α-N-Acetylgalactosaminidase
Die α-N-Acetylgalactosaminidase (kurz: Nagalase, EC 3.2.1.49)) ist ein Enzym, das zu den Glycosidasen zählt. Nagalase katalysiert reversibel die Abspaltung von N-Acetylgalactosamin von größeren Molekülen (Glycokonjugate). Im Menschen kodiert das Gen NAGA für die Nagalase.[1]
| α-N-Acetylgalactosaminidase | ||
|---|---|---|
| ||
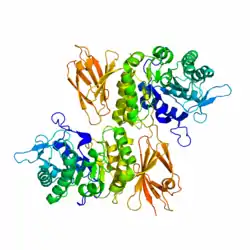
| Bändermodell der menschlichen α-N-Acetylgalactosaminidase, nach PDB 3H53. | ||
| Andere Namen |
| |
|
Vorhandene Strukturdaten: 1KTB, 1KTC, 3H53, 3H54, 3H55, 3IGU, 5WZN, 5WZP, 5WZQ, 2IXB, 5WZR | ||
| Eigenschaften des menschlichen Proteins | ||
| Masse/Länge Primärstruktur | 411 Aminosäuren | |
| Sekundär- bis Quartärstruktur | Homodimer | |
| Bezeichner | ||
| Gen-Namen | NAGA ; GALB; D22S674 | |
| Externe IDs | ||
| Enzymklassifikation | ||
| EC, Kategorie | 3.2.1.49, Hydrolase | |
| Reaktionsart | Hydrolyse | |
| Substrat | endständige (terminale) α-N-Acetylgalactosaminreste | |
| Produkte | N-Acetylgalactosamin + Aglycon | |
| Vorkommen | ||
| Homologie-Familie | HOG000161224 | |
| Übergeordnetes Taxon | Bakterien, Eukaryoten | |
| Orthologe | ||
| Mensch | Hausmaus | |
| Entrez | 4668 | 17939 |
| Ensembl | ENSG00000198951 | ENSMUSG00000022453 |
| UniProt | P17050 | Q9QWR8 |
| Refseq (mRNA) | NM_000262 | NM_008669 |
| Refseq (Protein) | NP_000253 | NP_032695 |
| Genlocus | Chr 22: 42.06 – 42.07 Mb | Chr 15: 82.33 – 82.34 Mb |
| PubMed-Suche | 4668 | 17939 |
Mutationen in NAGA sind sehr selten und führen zu einem α-N-Acetylgalactosaminidase-Mangel mit entsprechendem klinischen Bild: Morbus Schindler und in der adulten Form Morbus Kanzaki. Es wurde postuliert, dass Nagalase von Krebszellen ausgeschüttet werde, die durch Schwächung des Immunsystems die Proliferation der Krebsezellen erleichtern soll.[2][3] Die diesbezüglichen wissenschaftlichen Veröffentlichungen wurden von einigen Zeitschriften wegen erheblichen Qualitätsmängel widerrufen.
Von Impfgegner werden diverse Verschwörungstheorien verbreitet, beispielsweise soll Nagalase in Impfstoffen enthalten sein.[3][2] Dies ist nicht der Fall.
Bakterielle Nagalasen wurden für eine biotechnologische Anwendung vorgeschlagen: Durch enzymatische Abspaltung von N-Acetylgalactosamin können rote Blutkörperchen der Blutgruppe A zu Blutgruppe 0 konvertiert werden.[4] Mittlerweile wird hierfür aber ein anderes, wesentlich effizienteres System mit zwei Enzymen diskutiert.[5]
Struktur
Die menschliche Nagalase setzt sich aus zwei Domänen zusammen. Die Domäne 1 enthält acht α/β-Fässer und die Domäne 2 enthält acht antiparallele β-Stränge in zwei β-Faltblättern. Die Nagalase ist außerdem ein stark glykosyliertes und disulfidreiches Glykoprotein. Das reife Wildtyp-Protein enthält fünf N-gerbundene Glykosylierungsstellen (Asn124, Asn177, Asn201, Asn359 und Asn385), vier Disulfidbrücken (Cys38-Cys80, Cys42-Cys49, Cys127-Cys158 und Cys187-Cys209) und einen freien Cysteinrest (Cys343).
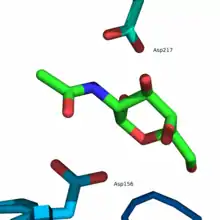
Das aktive Zentrum befindet sich im α/β-Fass am C-terminalen Ende der β-Stränge der Domäne 1. Das aktive Zentrum wird aus Schleifen gebildet, die sich C-terminal zu den sechs aufeinanderfolgenden β-Strängen (β1–β6) befinden. Zu den Aminosäureresten, die das aktive Zentrum bilden, gehören Trp33, Asp78, Asp79, Tyr119, Cys127, Lys154, Asp156, Cys158, Ser188, Ala191, Tyr192, Arg213 und Asp217. Außerdem sorgen Tyr192 und die Disulfidbrücke Cys127-Cys158 dafür, dass ein α-Anomer als Substrat selektiert wird.[6]
Funktion
Konversion von Blutgruppe A zur Blutgruppe 0
Das AB0-System basiert auf das Vorhandensein oder Fehlen der Blutgruppen-Antigene A und B. Die Antigene bestehen aus Kohlenhydratstrukturen, die sich an den Enden von Oligosaccharidketten von Glykoproteinen oder Glykolipiden befinden. Dabei befinden sich die Glykoproteine oder Glykolipide an der Oberfläche von Erythrozyten, sowie von Endothel- und den meisten Epithelzellen.[7][8] Das immundominante Monosaccharid, das die Spezifität der Blutgruppe A bestimmt, ist ein α-1,3-gebundenes N-Acetylgalactosamin, welches terminal an einer Oligosaccharidkette gebunden ist. Zudem werden Antikörper immer gegen die nicht-vorhandenen Antigene gebildet, bei Blutgruppe A also Antikörper gegen B und umgekehrt, bei Blutgruppe AB keine Antikörper und bei Blutgruppe 0 Antikörper gegen A und B. Deshalb können Individuen mit Anti-A (Blutgruppe B)- und/oder Anti-B (Blutgruppe A)-Antikörper keine Bluttransfusion mit inkompatiblen Antigenen erhalten, da es sonst zur Aktivierung des Komplementsystems und schließlich zur Auflösung der Erythrozyten führt, die oftmals eine akute hämolytische Transfusionsreaktion (AHTR) oder andere Reaktionen verursachen.[9][10] Die Erythrozyten der Blutgruppe 0 enthalten weder A- noch B-Antigene, weshalb die Bluttransfusion von Individuen mit Blutgruppe 0 in Trägern aller anderen Blutgruppen erfolgen kann.
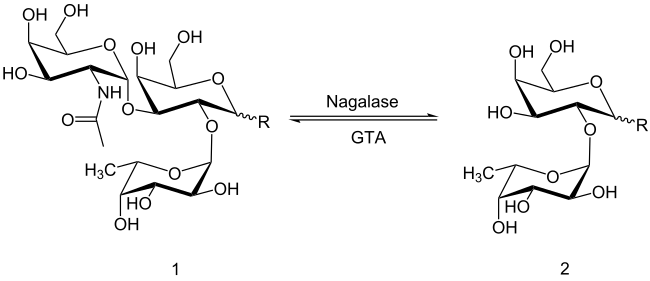
Das immundominante Trisaccharid-Epitop des Antigen A (1) wird aus dem Disaccharid-Epitop (2) mithilfe des Enzyms α-1,3-N-Acetylgalactosaminyltransferase (GTA) gebildet, wobei das Disaccharid-Epitop auch im Antigen H (Blutgruppe 0) vorkommt. Zur Konversion der Blutgruppe A zur Blutgruppe 0 wird eine bakterielle Exoglycosidase eingesetzt, beispielsweise die Nagalase des Bakteriums Elizabethkingia meningoseptica, welches die glykosidische Bindung hydrolysiert und somit die Rückreaktion katalysiert. Aufgrund der Eigenschaft zur Blutgruppen-Konversion von A zu 0 ist die Nagalase auch unter dem Namen A-zym bekannt.[4]
Mechanismus
Der Reaktionsmechanismus am aktiven Zentrum folgt dem sogenannten Ping-Pong-Mechanismus, wobei durch zwei aufeinanderfolgende nukleophile Angriffe am anomeren Zentrum das α-anomere Substrat gespalten wird und man als Produkt ebenfalls ein α-Anomer erhält. Zur Spaltung der glykosidischen Bindung werden zwei Carboxylatgruppen benötigt, wobei eine als Nukleophil und die andere zunächst als Säure und anschließend als Base agiert. Der erste Schritt ist der nukleophile Angriff der Carboxylatgruppe von Asp156 auf das C1-Atom des Substrats, was zur Spaltung der glykosidischen Bindung und zur Freisetzung das Aglycons (R–OH) führt, das anschließend durch Asp217 protoniert wird. Die daraus resultierende Bildung eines hochenergetischen kovalenten Intermediats in Twistkonformation führt zur anschließenden Hydrolyse des Intermediats durch ein Wassermolekül, das vorher durch Asp217 deprotoniert wurde und dadurch ein Hydroxidion erzeugt. Dabei führt das Hydroxidion einen nukleophilen Angriff auf das C1-Atom des kovalenten Intermediats aus, was zur Abspaltung von Asp156 und zur Bildung von N-Acetylgalactosamin als Endprodukt führt.[6]

Weblinks
- Fragen und Antworten zum Nagalase-Test. In: nagalase-test.de. Abgerufen am 15. Januar 2020.
Einzelnachweise
- NAGA alpha-N-acetylgalactosaminidase [Homo sapiens (human)] – Gene – NCBI. Abgerufen am 15. Januar 2020 (englisch).
- Kathrin Helmreich: “Alle Ärzte, die krebsverursachende Enzyme in Impfstoffen fanden, sind tot!” In: mimikama. 11. Januar 2018, abgerufen am 15. Januar 2020.
- Ralf Nowotny: "Krebs-Enzym" in Impfstoffen: Mussten Ärzte deshalb sterben? (Faktencheck). In: mimikama. 9. Januar 2020, abgerufen am 15. Januar 2020.
- Qiyong P. Liu et al.: Bacterial glycosidases for the production of universal red blood cells. In: Nature Biotechnology. Band 25, Nr. 4, April 2007, S. 454–464, doi:10.1038/nbt1298, PMID 17401360.
- Peter Rahfeld und Stephen G. Withers: Toward universal donor blood: Enzymatic conversion of A and B to O type. In: The Journal of Biological Chemistry. Band 295, Nr. 2, 10. Januar 2020, S. 325–334, doi:10.1074/jbc.REV119.008164, PMID 31792054, PMC 6956546 (freier Volltext).
- N. E. Clark, S. C. Garman: The 1.9 a structure of human alpha-N-acetylgalactosaminidase: The molecular basis of Schindler and Kanzaki diseases. In: Journal of molecular biology. Band 393, Nummer 2, Oktober 2009, S. 435–447, doi:10.1016/j.jmb.2009.08.021, PMID 19683538, PMC 2771859 (freier Volltext).
- W. M. Watkins: Biochemistry and Genetics of the ABO, Lewis, and P blood group systems. In: Advances in human genetics. Band 10, 1980, S. 1–136, 379, doi:10.1007/978-1-4615-8288-5_1, PMID 6156588 (Review).
- H. Clausen, S. Hakomori: ABH and related histo-blood group antigens; immunochemical differences in carrier isotypes and their distribution. In: Vox sanguinis. Band 56, Nummer 1, 1989, S. 1–20, doi:10.1111/j.1423-0410.1989.tb03040.x, PMID 2464874 (Review).
- K. Sazama: Transfusion errors: scope of the problem, consequences, and solutions. In: Current hematology reports. Band 2, Nummer 6, November 2003, S. 518–521, PMID 14561397 (Review).
- D. Stainsby, H. Jones, D. Asher, C. Atterbury, A. Boncinelli, L. Brant, C. E. Chapman, K. Davison, R. Gerrard, A. Gray, S. Knowles, E. M. Love, C. Milkins, D. B. McClelland, D. R. Norfolk, K. Soldan, C. Taylor, J. Revill, L. M. Williamson, H. Cohen: Serious hazards of transfusion: a decade of hemovigilance in the UK. In: Transfusion medicine reviews. Band 20, Nummer 4, Oktober 2006, S. 273–282, doi:10.1016/j.tmrv.2006.05.002, PMID 17008165.