendeckel sind für Atmung und Nahrungserwerb gleichermaßen wichtig; z. B. dient Operkel-Hebung bei den meisten Fischen, über das Interoperculum, zur Maulöffnung. Der Kiemenkorb ist mittels des Pharyngobranchiale I an der Schädelbasis (Parasphenoid) „aufgehängt“, um bei den Formen mit Retractor-Muskeln die zügigen Schlingbewegungen der dorsalen Pharyngealia (mit greifenden Zahnplatten) zu ermöglichen. Diese Pharyngealia sind aneinander gelenkende Pharyngo- und Epibranchalia (II–IV), die mit den erwähnten „Ceratobranchialia V“ zusammenwirken. Bei Molluskenfressern u. Ä. (Sparidae u. a.) kommt es sogar zur Ausbildung eines richtigen Kugelgelenks zwischen Pharyngealapparat und Parasphenoid. Bei den Scaridae gelenken die verschmolzenen „Ceratobranchialia V“ zwecks erhöhter Stabilität innen an den Cleithralia.
Etliche funktionelle Errungenschaften des Fischschädels vererbten sich von den Sarcopterygiern auf die Tetrapoden bis zu den Vögeln. Erst bei den Säugern ist das Meiste nicht mehr vorteilhaft und wieder aufgegeben (akinetischer Schädel – ähnlich schon bei den Dipnoi).
Siehe auch
Literatur
- William K. Gregory: Fish skulls. A study of the evolution of natural mechanisms. New York 1933, OCLC 251016995. (Ausgabe 1959 auf archive.org)
- Wilhelm Harder: Kopfregion und Schultergürtel der Fische. Schweizerbart, Stuttgart/ Berlin 1965.
- Wilhelm Marinelli, Anneliese Strenger: Vergleichende Anatomie und Morphologie der Wirbeltiere I-IV. Deuticke, Wien 1953–1973.
- Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbeltiere. Gustav Fischer, Stuttgart/ Jena/ New York 2004.
Weblinks
- Kompaktlexikon der Biologie, Spektrum Akademischer Verlag Schädel, Cranium