Kleiner Japanischer Maulwurf
Der Kleine Japanische Maulwurf (Mogera imaizumii, Syn.: Mogera minor) ist eine Säugetierart aus der Gattung der Ostasiatischen Maulwürfe innerhalb der Maulwürfe (Talpidae). Er kommt in Japan vor und bewohnt den östlichen und zentralen Teil der Insel Honshū. Im westlichen Bereich der Insel und auf einigen weiteren Inseln sind einzelne verstreute Subpopulationen verbreitet. Die Tiere nutzen vor allem Landschaften mit mächtiger Bodendecke. In einigen Regionen besteht eine direkte Konkurrenz zum Japanischen Maulwurf und zum Echigo-Maulwurf. Es handelt sich beim Kleinen Japanischen Maulwurf um einen mittelgroßen bis großen Vertreter der Ostasiatischen Maulwürfe. Wie seine Verwandten charakterisiert ihn ein zylindrischer Körper, ein kurzer Hals und zu Grabschaufeln umgewandelte Vorderbeine. Das Fell ist meist in dunkleren Brauntönen gehalten.
| Kleiner Japanischer Maulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Kleiner Japanischer Maulwurf (Mogera imaizumii) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Mogera imaizumii | ||||||||||||
| (Kuroda, 1957) |
Die Tiere leben unterirdisch in selbst gegrabenen Tunneln und Gängen. Sie unterhalten Reviere, deren Größe von äußerlichen Bedingungen und der Paarungszeit beeinflusst wird. Als Nahrung dienen vor allem Regenwürmer und Insekten. Die Fortpflanzung erfolgt überwiegend vom Spätwinter bis Frühjahr, gelegentlich tritt eine zweite Fortpflanzungsphase im Sommer auf. Bei einem Wurf kommen zwei bis sechs Jungen zur Welt. Die Art erhielt im Jahr 1957 ihren wissenschaftlichen Namen, wurde aber bereits 21 Jahre vorher eingeführt. Eine Zeitlang bestand eine Verwechslung in der Namensgebung mit dem Japanischen Maulwurf. Unterarten werden nicht anerkannt, allerdings gibt es verschiedene genetisch unterscheidbare Linien. Der Kleine Japanische Maulwurf gilt als nicht gefährdet.
Merkmale
Habitus

Der Kleine Japanische Maulwurf ist ein mittelgroßer bis großer Vertreter der Ostasiatischen Maulwürfe, aber kleiner als der Japanische Maulwurf (Mogera wogura) und der Sado-Maulwurf (Mogera tokudae). Die Kopf-Rumpf-Länge beträgt 10,2 bis 15,4 cm, die Schwanzlänge 0,8 bis 2,3 cm. Der Schwanz entspricht dadurch 6,9 bis 17,5 % der Länge des übrigen Körpers. Das Gewicht variiert von 36 bis 109 g. Hinsichtlich der Körpergröße gibt es starke geographische Unterschiede. Generell sind die Tiere in Schwemmland-Tiefebenen größer als im Bergland oder in engen Tälern. Daneben bestehen Verbindungen zu Niederschlagsmengen und Jahrestemperaturen. So werden Individuen in Regionen mit kräftigen Schneefällen kleiner als in solchen ohne.[1] Auch sind jährliche Gewichtsschwankungen feststellbar,[2] ebenso wie eine Tendenz zu einem geringeren Körpergewicht bei direkter Konkurrenz zu anderen Ostasiatischen Maulwürfen.[3] Prinzipiell ähnelt der Kleine Japanische Maulwurf anderen Gattungsvertretern. Sein Körper ist zylinderförmig und der Hals kurz, die Vorderbeine sind zu Grabschaufeln umgewandelt. Deren Handflächen werden im Mittel 1,6 cm lang und 1,7 cm breit. Die Hinterfüße weisen ebenfalls durchschnittlich 1,6 cm Länge auf. Das Fell ist variabel gefärbt, es tendiert zu einem dunkleren Braun bei Populationen in gebirgigen Landschaften und zu einem etwas helleren Braun bei solchen in Tiefebenen. Letztere weisen häufig einen rötlich braun bis nelkenbraun gefärbten Rücken sowie eine etwas hellere, zimtbraune Unterseite auf. Erstere sind generell oberseits nelkenbraun, unterseits marginal heller. Im Winter dunkelt das Fell allgemein ab. Vor allem auf der Insel Kinkasan ist mit 90 % ein hoher Anteil von albinotischen Individuen belegt.[4][5][6]
Schädel- und Gebissmerkmale
Der Schädel weist Längendurchschnittswerte auf, die je nach Region zwischen 34,4 und 36,7 mm variieren. Die größte Breite des Hirnschädels besitzt dementsprechend Werte von 16,5 bis 17,6 mm. Die schlanken Jochbögen weiten sich auf 12,2 bis 13,0 mm. Insgesamt auffällig sind das schmale Rostrum und die weite Zwischenaugenregion. Bei letzterer verengt sich der Schädel auf 7,8 bis 8,0 mm. Der Hirnschädel ist gerundet und weniger eckig als beim Japanischen Maulwurf. Der Unterkiefer wird je nach Region 21,7 bis 23,3 mm lang. Das Gebiss weist 42 Zähne auf, zusammen bilden sie folgende Zahnformel: Gelegentlich treten aber Individuen mit Zahnanomalien auf. Mit 1,8 % ist der Anteil allerdings deutlich geringer als bei anderen Ostasiatischen Maulwürfen Japans. In der Regel sind dabei die jeweils zweiten Prämolaren der oberen und unteren Zahnreihe involviert, selten auch der erste Vormahlzahn. In allen bisher dokumentierten Fällen handelt es sich um fehlende Zähne.[7] Die Schneidezähne stehen in der oberen Zahnreihe V- oder U-förmig zueinander, bei Populationen mit deutlich größeren Individuen meist breit V-förmig. Außerdem sind sie markant nach vorn gelehnt. In der Regel nimmt bei kleinen Individuen die Größe der Schneidezähne von vorn nach hinten ab, bei größeren Tieren haben sie etwa die gleichen Ausmaße. Die Prämolaren bilden keine so kompakte Reihe wie beim Japanischen Maulwurf. Die obere Zahnreihe erstreckt sich über eine Länge von 12,7 bis 16,3 mm, die unter über 11,3 bis 15,2 mm.[4][8][6]

Skelettmerkmale
Die Wirbelsäule setzt sich aus 7 Hals-, 14 Brust-, 5 Lenden-, 6 Kreuzbein- und 12 Schwanzwirbeln zusammen.[6]
Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 36. Er besteht aus 4 metazentrischen, 3 submetazentrischen, 3 subtelozentrischen und 7 acrozentrischen Autosomenpaaren. Das Y-Chromosom ist fleckenartig, das X-Chromosom submetazentrisch.[9][10][6]
Verbreitung und Lebensraum
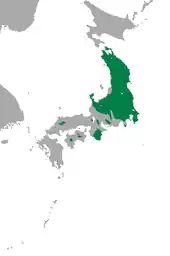
Der Kleine Japanische Maulwurf ist in Japan heimisch. Sein Vorkommen umfasst größere Teile der Hauptinsel Honshū zuzüglich einiger vorgelagerter Inseln. Ein großes und zusammenhängendes Verbreitungsgebiet besteht im nordwestlichen und zentralen Teil der Insel. Dessen südwestliche Grenze verläuft in etwa durch die Präfekturen Ishikawa, Gifu, Nagano und Shizuoka. Südwestlich davon tritt der Kleine Japanische Maulwurf in einzelnen Inselpopulationen auf. Dies betrifft etwa die Regionen um Shōbara, Kyoto und Ōno[11] sowie die Kii-Halbinsel auf Honshū, aber auch die Gebiete um die Berge Ishizuchi und Tsurugi auf der Insel Shikoku sowie die Insel Shōdoshima in der Seto-Inlandsee.[6]
Vor allem an der Südwestgrenze des zusammenhängenden Verbreitungsgebietes im zentralen Honshū kann es zu einem sympatrischen oder parapatrischen Aufeinandertreffen mit dem Japanischen Maulwurf kommen. Ersteres ist beispielsweise in der Schwemmebene des Tadori in der Kanazawa-Ebene der Präfektur Ishikawa belegt. Hier nutzt der Kleine Japanische Maulwurf härtere Böden, der Japanische Maulwurf hingegen weichere.[12] In einzelnen Regionen wird der Kleine Japanische Maulwurf vom Japanischen Maulwurf verdrängt, wobei wiederum härtere Böden die Ausbreitung von letzterem behindern. Jedoch verschoben sich im Zeitraum von 1959 und 2009 die Grenzen im Kontaktgebiet der beiden Arten um 2,4 bis 16 km Richtung Nordost zugunsten des Japanischen Maulwurfs.[13][14] In weiteren Bereichen Honshūs überschneidet sich der Lebensraum des Kleinen Japanischen Maulwurfs mit dem des Japanischen Bergmaulwurfs (Oreoscaptor mizura). In den Echigo-Ebenen der Präfektur Niigata grenzt das Vorkommen des Kleinen Japanischen Maulwurfs an das des Echigo-Maulwurfs (Mogera etigo). Dabei findet sich letzterer im Rückzug gegenüber ersteren.[15][5][6]
In der Regel ist der Kleine Japanische Maulwurf in ebenen Gebieten mit mächtigen und gut durchfeuchteten Böden anzutreffen. Die Höhenverbreitung reicht vom Meeresspiegelniveau bis auf rund 2000 m Gebirgslage.[12][6]
Lebensweise
Territorialverhalten
Über die Sozialstruktur des Kleinen Japanischen Maulwurfs liegen keine Informationen vor, es wird jedoch angenommen, dass er einzelgängerisch lebt. Die Tiere legen unterirdische Tunnel und Gänge an. Diese können in unterschiedlichen Tiefen von 4 bis 9 cm, 10 bis 21 cm und 28 bis 42 cm verlaufen. Tiefere Bereiche werden zumeist im Winter aufgesucht. Untersuchungen ergaben, dass eine Bodentemperatur von 23 °C für die flach angelegten Gänge optimale Voraussetzung ist. Die einzelnen Tunnelröhren haben einen Durchmesser von im Mittel 3,9 cm. In den Echigo-Ebenen liegt der Durchmesser bei 4,0 bis 4,5 cm und ist dadurch deutlich geringer als bei dem ebenfalls dort lebenden Echigo-Maulwurf.[15] In der Region um Kyoto lassen sich die Gänge mit einem Durchmesser von 3,0 bis 4,6 cm gut von den kleineren des Japanischen Bergmaulwurfs und den größeren des Japanischen Maulwurfs abtrennen.[16] Die Gangsysteme werden teilweise durch Pilze aus der Gattung der Fälblinge angezeigt, die möglicherweise mit Latrinen in der Umgebung in Verbindung stehen. Die Gangsysteme und die darin enthaltenen Nester, die aus Pflanzenmaterial bestehen, werden über längere Zeit besiedelt. In einzelnen Fällen konnte eine Nutzung über fünf Jahre belegt werden. Wahrscheinlich geschieht dies über eine Generationenabfolge, in einem Fall ließ sich auch ein Wechsel vom Kleinen Japanischen Maulwurf zum Japanischen Maulwurf dokumentieren.[17][16][6]
Der Kleine Japanische Maulwurf zeigt ein ausgeprägtes territoriales Verhalten. Reviere sind zwischen 113 und 8165 m² groß. Die größten Gebiete unterhalten Männchen während der Paarungszeit, wenn sie die Grenzen ihrer Reviere deutlich ausdehnen. Daneben besteht eine negative Verbindung zur lokalen Biomasse der Beutetiere, das heißt, die Territorien sind größer in Regionen mit geringerer Beutedichte. Innerhalb eines 24-Stunden-Tages konnten drei bis vier Aktivitätsperioden festgestellt werden, die jeweils von Ruhephasen unterbrochen werden. Eine Aktivitätsdauer beläuft sich auf 107 bis 341 Minuten. Innerhalb von fünf Minuten legt ein Tier 0,7 bis 4,1 m zurück.[18][6]
Ernährung
Die Hauptnahrung des Kleinen Japanischen Maulwurfs besteht nach Untersuchung von rund 150 Mageninhalten aus Regenwürmern vor allem der Gattung Pheretima. Diese machen rund 45 % der Nahrungsmenge aus, wobei ein geringer Prozentsatz auf Eier entfällt. Weitere knapp 40 % setzen sich aus Insekten zusammen. Besonders häufig sind hierbei Grillen, Schmetterlinge sowie verschiedene Käfer wie Laufkäfer, Blatthornkäfer und Schnellkäfer vertreten, aber auch Zweiflügler, etwa Schnaken und Bremsen beziehungsweise Ameisen. Gelegentlich wurden kleiner Wirbeltiere wie Frösche nachgewiesen. Während bei den Käfern sowohl ausgewachsene Individuen als auch die Larven vertilgt werden, sind es bei den Schmetterlingen und Zweiflüglern fast ausschließlich die Jungstadien. Darüber hinaus erreicht pflanzliches Material wie Samen und Gräser einen Anteil von über 9 %.[19][6]
Seine Beute erspäht der Kleine Japanische Maulwurf mit Hilfe des Geruchssinn. Die Regenwürmer werden entweder mit einem kurzen Biss in die Tunnel gezogen oder mit den Vorderfüßen ausgegraben. Anschließend tötet oder betäubt er sie durch Schläge auf den Boden oder mit Bissen in variable Körperpartien. In gut 29 % der Fälle werfen die Regenwürmer Teile ihres Körpers dabei ab. Anschließend frisst der Kleine Japanische Maulwurf entweder den Kopf oder den Schwanz zuerst, wechselt aber mitunter die Körperpartie. Für einen rund 1 bis 2 g schweren Regenwurm benötigt ein Tier rund 3,4 Sekunden, für einen 7 bis 8 g schweren Wurm etwa 92 Sekunden. Typisch sind dabei Rückwärtsbewegungen mit dem Kopf. Nicht vertilgte Regenwürmer werden aufbewahrt.[20][6]
Fortpflanzung
Die Paarungszeit beginnt beim Kleinen Japanischen Maulwurf häufig schon im späten Winter und setzt sich durch den gesamten Frühling fort. Dies lässt sich unter anderem bei den Männchen durch eine Vergrößerung der Hoden und eine Zunahme in der Produktion von Spermien in diesem Zeitraum feststellen.[21] Außerdem wurden trächtige Weibchen zumeist in den Monaten April bis Juli beobachtet. Es gibt aber selten eine zweite Fortpflanzungsphase im Sommer vom Juli bis August. Diese wurde beispielsweise häufiger bei Tieren in den Echigo-Ebenen dokumentiert, wo der sympatrisch auftretende Echigo-Maulwurf dieses Verhalten kaum zeigt.[3] Die Wurfgröße beträgt zwei bis sechs Junge. Der frühjährliche Nachwuchs ist zumeist im August ausgewachsen,[2] die Geschlechtsreife wird erst im Folgejahr erreicht.[19][6]
Bei genetischen Untersuchungen von drei Populationen, die sich in der Präfektur Yamagata in 16 bis 38 km Entfernung zueinander befanden, bestanden innerhalb der jeweiligen Gruppe nur wenige individuelle Differenzierungsmöglichkeiten. Dies könnte darauf zurückzuführen sein, dass es beim Kleinen Japanischen Maulwurf nicht zu einer stärkeren Abwanderung der Jungtiere nach der Entwöhnung kommt, wie es bei einigen anderen Vertretern der grabend lebenden Maulwürfe belegt ist. Die Studien fanden allerdings außerhalb der Paarungszeit statt, so dass hier auch externe Einflüsse wie die Häufigkeit der Beutetiere einen Einfluss auf die Wanderungsbewegungen und die regionale Gruppenstruktur haben. Allerdings ergab die Studie deutliche Unterschiede der Gruppen zueinander, was weitgehend durch Gendrift erklärt wird.[22] Für eine Population nördlich von Tokio ließ sich eine prinzipielle Altersgruppierung feststellen. Demnach setzte diese sich zu 63 % aus Jungtieren und zu 26 % aus zweijährigen Tieren zusammen. Der Rest entfiel auf drei- bis vierjährige Individuen, wobei letztere nur einen Anteil von 3 % ausmachten.[23][6]
Fressfeinde und Parasiten
Als bedeutendste Fressfeinde treten Eulen, Greifvögel, Füchse, Dachse und Wiesel auf.[6] Unter den Greifvögeln ist unter anderem der Nepalhaubenadler zu nennen.[24][25] Innere Parasiten sind mit Saugwürmern wie Echinostoma, mit Bandwürmern wie Hymenolepis und Choanotaenia sowie mit Fadenwürmern dokumentiert. Zu letzteren gehören unter anderem Ascarops, Parastrongyloides, Protospirura, Tricholinstowia, Trichuris und Capillaria.[26][27] Teilweise wurden beim Kleinen Japanischen Maulwurf auch Fadenwürmer gefunden, die normalerweise an Insekten parasitieren und so wohl mit der Nahrung in den Magen-Darm-Trakt gelangten.[28] Es besteht außerdem ein hoher Befall an Kokzidien.[29] Die Nester der Tiere werden teilweise von Asseln aufgesucht, etwa der Gattung Lucasioides.[30]
Systematik
Innere Systematik der Ostasiatischen Maulwürfe nach Shinohara et al. 2014[31]
|
Innere Systematik der Ostasiatischen Maulwürfe nach Kirihara et al. 2013, beschränkt auf die Arten Japans[32]
|
Der Kleine Japanische Maulwurf ist eine Art innerhalb der Gattung der Ostasiatischen Maulwürfe (Mogera), zu der insgesamt neun Arten gehören. Diese werden wiederum zur Familie der Maulwürfe (Talpidae) gezählt, innerhalb derer sie gemeinsam mit einigen anderen, vorwiegend eurasisch verbreiteten Artengruppen die Tribus der Eigentlichen Maulwürfe (Talpini) formen. Die Tribus vereint die zumeist grabenden Vertreter der Maulwürfe, andere Mitglieder der Familie leben dagegen nur teilweise unterirdisch, bewegen sich oberirdisch fort oder sind an eine semi-aquatische Lebensweise angepasst.[33] Molekulargenetische Untersuchungen sprechen für eine Aufteilung der Ostasiatischen Maulwürfe in zwei Verwandtschaftslinien. Von diesen schließt eine die ost- und südostasiatisch-festländischen Vertreter um den Chinesischen Inselmaulwurf (Mogera insularis) ein, die andere setzt sich aus den Arten der japanischen Inselwelt um den Japanischen Maulwurf (Mogera wogura) zusammen. Letztere beinhalten zusätzlich noch den Ussuri-Maulwurf (Mogera robusta) der Koreanischen Halbinsel und der fernöstlichen russischen Region Primorje. Die Teilung der Ostasiatischen Maulwürfe in diese beiden Linien reicht bis in das Obere Miozän vor etwa 10 bis 8 Millionen Jahre zurück. Der Kleine Japanische Maulwurf ist Teil der zweiten genannten Linie, deren weitere Differenzierung in das Obere Pliozän und das Untere Pleistozän vor rund 3,6 bis 1,2 Millionen Jahren datiert. Er bildet in mehreren genetischen Untersuchungen die Schwestergruppe einer Klade aus dem Japanischen Maulwurf und dem Ussuri-Maulwurf.[34][31][35] Die drei Arten können unter anderem cytogenetisch unterschieden werden. Sie weisen jeweils einen Chromosomensatz von 2n = 36 auf, die Anzahl der Arme (fundamentale Anzahl) der Autosomen variiert aber von 52 beim Japanischen Maulwurf über 54 beim Kleinen Japanischen Maulwurf bis hin zu 58 beim Ussuri-Maulwurf.[9][10] Nach Meinung einiger Autoren repräsentieren der Kleine Japanische Maulwurf und der Japanische Maulwurf eine eher späte Einwanderungswelle der Ostasiatischen Maulwürfe auf den japanischen Inseln. Diese besiedelte vor allem die Inlandgebiete, während eine früher eingewanderte Gruppe mit dem Sado-Maulwurf (Mogera tokudae) und dem Echigo-Maulwurf (Mogera etigo) auf die Küstenländern und einigen vorgelagerten Inseln beschränkt blieb.[36][37][32]
Die wissenschaftliche Artbezeichnung Mogera imaizumii geht auf Nagamichi Kuroda aus dem Jahr 1957 zurück, wobei Kuroda Talpa micrura imaizumii verwendete. Die relativ kurz gehaltene Notiz stellt lediglich eine Namenskorrektur dar und bezieht sich auf einen in japanisch verfassten Aufsatz von Kuroda aus dem Jahr 1936, in dem er die Erstbeschreibung des Kleinen Japanischen Maulwurfs unter der wissenschaftlichen Bezeichnung Mogera wogura minor ausgeführt hatte. Diese basierte auf einzelnen Individuen aus Shiobara in der Präfektur Tochigi im zentralen Teil der japanischen Hauptinsel Honshū. Die Region bildet das Typusgebiet der Art. Der Holotypus einschließlich von vier Paratypen gingen bei einem Feuer im Jahr 1945 verloren.[38] Für die gut 21 Jahre später vorgenommene Namensänderung sah sich Kuroda durch eine zuvor im Jahr 1948 durch Ernst Schwarz veröffentlichte Revision der Eigentlichen Maulwürfe veranlasst. Schwarz' Revision beinhaltete eine Vereinigung aller talpinen ost- und südostasiatischen Maulwürfe mit den Eurasischen Maulwürfen unter der Gattungsbezeichnung Talpa. Darüber hinaus fasste sie alle ost- und südostasiatischen Maulwürfe in einer Art, Talpa micrura, zusammen (das Artepitheton micrura bezieht sich heute auf den Himalaya-Maulwurf aus der Gattung der Südostasiatischen Maulwürfe (Euroscaptor)). Für die japanischen Maulwürfe wies Schwarz mit Talpa micrura wogura eine Unterart aus, die neben dem Japanischen und dem Kleinen Japanischen Maulwurf auch noch den Japanischen Bergmaulwurf (Oreoscaptor) einschloss.[39] Durch die Eingliederung aller talpinen Maulwürfe in die Gattung Talpa war allerdings die von Kuroda geprägte Bezeichnung minor präokkupiert, denn bereits 1914 hatte Wilhelm Freudenberg mit Talpa europaea minor eine fossile Form der Eurasischen Maulwürfe aus dem Mittelpleistozän ausgewiesen.[40] Auf diesen Umstand wurde Kuroda von Yoshinori Imaizumi aufmerksam gemacht. Ihm zu Ehren wählte er den neuen Namen imaizumii.[41]
In den 1960er Jahren wurde die Gattung Mogera teilweise wieder von japanischen Wissenschaftlern verwendet, wodurch sich auch die Originalbezeichnung Mogera wogura minor mehr oder weniger verbreitete. Allerdings kam es zu dieser Zeit zu einer Verwechslung mit der Bezeichnung Mogera wogura für den Japanischen Maulwurf. Ausgangspunkt hierfür ist wiederum Kuroda, der im Jahr 1940 eine zuvor 1905 von Oldfield Thomas getätigte Aussage missinterpretierte. Thomas hatte in dem genannten Jahr die Form Talpa wogura kobeae eingeführt, einen großen Maulwurfsvertreter aus dem südlichen Japan und mit einer Serie kleinerer Maulwurfsschädel aus der Region um Yokohama verglichen, die seiner Meinung nach aber eindeutig zum Japanischen Maulwurf zu stellen waren.[42] Kuroda fasste diese jedoch als zum Kleinen Japanischen Maulwurf gehörig auf. In der Folgezeit wurden daher die kleinen Maulwürfe aus dem nördlichen Japan häufig mit Mogera wogura wogura bezeichnet, die größeren aus dem südlichen Landesteil mit Mogera wogura kobeae. Im Rahmen einer Revision der japanischen Insektenfresser hob Hisashi Abe im Jahr 1967 beide Formen in den Artstatus, wodurch der Kleine Japanische Maulwurf die Bezeichnung Mogera wogura erhielt, der Japanische Maulwurf entsprechend die Bezeichnung Mogera kobeae.[4][9][5] Die Problematik konnte erst in den 1990er Jahren aufgelöst werden. In diesem Zuge klärten Masaharu Motokawa und Hisashi Abe auch auf, dass die Bezeichnung Mogera minor nach den Regularien der ICZN invalid und Mogera imaizumii der heute anerkannte Artname für den Kleinen Japanischen Maulwurf ist.[43]
Es ist uneindeutig, ob der Kleine Japanische Maulwurf Unterarten enthält. Er wird momentan als monotypisch geführt, so unter anderem im achten Band des Standardwerkes Handbook of the Mammals of the World aus dem Jahr 2018.[6] Genetisch bestehen aber laut mehrerer Untersuchungen aus den Jahren 2000 bis 2013 drei phylogeographisch unterscheidbare Linien. Sie verteilen sich auf die pazifische Ostküste, die Westküste am Japanischen Meer und auf das Landesinnere von Honshū. Erstere beiden sind im kompakten Verbreitungsgebiet im Norden anzutreffen, letztere in den zersplitterten Arealen weiter südlich. Alle drei Gruppen weisen relativ große genetische Abstände von 3,6 bis 4,9 % zueinander auf. Die Zentral-Honshū-Gruppe ist in weitere Subpopulationen differenzierbar, dies betrifft die Noto-Halbinsel, die Region Shiga-Kyoto und die Kii-Halbinsel mit genetischen Distanzen von 1,3 bis 1,9 %. Innerhalb der Gruppe der pazifischen Westküste ließ sich nur ein Individuum von der Izu-Halbinsel ganz im Süden des zusammenhängenden nördlichen Vorkommens anhand von Haplotypen und aufgrund einer genetischen Distanz von 2 % unterscheiden. Wahrscheinlich erfolgte die Aufgliederung des Kleinen Japanischen Maulwurfs in die drei Hauptlinien im Unteren Pleistozän vor rund 940.000 bis 740.000 Jahren.[36][44][32]
Bedrohung und Schutz
Der Kleine Japanische Maulwurf ist weit verbreitet und häufig sowie vor allem im nördlichen Teil seines Verbreitungsgebietes der dominante Maulwurfsvertreter. Es sind außerdem keine bestandsgefährdenden Bedrohungen bekannt. Die IUCN stuft ihn daher als „nicht gefährdet“ (least concern) ein. Die Art ist in mehreren Schutzgebieten anwesend.[45]
Literatur
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 615–616) ISBN 978-84-16728-08-4
Einzelnachweise
- Hisashi Abe: Habitat factors affecting the geographic size variation of Japanese moles. Mammal Study 21 (2), 1996, S. 71–87 ()
- Sherman A. Hoslett und Yoshiharu Imaizumi: Seasonal Weight Changes in a Japanese Mole Population. Journal of the Mammalogical Society of Japan 3 (3), 1967, S. 53–56 ()
- Takuma Hashimoto und Manabu Abe: Body size and reproductive schedules in two parapatric moles, Mogera tokudae and Mogera imaizumii, in the Echigo Plain. Mammal Study 26, 2001, S. 35–44 ()
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture, Hokkaido University 55, 1967, S. 191–265 ()
- Hisashi Abe: Revision of the Asian moles of the genus Mogera. Journal of the Mammalogical Society of Japan 20 (1), 1995, S. 51–68 ()
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 615–616) ISBN 978-84-16728-08-4
- Shin-ichiro Kawada, Hideki Endo, Sen-ichi Oda und Kazuhiro Koyasu: Dental Anomalies in Four Mole Species of the Genus Mogera (Insectivora, Talpidae) from Japan. Bulletin of the National Science Museum Series A 37 (1), 2011, S, 63–72 ()
- Hideki Endo, Schu Kawashima, Takao Tsutsumi, Motoki Sasaki, Daishiro Yamagiwa und Yoshihiro Hayashi: Skull Morphology and Mitochondrial DNA Sequence Analysis in the Lesser Japanese Mole (Mogera imaizumii) from the Imperial Palace (Tokyo, Japan). Journal of Veterinary Medical Science 61 (10), 1999, S. 1087–1091 ()
- Kimiyuki Tsuchiya: Cytotaxonomic studies of the Family Talpidae from Japan. Honyurui Kagaku 28, 1988, S. 49–61 ()
- Shin-ichiro Kawada, Masashi Harada, Yoshitaka Obara, Shuji Kobayashi, Kazuhiro Koyasu und Sen-ichi Oda: Karyosystematic Analysis of Japanese Talpine Moles in the Genera Euroscaptor and Mogera (Insectivora, Talpidae). Zoological Science 18, 2001, S. 1003–1010
- Yukako Naitoh: The record of Mogera imaizumii (Kuroda, 1957) in Ohno City, Fukui Prefecture. Fukui City Museum of Natural History 53, 2006, S. 135–136
- Junji Moribe und Yasushi Yokohata: Mixed distribution of Mogera imaizumii and Mogera wogura in the Tedori Alluvial Fan in Ishikawa Prefecture, Japan. Mammal Study 36, 2011, S. 135–139, doi:10.3106/041.036.0303
- Hisashi Abe: Soil hardness, affecting the range expansion of Mogera wogura in Japan. Mammal Study 26, 2001, S. 45–52 ()
- Hisashi Abe: The northeastern front in the distribution of Mogera wogura in the central Honshu, Japan in 2009, especially the past 50 years’ change in Nagano Prefecture. Honyurui Kagaku 50 (1), 2010, S. 55–66 ()
- Yoshiharu Imaizumi und Tadaaki Imaizumi: Interspecific relationship in two mole species in the plains of Niigata, Honshu. 1. Geographic distribution. Journal of the Mammalogical Society of Japan 5, 1970, S. 15–18 ()
- Naohiko Sagara und Yu Fukasawa: Inhabitant changes in long-term mole nesting at the same site, revealed by observing mushroom fruiting at the site. Mammalia 78 (3), 2014, S. 383–391, doi:10.1515/mammalia-2013-0073
- Naohiko Sagara, Hisashi Abe und Hiroaki Okabe: The persistence of moles in nesting at the same site as indicated by mushroom fruiting and nest reconstruction. Canadian Journal of Zoology 71, 1993, S. 1690–1693
- Atsushi Kashimura, Kunihito Moteki, Yosuke Kitamura, Daiju Hayashi, Yuu Shimoyokkaichi, Akio Shinohara, Tetsuo Morita und Kimiyuki Tsuchiya: Subterranean spatial utilization of the lesser Japanese mole, Mogera imaizumii (Kuroda, 1957). Japanese Journal of Environmental Entomology and Zoology 21, 2010, S. 155–164 ()
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). II. Biological aspects. Journal of the Faculty of Agriculture, Hokkaido University 55 (4), 1968, S. 429–458 ()
- Yoshiharu Imaizumi: Hunting Methods in Relation to Hunting Situations in Japanese Shrew-mole, Urotrichus talpoides. II. Detection of the Earthworm "Head". Annotations Zoologicae Japonenses 52 (3), 1979, S. 212–224 ()
- Yoshimasa Kuroi und Masahiro A. Iwasa: Activity of male internal genitalia and breeding season in the lesser Japanese mole, Mogera imaizumii, inferred from morphometric and histological analyses. Mammal Study 41, 2016, S. 25–30, doi:10.3106/041.041.0105
- Shin S. Satoh, Chie Murata, Kenji Yoshida, Kiyotaka Shibata und Hidetoshi B. Tamate: Population genetic study of the lesser Japanese mole Mogera imaizumii using novel microsatellite markers with special reference to sex-biased dispersal. Mammal Study 44, 2019, S. 91–97, doi:10.3106/ms2018-0031
- Sherman A. Hoslett und Yoshiharu Imaizumi: Age Structure of a Japanese Mole Population. Journal of the Mammalogical Society of Japan 2 (6), 1966, S. 151–156 ()
- Takayuki Funo, Shin-ichiro Fujitsuka, Ken Motomura, Mami Oishi, Kazuaka Takahashi, Yoshio Sekiya, M. Horitou, Y. Wakui, H. Kaminushi, Yosuke Hironaka, Tsuneo Sekijima und Manabu Abe: Food habits analysis of Hodgson's hawk eagle, Spizaetus nipalensis by CCD camera observation system in the breeding season. Bulletin of the Faculty of Agriculture. Niigata University (Japan) 53 (1), 2000, S. 71–79
- Hiroshi Kaneda: Prey selection and provisioning rate of a breeding pair of Hodgson’s Hawk-eagles Nisaetus nipalensis. Ornithological Science 8, 2009, S. 151–156 ()
- Yasushi Yokohata, Hisashi Abe, Vue Ping Jiang und Masao Kamiya: Gastrointestinal helminth fauna of Japanese moles, Mogera spp. Japanese Journal of Veterinary Research 37, 1989, S. 1–13 ()
- Tetsuro Koizumi, Sadao Nogami und Yasushi Yokohata: Gastrointestinal Helminth Fauna of the Lesser Japanese Mole (Mogera imaizumii) in Kanagawa Prefecture, Japan, and Analyses on Infection Status of Two Parasitic Nematode Species of the Host. Japanese Journal of Zoo and Wildlife Medicine 16 (2), 2011, S. 121–126()
- Yasushi Yokohata, Yue Ping Jiang, Hisashi Abe und Masashi Ohbayashi: Pseudoparasitism by thelastomatid nematodes in moles, Mogera ssp., in Japan. Japanese Journal of Veterinary Research 36,1988, S. 53–67 ()
- Donald W. Duszynski und Alice R. Wattam: Coccidian Parasites (Apicomplexa: Eimeriidae) from Insectivores. V. Ten Forms from the Moles of Japan (Euroscaptor, Mogera spp.). Journal of Parasitology 35 (1), 1988, S. 55–57
- Noboru Nunomura: A new Species of the Genus Lucasioides (Crustacea: Isopoda) from nest material of Mogera imaizumii, Toyama-ken, Central Japan. Contributions fom the Toyama Science Museum 381, 2010, S. 33–37 ()
- Akio Shinohara, Shin-Ichiro Kawada, Nguyen Truong Son, Chihiro Koshimoto, Hideki Endo, Dang Ngoc Can und Hitoshi Suzuki: Molecular phylogeny of East and Southeast Asian fossorial moles (Lipotyphla, Talpidae). Journal of Mammalogy 95 (3), 2014, S. 455–466
- Takashi Kirihara, Akio Shinohara, Kimiyuki Tsuchiya, Masashi Harada, Alexey P. Kryukov und Hitoshi Suzuki: Spatial and Temporal Aspects of Occurrence of Mogera Species in the Japanese Islands Inferred from Mitochondrial and Nuclear Gene Sequences. Zoological Science 30, 2013, S. 267–281
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- E. D. Zemlemerova, A. A. Bannikova, A. V. Abramov, V. S. Lebedev und V. V. Rozhnov: New Data on Molecular Phylogeny of the East Asian Moles. Doklady Biological Sciences 451, 2013, S. 257–260
- Elena Zemlemerova, Alexey Abramov, Alexey Kryukov, Vladimir Lebedev, Mi-Sook Min, Seo-Jin Lee und Anna Bannikova: Genetic and morphologic diversity of the moles (Talpomorpha, Talpidae, Mogera) from the continental Far East. Journal of Zoological Systematics and Evolutionary Research 57, 2019, S. 662–678, doi:10.1111/jzs.12272
- Kimiyuki Tsuchiya, Hitoshi Suzuki, Akio Shinohara, Masashi Harada, Shigeharu Wakana, Mitsuru Sakaizumi, Sang-Hoon Han, Liang-Kong Lin und Alexei P. Kryukov: Molecular phylogeny of East Asian moles inferred from the sequence variation of the mitochondrial cytochrome b gene. Genes & Genetic Systems 75 (1), 2000, S. 17–24 ()
- Akio Shinohara, Kevin L. Campbell und Hitoshi Suzuki: An evolutionary view on the Japanese talpids based on nucleotide sequences. Mammal Study 30, 2005, S. S19–S24 ()
- Yoshinori Imaizumi: List of type specimens of mammals which are present or have been preserved in the past in Japan (I.). Circular of the Japanese Society of Systematic Zoology 27, 1962, S. 6–10 ()
- Ernst Schwarz: Revision of the Old‐World Moles of the Genus Talpa Linnaeus. Proceedings of the Zoological Society of London 118 (1), 1948, S. 36–48
- Wilhelm Freudenberg: Die Säugetiere des älteren Quartärs von Mitteleuropa mit besonderer Berücksichtigung der Fauna von Hundsheim in Niederösterreich nebst Ausführungen über verwandte Formen aus Mauer, Mosbach, Cromer und anderen Fundorten. Geologische und Paläontologische Abhandlungen N. F. 12, 1914, S. 455–670 (S. 661) ()
- Nagamichi Kuroda: A new name for the lesser Japanese mole. Journal of the Mammalogocal Society of Japan 1 (4), 1957, S. 74 ()
- Oldfield Thomas: On some new japanese mammals presented to the British Museum by Mr. R. Gordon Smith. Annals and Magazine of Natural Histora 7 (15), 1905, S. 487–495 ()
- Masaharu Motokawa und Hisashi Abe: On the specific names of the Japanese moles of the genus Mogera (Insectivora, Talpidae). Mammal Study 21, 1996, S. 115–123 ()
- Masahiro A. Iwasa, Chizu Kawakubo, Kimiyuki Tsuchiya und Hitoshi Suzuki: Intraspecific Differentiation in the Lesser Japanese Mole in Eastern Honshu, Japan, Indicated by Nuclear and Mitochondrial Gene Analyses. Zoological Science 23, 2006, S. 955–961, doi:10.2108/zsj.23.955
- F. Cassola: Mogera imaizumii. The IUCN Red List of Threatened Species 2016. e.T41465A22323581 (); zuletzt aufgerufen am 20. Januar 2021
Weblinks
- Mogera imaizumii in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: F. Cassola, 2016. Abgerufen am 20. Januar 2021.