Kurzgesichtmaulwurf
Der Kurzgesichtmaulwurf (Scaptochirus moschatus) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Er kommt im nordöstlichen Teil Chinas vor und bewohnt dort trockene, offene Landschaften bestehend aus Grasflächen bis hin zu wüstenartigen Gebieten. Über die Lebensweise liegen kaum Informationen vor. Es handelt sich um einen mittelgroßen Vertreter der Maulwürfe. Wie alle Eigentlichen Maulwürfe zeichnet er sich durch einen walzenförmigen Körper mit kurzem Hals und grabschaufelartigen Vordergliedmaßen aus. Das Fell ist grau braun gefärbt, die Schnauze kurz gestaltet. Die Art wurde im Jahr 1867 wissenschaftlich eingeführt, gemeinsam mit der Gattung Scaptochirus, deren einzigen heute lebenden Angehörigen sie repräsentiert. Teilweise wurde ihr aber der im westlichen Eurasien auftretende Pater-David-Maulwurf zur Seite gestellt. Fossil trat Scaptochirus bereits im Pliozän in Erscheinung, aus dem Pleistozän wurden zwei ausgestorbene Arten beschrieben. In der stammesgeschichtlichen Vergangenheit kamen die Vertreter der Gattung auch deutlich weiter südlich in Ostasien vor. Der Bestand des Kurzgesichtmaulwurfs gilt als nicht gefährdet.
| Kurzgesichtmaulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Scaptochirus | ||||||||||||
| Milne-Edwards, 1867 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Scaptochirus moschatus | ||||||||||||
| Milne-Edwards, 1867 |
Merkmale
Habitus
Der Kurzgesichtmaulwurf ist ein mittelgroßer Vertreter der Maulwürfe. Seine Kopf-Rumpf-Länge variiert von 10,0 bis 12,6 cm. Der Schwanz wird 1,4 bis 2,3 cm lang. Angaben zum Gewicht liegen nicht vor. Ähnlich den anderen Angehörigen der Eigentlichen Maulwürfe (Talpini) weist er eine an die unterirdische Lebensweise angepasste Gestalt auf. Der Körper ist walzenförmig, der Hals kurz und die Vordergliedmaßen sind zu Grabwerkzeugen umgebildet. Die Hinterfüße messen zwischen 1,5 und 1,9 cm in der Länge. Das Fell ist deutlich grau braun gefärbt. Lediglich an der Schnauze und an den Vorderbeinen beziehungsweise auf der Unterseite kommen einzelne hellere Bereiche vor, die unter bestimmten Lichtverhältnissen teils silbrig schimmern. Die einzelnen Haare sind kurz. Sie haben schieferfarbene Basen und kurze bräunliche Spitzen. Am Maul selbst tritt nur ein dünner Haarflaum auf, ebenso auf der Rückseite der breiten Vorder- und der schmalen Hinterfüße. Der Schwanz ist schlank, kürzer als die Hinterfüße und nur spärlich mit Haaren bedeckt. Allerdings sitzt an der Spitze ein kleines Büschel an.[1][2][3]
Schädel- und Gebissmerkmale
Der Schädel ist 30,2 bis 35,7 mm lang und an den Jochbögen 13,3 bis 14,7 mm breit. An den Warzenfortsätzen wird der Hirnschädel 16,9 bis 18,1 mm breit. Insgesamt wirkt der Schädel vergleichsweise robust, allerdings ist das Rostrum kurz und breit. Seine Weite auf Höhe der Molaren beträgt 9,7 bis 11,1 mm. Die Paukenblasen sind breit und lang sowie flach. Der Unterkiefer ist massig, die drei Fortsätze (Kronenfortsatz, Gelenkfortsatz und Winkelfortsatz) heben sich durch ihren kräftigen Bau hervor. Im Gebiss sind 40 Zähne ausgebildet, die Zahnformel lautet: . Demnach ist gegenüber anderen Vertretern der Eigentlichen Maulwürfe die Anzahl der Prämolaren reduziert, was zu Kürzungen im Gesichtsbereich und somit zu der kurzen Schnauze führte. Es treten aber Variationen in der Zahnanzahl auf, da mitunter weitere Prämolaren reduziert sein können oder zusätzliche auftreten. Die individuelle Zahnanzahl schwankt daher zwischen 38 und 42. Wie bei den anderen Eigentlichen Maulwürfen ähnelt der untere Eckzahn den Schneidezähnen (incisiviform), während der vordere untere Prämolar vergrößert ist und eine eckzahnartige Gestalt (caniniform) besitzt. Die oberen Schneidezähne sind relativ klein, der Eckzahn groß und spitz. Die Länge der oberen Zahnreihe variiert von 12,5 bis 15,7 mm, die der unteren von 12,5 bis 14,4 mm.[1][4][2][3]

Skelettmerkmale
Der Kurzgesichtmaulwurf besitzt ein sogenanntes mogerides Becken. Bei dieser Form wird die Öffnung des vierten und fünften Kreuzbeinwirbels hinten von einer Knochenbrücke überdeckt. Es stimmt dadurch mit dem der Ostasiatischen Maulwürfe (Mogera) überein, weicht aber von den Südostasiatischen Maulwürfen (Euroscaptor) mit ihrem caecoidalen Becken ab. Der nahe verwandte Weißschwanzmaulwurf (Parascaptor) hat hingegen ein europaeoidales Becken, bei den Eurasischen Maulwürfen (Talpa) kommen sowohl caecoidale als auch europaeoidale Becken vor. Bei letzterer Struktur, benannt nach dem Europäischen Maulwurf (Talpa europaea), ist nur die Öffnung des vierten Kreuzbeinwirbels durch eine Knochenbrücke überspannt, bei ersterer, benannt nach dem Blindmaulwurf (Talpa caeca), fehlt die Brücke.[5][4]
Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 48, was innerhalb der Maulwürfe die höchste Anzahl ist. Er besteht aus 3 meta- bis submetazentrischen, 2 subtelozentrischen und 19 acrozentrischen Autosomenpaaren. Das Y-Chromosom und das X-Chromosom konnten noch nicht bestimmt werden. Dadurch kann auch die genaue Anzahl der Arme der Autosomen nicht angegeben werden (fundamentale Anzahl). Sie dürfte aber zwischen 54 und 56 liegen, was weitgehend den Verhältnissen bei den Ostasiatischen Maulwürfen entspricht, während die Eurasischen Maulwürfe eine weitaus höhere fundamentale Anzahl aufweisen.[6][3] Das gesamte Mitogenom setzt sich aus 16.699 Basenpaaren zusammen.[7]
Verbreitung und Lebensraum
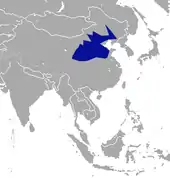
Der Kurzgesichtmaulwurf ist im nordöstlichen Teil von China beheimatet. Sein Verbreitungsgebiet erstreckt sich von West nach Ost über die Provinzen Gansu, Shaanxi, Hubei, Shanxi, Hebei, Henan bis nach Shandong und Jiangsu sowie weiter nordöstlich über Liaoning bis nach Heilongjiang. Es schließt auch die Autonomen Gebiete Ningxia und die östliche und zentrale Innere Mongolei mit ein. Die Art bewohnt dadurch den kalten und trockenen Bereich Chinas und ist an diese Bedingungen stärker angepasst als andere Vertreter der Maulwürfe. Als Lebensräume dienen sandige Graslandschaften, Wiesen und Lössregionen. Teilweise kommen die Tiere auch in Randgebieten von Wüsten vor, so etwa auf dem Ordos-Plateau.[2][3]
Lebensweise
Über die Lebensweise des Kurzgesichtmaulsurfs ist wenig bekannt, sie dürfte aber mit der der übrigen Eigentlichen Maulwürfe übereinstimmen. Demnach leben die Tiere größtenteils unter der Erde, wo sie Gangsysteme errichten. Ihre Nahrung besteht vorrangig aus Regenwürmern, Larven von Käfern und anderen Gliederfüßern.[2][3]
Systematik
Innere Systematik der Eigentlichen Maulwürfe nach He et al. 2016[8]
|
Der Kurzgesichtmaulwurf ist eine Art aus der Gattung Scaptochirus, die keine weiteren rezenten Vertreter beinhaltet. Die Gattung gehört wiederum zur Familie der Maulwürfe (Talpidae) und der Ordnung der Insektenfresser (Eulipotyphla). Sie bildet innerhalb der Maulwürfe gemeinsam mit einigen weiteren Artengruppen vorwiegend eurasischer Herkunft die Tribus der Eigentlichen Maulwürfe (Talpini). Diese stellt die größte Gruppe der Maulwürfe dar. Die Angehörigen der Tribus umfassen mittelgroße bis große Maulwürfe, die sich durch eine Anpassung an eine grabende Lebensweise auszeichnen. Dies drückt sich unter anderem durch die kurzen und kräftigen Ober- beziehungsweise Unterarme und die großen, schaufelartigen sowie nach außen gedrehten Hände aus. Die Hände der Eigentlichen Maulwürfe weisen ein Sesambein auf, der sogenannte Präpollex („Vordaumen“), das seitlich ansitzt und die Fläche dadurch vergrößert.[9][10] Ein weiteres gemeinsames Charakteristikum findet sich in dem kurzen Schwanz. Als besondere Skelettmerkmale können die verknöcherte Symphyse des Schambeins und das Fehlen eines Zahnwechsels hervorgehoben werden.[3] Laut molekulargenetischen Untersuchungen trennten sich die Eigentlichen Maulwürfe im Oberen Eozän vor rund 34 bis 36 Millionen Jahren von den anderen Triben der Maulwürfe ab. Eine stärkere Diversifizierung der Talpini fand im Mittleren Miozän vor rund 12 Millionen Jahren statt. Die Linie von Scaptichirus bildete sich hierbei vermutlich im Oberen Miozän heraus. Als nächster Verwandter dürfte der Weißschwanzmaulwurf (Parascaptor) anzusehen sein. Ebenfalls in einer näheren Beziehung stehen die Südostasiatischen Maulwürfe (Euroscaptor).[11][12][13][14][15][16][8][17]
Einzelne Autoren unterschieden bis zu drei Unterarten des Kurzgesichtmaulwurfs.[2] Im achten Band des Standardwerkes Handbook of the Mammals of the World aus dem Jahr 2018 wird dieser jedoch als monotypisch eingestuft.[3] Allerdings sind neben der rezenten Art noch mehrere fossile Formen beschrieben worden:
- Scaptochirus jiangnanensis Jin & Liu, 2008
- Scaptochirus primitivus Zdansky, 1928
Eine weitere Art, Scaptochirus primaevus, aufgestellt im Jahr 1924 von Max Schlosser an Fundmaterial von Ertemte in der Inneren Mongolei, wurde im Jahr 1983 in die Gattung Yanshuella ausgelagert und der Tribus der Neuweltmaulwürfe (Scalopini) zugeschlagen.[18][19] Von Li Yong-xiang wiederum stammt die Art Scaptochirus minor aus der Shanyangzhai-Höhle in der chinesischen Provinz Hebei, die Benennung erfolgte im Jahr 2012. Das spärliche Fossilmaterial dieser Form gehört aber wahrscheinlich eher zum Gansu-Maulwurf (Scapanulus oweni).[20]
Forschungsgeschichte

Die wissenschaftliche Erstbeschreibung des Kurzgesichtmaulswurfs geht auf Alphonse Milne-Edwards aus dem Jahr 1867 zurück. Hierin gab Milne-Edwards die geringere Zahnanzahl gegenüber den Eurasischen Maulwürfen (Talpa) als Unterscheidungsmerkmal an. Da sich auch die einzelnen Zähne als abweichend von den Eurasischen Maulwürfen erwiesen, wählte er für die neue Form mit Scaptochirus eine neue Gattung. Milne-Edwards Beschreibung beruhte auf einer zoologischen Sammlung, die Armand David in Ostasien zusammengetragen hatte. Als Herkunftsgebiet gab er lediglich „Mongolei“ an (en Mongolie).[21] Erst im Jahr 1910 wurde dies von Oldfield Thomas mit Zhangjiakou rund 160 km nordwestlich von Peking in der chinesischen Provinz Hebei präzisiert, was seitdem als Typusgebiet der Art gilt.[22]
Die generische Eigenständigkeit des Kurzgesichtmaulwurfs stand durch den Verweis zu Scaptochirus schon sehr früh fest. Nachfolgende Autoren zweifelten dies kaum an.[23][24][22][1] Ernst Schwarz allerdings vereinte im Jahr 1948 alle ost- und südostasiatischen Formen der talpinen Maulwürfe mit den Eurasischen Maulwürfen unter der Gattung Talpa. Hierbei unterschied er für den westeurasischen Bereich, also dem klassischen Verbreitungsgebiet der Eurasischen Maulwürfe, mehrere Arten. Die Vertreter des ost- und südostasiatischen Raumes versammelte er wiederum in einer Art, die er mit Talpa micrura bezeichnete; das Artepitheton bezieht sich heute auf den Himalaya-Maulwurf (Euroscaptor micrurus) aus der Gattung der Südostasiatischen Maulwürfe (Euroscaptor). Talpa micrura wiederum unterteilte er in mehrere Unterarten. Den Kurzgesichtmaulwurf hielt er so mit Talpa micrura moschata aus.[25] Das Konzept wurde insbesondere in den 1950er und 1960er Jahren vielfach übernommen, doch seitens einiger japanischer Zoologen zumindest für die Vertreter der Ostasiatischen Maulwürfe (Mogera) relativ früh wieder aufgegeben.[26] Doch auch bezüglich der weiteren ost- und südostasiatischen Formen kamen spätestens in den 1970er Jahren Zweifel auf, etwa durch Ivo Grulich. Dieser hatte die Zahn- und Skelettstruktur der Maulwürfe untersucht. Neben den bereits bekannten Differenzen in der Zahnformel konnte er so auch Abweichungen im Bau des Beckens aufzeigen. Da der Kurzgesichtmaulwurf durch sein mogerides Becken den Ostasiatischen Maulwürfen näherstand als den Eurasischen Maulwürfen, plädierte er für eine Eigenständigkeit der Gattung.[5][4] Im ausgehenden 20. Jahrhundert verlor sich dann Schwarz' Konzept wieder.[27]
Im gleichen Aufsatz aus dem Jahr 1910, in dem Thomas die Typuslokalität des Kurzgesichtmaulwurfs einengte, führte er mit Scaptochirus gilliesi eine weitere Art ein, die er auf einem ausgewachsenen Tier aus Hejin im Südwesten der Provinz Shanxi begründete, das aber deutlich kleiner war als der Kurzgesichtmaulwurf.[22] Bereits im Jahr 1881 hatte er mit Talpa leptura eine weitere Form aus der Umgebung von Peking benannt, die sich durch ihren dünnen Schwanz hervorhob. Das Individuum war 1860 von Robert Swinhoe während einer Expedition nach Ostasien gesammelt und im Jahr darauf von John Edward Gray als mögliche neue Maulwurfsart vorgestellt worden, jedoch ohne dieser einen Namen zu geben (in einem unveröffentlichten Manuskript verwendete Gray allerdings Talpa chinensis[1]).[28] Thomas erkannte Ähnlichkeiten zum Kurzgesichtmaulwurf, vermerkte aber, dass im Unterkiefer ein zusätzlicher Prämolar auftrat, so dass die Art in der Zahnformel dem Weißschwanzmaulwurf glich.[29] Wiederum 1910 ordnete er Talpa leptura in die Gattung Scaptochirus ein, da ähnliche Maulwürfe aus dem nordöstlichen China ebenfalls eine variierende Zahnanzahl zeigten.[22] Eine eigenständige Art und Gattung war im Jahr 1898 von Pierre Heude mit Chiroscaptor sinensis kreiert worden. Als Belegexemplar diente ein Individuum aus dem Südwesten der damaligen Provinz Zhili, was in etwa mit der heutigen Provinz Hebei übereinstimmt.[30] Zuletzt sei noch auf die von Sergei Uljanowitsch Stroganow im Jahr 1941 etablierte Form Parascaptor grandidens aus dem Großen Hinggan-Gebirge in der Inneren Mongolei verwiesen. Das ihr zugrunde gelegte Exemplar war zuvor im Jahr 1891 bei einer Expedition von Dmitri Wasiljewitsch Putjata aufgesammelt worden und wies im Oberkiefer vier Prämolaren auf.[31][4] Alle genannten Bezeichnungen gelten heute als synonym zum Kurzgesichtmaulwurf.[32]
Unabhängig der bereits genannten Synonyme stammt mit Scaptochirus davidianus eine Bezeichnung für einen Maulwurf aus der Region um Meydan Ekbaz im Norden Syriens, die von Milne-Edwards erstmals im Jahr 1884 verwendet worden war. Die enge Bindung der beiden Formen begründete er mit einer identischen Zahnanzahl.[33] Diese heute als Pater-David-Maulwurf bekannte Art galt dadurch lange Zeit als mit dem Kurzgesichtmaulwurf verwandt und von diesem räumlich weit isoliert. Im Jahr 1948 sah Ernst Schwarz eine nähere Beziehung zum Blindmaulwurf (Talpa caeca) und somit zu den Eurasischen Maulwürfen. Allerdings schien eine anatomische Studie aus dem Jahr 1982 von Ivo Grulich die engere Bindung an den Kurzgesichtmaulwurf zu bestätigen. Grulich vermutete dies anhand des Beckenaufbaus, da er beim Kurzgesichtmaulwurf einen mogeriden Typus erkannte, der seiner Meinung nach auch beim Pater-David-Maulwurf bestand.[4] Nur wenige Jahre später erbrachte aber eine erneute Untersuchung beim Pater-David-Maulwurf einen caecoidalen Beckenbau, wodurch die Form wieder in die Nähe der Eurasischen Maulwürfe rückte. Folgend darauf bestätigte eine weitere Analyse im Jahr 2001 auch bezüglich der Gebissstruktur eine stärkere Ähnlichkeit mit den Eurasischen Maulwürfen. Die von Milne-Edwards beobachtete Übereinstimmung der Zahnformel mit dem Kurzgesichtmaulwurf konnte auf die starke Oligodontie beim Pater-David-Maulwurf zurückgeführt werden.[34] Vor allem die seit dem 21. Jahrhundert verstärkt vorgenommenen genetischen Untersuchungen unterstreichen die Stellung des Pater-David-Maulwurfs innerhalb der Gattung Talpa und die deutliche Trennung von Scaptochirus.[16][35]
Stammesgeschichte
Die Gattung Scaptochirus verfügt über einen recht umfangreichen Fossilbericht. Im Gegensatz zum heutigen Kurzgesichtmaulwurf mit seiner Beschränkung auf das nordöstliche China war die frühere Verbreitung deutlich ausgedehnter. Einzelne Funde sind bereits im Pliozän nachweisbar. Hierzu gehören ein unterer Molar aus der Gaozhuang-Formation, deren Alter mit rund 4,3 Millionen Jahren angegeben wird, und ein 14 mm langer Oberarmknochen aus der Mazegou-Formation, die wiederum vor rund 3,5 Millionen Jahren entstand. Beide Gesteinseinheiten sind im Yushe-Becken der nordchinesischen Provinz Shanxi aufgeschlossen. Die Funde ähneln den entsprechenden Elementen beim Kurzgesichtmaulwurf, wurden aber keiner bestimmten Art zugewiesen. Möglicherweise ähnlich alt sind Reste aus Shouyang in der gleichen Provinz, die aber noch nicht genau analysiert wurden.[19] Im Altpleistozän erstreckte sich das Vorkommen von Scaptochirus dann bis in den Süden des heutigen Chinas, wie einige Zahnfunde aus der Gigantopithecus-Höhle von Juyuan im Karstgebiet der Boyue-Berge bei Chongzuo im Autonomen Gebiet von Guangxi zeigen. Auch hier erfolgte keine genaue Artzuweisung.[36] Etwa gleichalt könnten die Funde aus der Höhle Renzidong am Hang des Laili-Hügels bei Fanchang in der ostchinesischen Provinz Anhui sein. Die Reste von mehr als zehn Individuen wurden im Jahr 2000 erstmals vorgestellt,[37] acht Jahre später dann der Art Scaptochirus jiangnanensis zugewiesen, die sich durch ihre geringere Größe im Vergleich zum heutigen Kurzgesichtmaulwurf auszeichnet.[19] Von großer Bedeutung sind die Funde aus Choukoutien am Longgushan („Drachenknochenberg“) südwestlich von Peking. Die komplexe Fundstelle, an der der Peking-Mensch definiert wurde, enthält in der Lokalität 1 umfangreiche Fossilreste von Maulwürfen, häufig in Form von Zähnen, die bereits 1928 von Otto Zdansky als zur Art Scaptochirus primitivus gehörig beschrieben wurden. Nachfolgend diskutierten einzelne Autoren die Eigenständigkeit der Art gegenüber dem Kurzgesichtmaulwurf.[38][19] Nach radiometrischen Datierungen gehören die fossilführenden Ablagerungen weitgehend in den älteren Abschnitt des Mittelpleistozäns. Deutlich jünger sind die Funde der unmittelbar benachbarten Lokalität 4, welche dem ausgehenden Mittelpleistozän zuzuschreiben sind. Eventuell trat hier bereits der Kurzgesichtmauslwurf in seiner heutigen Ausprägung auf. Von diesem liegen wiederum zahlreiche Reste aus der Shanyangzhai-Höhle bei Qinhuangdao in der Provinz Hebei vor. Das insgesamt reiche Fossilmaterial datiert ebenfalls in das Mittelpleistozän.[39][20] Allerdings wird der heutige Kurzgesichtmaulwurf mitunter bereits für die alt- oder mittelpleistozäne Fundstelle von Gongwangling, der Fundstelle des Lantian-Menschen, in der zentralchinesischen Provinz Shaanxi angegeben.[40]
Bedrohung und Schutz
Laut IUCN ist der Bestand des Kurzgesichtmaulwurfs „nicht gefährdet“ (least concern) Begründet wird dies mit der weiten Verbreitung und der relativ hohen Populationsdichte. Größere Gefährdungen für den Bestand sind nicht bekannt. Die Art ist in zahlreichen Naturschutzgebieten präsent.[41]
Literatur
- Robert S. Hoffmann und Darrin Lunde: Order Soricomorpha – Shrews and Moles. In: Andrew T. Smith und Yan Xie (Hrsg.): A Guide to the Mammals of China. Princeton University Press, 2008, ISBN 978-0-691-09984-2, S. 325
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 619) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9.
Einzelnachweise
- Glover M. Allen: The Mammals of China and Mongolia. Natural History of Central Asia 9 (1), New York, 1938, S. 1–620 (S. 72–76) ()
- Robert S. Hoffmann und Darrin Lunde: Order Soricomorpha – Shrews and Moles. In: Andrew T. Smith und Yan Xie (Hrsg.): A Guide to the Mammals of China. Princeton University Press, 2008, ISBN 978-0-691-09984-2, S. 325
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 619) ISBN 978-84-16728-08-4
- Ivo Grulich: Zur Kenntnis der Gattungen Scaptochirus und Parascaptor (Talpini, Mammalia). Folia Zoologica 31, 1982, S. 1–20 ()
- Ivo Grulich: Zum Bau des Beckens (Pelvis), eines systematisch-taxonomischen Merkmales, bei der Unterfamilie Talpinae. Zoologické Listy 20, 1971, S. 15–28 ()
- Shin-ichiro Kawada, Masashi Harada, Kazuhiru Koyasu und Sen-ichi Oda: Karyological note on the short-faced mole, Scaptochirus moschatus (Insectivore, Talpidae). Mammal Study 27, 2002, S. 91–94, ()
- Lei Chen, Di Xu und Mengyao Sun: The complete mitochondrial genome of short-faced mole (Scaptochirus moschatus, Talpidae). Mitochondrial DNA Part B 7 (1), 2022, S. 79–80, doi:10.1080/23802359.2021.2008833
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang, Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumenting the polydactyly ‚constraint‘: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- Constanze Bickelmann, Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Transcriptional heterochrony in talpid mole autopods. EvoDevo 3, 2012, S. 16, doi:10.1186/2041-9139-3-16
- Akio Shinohara, Shin-ichiro Kawada, Masashi Harada, Kazuhiro Koyasu, Sen-ichi Oda und Hitoshi Suzuki: Phylogenetic relationships of the short-faced mole, Scaptochirus moschatus (Mammalia: Eulipotyphla), among Eurasian fossorial moles, as inferred from mitochondrial and nuclear gene sequences. Mammal Study 33, 2008, S. 77–82
- E. D. Zemlemerova, A. A. Bannikova, A. V. Abramov, V. S. Lebedev und V. V. Rozhnov: New Data on Molecular Phylogeny of the East Asian Moles. Doklady Biological Sciences 451, 2013, S. 257–260
- Kai He, Akio Shinohara, Xue-Long Jiang und Kevin L. Campbell: Multilocus phylogeny of talpine moles (Talpini, Talpidae, Eulipotyphla) and its implications for systematics. Molecular Phylogenetics and Evolution 70, 2014, S. 513–521, doi:10.1016/j.ympev.2013.10.002
- Akio Shinohara, Shin-Ichiro Kawada, Nguyen Truong Son, Chihiro Koshimoto, Hideki Endo, Dang Ngoc Can und Hitoshi Suzuki: Molecular phylogeny of East and Southeast Asian fossorial moles (Lipotyphla, Talpidae). Journal of Mammalogy 95 (3), 2014, S. 455–466
- Akio Shinohara, Shin-ichiro Kawada, Nguyen Truong Son, Dang Ngoc Can, Shinsuke H. Sakamoto und Chihiro Koshimoto: Molecular phylogenetic relationships and intra-species diversities of three Euroscaptor spp. (Talpidae: Lipotyphla: Mammalia) from Vietnam. Raffles Bulletin of Zoology 63, 2015, S. 366–375
- Anna A. Bannikova, Elena D. Zemlemerova, Paolo Colangelo, Mustafa Sözen, M. Sevindik, Artem A. Kidov, Ruslan I. Dzuev, Boris Kryštufek und Vladimir S. Lebedev: An underground burst of diversity – a new look at the phylogeny and taxonomy of the genus Talpa Linnaeus, 1758 (Mammalia: Talpidae) as revealed by nuclear and mitochondrial genes. Zoological Journal of the Linnean Society 175, 2015, S. 930–948
- E. D. Zemlemerova, A. A. Bannikova, V. S. Lebedev, V. V. Rozhnov, A. V. Abramov: Secrets of the underground Vietnam: an underestimated species diversity of Asian moles (Lipotyphla: Talpidae: Euroscaptor). Proceedings of the Zoological Institute RAS 320 (2), 2016, S. 193–220
- Gerhard Storch und Z. Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Lawrence J. Flynn und Wen-Yu Wu: The Lipotyphla of Yushe Basin. In: Lawrence J. Flynn und Wen-Yu Wu (Hrsg.): Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals, Volume II: Small Mammal Fossils of Yushe Basin, Vertebrate Paleobiology and Paleoanthropology. Springer Science+Business Media B.V., 2017, S. 11–26
- Yong-xiang Li, Ji Li und Yun-xiang Zhang: Fossil Scapanulus oweni (Eulipotyphla, Mammalia) from the Shanyangzhai Cave, Middle Pleistocene, Qinhuangdao, China. Quaternary International 392, 2016, S. 197–202, doi:10.1016/j.quaint.2015.08.001
- Alphonse Milne-Edwards: Observations sur quelques mammifères du Nord de la Chine. Annales du sciences naturelles 5 (7), 1867, S. 375–377 ()
- Oldfield Thomas: A new Chinese mole of the genus Scaptochirus. Annals and Magazine of Natural History 8 (5), 1910, S. 350–351 ()
- Theodore Gill: Synopsis of Insectivorous Mammals. Washington D.C., 1875, S. S. 1–32 (S. 22) ()
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 (S. 140 und 161) ()
- Ernst Schwarz: Revision of the Old‐World Moles of the Genus Talpa Linnaeus. Proceedings of the Zoological Society of London 118 (1), 1948, S. 36–48
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture, Hokkaido University 55, 1967, S. 191–265 ()
- Shin-ichiro Kawada: The historical notes and taxonomic problems of East Asian moles, Euroscaptor, Parascaptor and Scaptochirus, of continental Asia (Insectivora, Talpidae). Mammal Study 30, 2005, S. S5–S11
- John Edward Gray (ohne Titel). Proceedings of the Zoological Society of London, 1861, S. 390 ()
- Oldfield Thomas: Description of a new species of mole from China. Annals and Magazine of Natural History 5 (7), 1881, S. 469–471 ()
- Pierre Heude: Talpidés. Mémoires concernant l’histoire naturelle de lempire chinois par des pères de la Compagnie de Jésus 4, 1898, S. 36–43 ()
- Sergei U. Stroganov: Insectivore mammals of the fauna of the USSR. Doklady Akademii Nauk 33 (3), 1941, S. 270–272
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Alphonse Milne-Edwards: Sur la Classification des Taupes de l'ancien continent. Comptes rendus des seances de l'Academie des Sciences 99, 1884, S. 1141–1143 ()
- Boris Kryštufek, Friederike Spitzenberger und H. Kefelioğlu: Description, taxonomy, and distribution of Talpa davidiana. Mammalian Biology 66, 2001, S. 135–143
- Sadık Demırtaş, Metin Silsüpür, Jeremy B. Searle, David Bilton und İslam Gündüz: What should we call the Levant mole? Unravelling the systematics and demography of Talpa levantis Thomas, 1906 sensu lato (Mammalia: Talpidae). Mammalian Biology 100, 2020, S. 1–18, doi:10.1007/s42991-020-00010-4
- Yuan Wang, Changzhu Jin, Wenshi Pan, Dagong Qin, Yaling Yan, Yingqi Zhang, Jinyi Liu, Wei Dong und Chenglong Deng: The Early Pleistocene Gigantopithecus-Sinomastodon fauna from Juyuan karst cave in Boyue Mountain, Guangxi, South China. Quaternary International 434 (A), 2017, S. 4–16, doi:10.1016/j.quaint.2015.11.071
- Jin Chang-zhu, Dong Wei, Liu Jin-yi, Wei Guan-biao, Xu Qin-qi, Zheng Jia-jian, Zheng Long-ting, Han Li-gang und Wang Fa-zhi: A Preliminary Study on the Early Pleistocene Deposits and the Mammalian Fauna from the Renzi Cave, Fanchang, Anhui, China. Acta Anthropologica Sinica 19 (suppl.), 2000, S. 235–245
- Chao Tze-kuei und Li Yen-hsien: Report on the excavation of the Choukoutien Sinanthropus site in 1959. Vertebrata Palasiatica 4 (1), 1960, S. 30–32
- Yun Xiang Zhang, Li Yong Xiang, Wei Wang und Hu Jun Gong: Middle Pleistocene mammalian fauna of Shanyangzhai cave in Qinhuangdao area, China and its zoogeographical significance. Chinese Science Bulletin 55 (1), 2010, S. 72–76, doi:10.1007/s11434-009-0360-7
- Dong Wei, Jin Chang-zhu, Xu Qin-qi, Liu Jin-yi, Tong Hao-wen und Zheng Long-ting: A Comparative Analysis on the Mammalian Faunas Associated with Homo erectus in China. Acta Anthropologica Sinica 19 (suppl.), 2000, S. 246–256
- A. T. Smith und C. H. Johnston: Scaptochirus moschatus (errata version published in 2017). The IUCN Red List of Threatened Species 2016. e.T41476A115188844 (); zuletzt aufgerufen am 2. März 2021
Weblinks
- Scaptochirus moschatus in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: A. T. Smith & C. H. Johnston, 2016. Abgerufen am 2. März 2021.