Sechspunkt-Jagdspinne
Die Sechspunkt-Jagdspinne (Dolomedes triton) ist eine Spinne aus der Familie der Jagdspinnen (Pisauridae). Die mit einer maximalen Körperlänge von über 30 Millimeter beim Weibchen und demzufolge für Echte Webspinnen (Araneomorphae) vergleichsweise sehr große Art ist großflächig in Nordamerika und vermutlich auch in Mittel- sowie Südamerika vertreten. Der Trivialname deutet auf die drei Paare kleinerer, punktartiger und weißer Flecken auf dem Opisthosoma (Hinterleib) der Spinne, die von weiteren Paaren größerer Flecken umgeben sind. Die im englischen Sprachraum gängige Trivialbezeichnung Six-spotted fishing spider, deren Bedeutung mit der deutschen identisch ist, spielt demzufolge ebenso auf diese Eigenschaft an.
| Sechspunkt-Jagdspinne | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Sechspunkt-Jagdspinne (Dolomedes triton), Weibchen | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Dolomedes triton | ||||||||||||
| (Walckenaer, 1837) |
Die Sechspunkt-Jagdspinne nimmt vor allem stillstehende Gewässer als Habitat (Lebensraum) an und zeichnet sich wie alle Uferjäger (Dolomedes) durch ihre amphibisch geprägte Lebensweise aus. Sie kann sich dank ihres dichten Kleides aus Setae (chitinisierten Haaren) sowohl problemlos auf der Wasseroberfläche fortbewegen als auch untertauchen. Auch der Beutefang geschieht auf Gewässern. Dabei wartet die Sechspunkt-Jagdspinne für längere Zeit reglos, bis im Wasser befindliche Beutetiere in Reichweite gelangen. Ein solches wird dann erbeutet, indem die Spinne blitzartig abtaucht und auf dieses unter Wasser zuschnellt. Damit entspricht die Jagdweise der Sechspunkt-Jagdspinne der Lauerjagd. Das Beutespektrum der Art setzt sich sowohl aus anderen Gliederfüßern als auch aus kleineren Wirbeltieren zusammen.
Der Lebenszyklus der Sechspunkt-Jagdspinne entspricht grundsätzlich dem der Gattung. Ein durch abgesonderte Pheromone (Botenstoffe) des Weibchens angelocktes Männchen übt bei Annäherung ein charakteristisches Balzverhalten aus, wobei es dabei nicht selten zu Kannibalismus seitens des Weibchens kommt. Nach der Paarung legt das Weibchen einen oder mehrere Eikokons an und befestigt diesen in einem für die Familie typischen Brutgespinst. Dort bewacht das Weibchen den Kokon und für kurze Zeit seine Nachkommen nach deren Schlupf. Sobald die Jungtiere sich voneinander und ihrer Mutter trennen, wachsen sie wie alle Spinnen über mehrere Häutungsstadien heran.
Merkmale

Die Sechspunkt-Jagdspinne ist wie viele Arten der Uferjäger (Dolomedes) ein vergleichsweise großer Vertreter der Echten Webspinnen (Araneomorphae) und kann eine Körperlänge von 17 bis 26 (in seltenen Fällen 30) Millimetern beim Weibchen und eine von 9 bis 13 Millimetern beim Männchen erreichen.[1] Der grundsätzliche Körperbau ist mit dem anderer Arten der Gattung identisch.
Das Gewicht der ausgewachsenen Individuen der Sechspunkt-Jagdspinne kann zwischen einem und anderthalb Gramm variieren. Eine bei allen Uferjägern und somit auch bei der Sechspunkt-Jagdspinne vorhandene Eigenschaft ist die dichte Behaarung, die der Spinne eine Fortbewegung auf und unter dem Wasser ermöglicht. Es wird Luft durch dieses Haarkleid festgehalten, die sowohl der Schwimmfähigkeit als auch der Atmung dient.[1]
Sexualdimorphismus
Wie viele Spinnen, so weist auch die Sechspunkt-Jagdspinne einen stark ausgeprägten Sexualdimorphismus (Unterschied der Geschlechter) auf, der sich neben der Gestalt vor allem in der Größe und der Färbung beider Geschlechter bemerkbar macht.
Männchen
.jpg.webp)
Das Prosoma (Vorderkörper) des Männchens besitzt bei einer Körperlänge von 9,52 ± 1,54 Millimetern eine Länge von 4,33 ± 0,6 und eine Breite von 5,15 ± 1,02 Millimetern.[2] Der Carapax (Rückenschild des Prosomas) hat eine graugrüne bis braune Grundfärbung, wobei der Augenbereich dunkler erscheint. Der Clypeus (Abschnitt zwischen dem vorderen Augenpaar und dem Rand des Carapax) hat einen median (mittig) bis submarginal (nahe dem Rand) befindliche hellere Fläche zwischen zwei Grauflächen. Außerdem befindet sich ein weißes, ebenfalls submarginales Band auf dem Carapax. Dieses erstreckt sich von jedem anterolateralen (vorne und seitlich gelegenen) Winkel bis zum hinteren Rand des Carapax und umschließt dessen braunem Zentrum. Auf dem Carapax verlaufen überdies schwarze Lateral- bzw. Seitenbänder. Außerdem erstreckt sich ein gekrümmtes und blasseres Band posterior (hinterseits) von dem Bereich der oberen Seitenaugen. Vor der Fovea (Apodem) befinden sich zwei undeutliche, dunklere und mediane sowie paarig angelegte Flecken. Ein schmaleres, helleres und median angelegtes Band verläuft von nahe der oberen Mittelaugen bis zum hinteren Ende des Carapax. Ein Paar, bestehend aus kurzen und ebenfalls schwarzen Bändern, umschließt das vordere Ende des hellen Medianbands. Außerdem verlaufen von der Fovea aus radiär mehrere dunkle Linien unterschiedlicher Deutlichkeit, die jeweils am Rand der Mittelscheibe des Carapax enden. Die Cheliceren (Kieferklauen) des Männchens sind hell gefärbt und weisen jeweils zwei bis drei dunkle Längslinien und anterior (vorhergehend) lange, helle Setae (chitinisierte Haare) auf. Die Laden (umgebildete Coxen bzw. Hüftglieder der Pedipalpen) sind leicht hellbraun gefärbt. Das Labium (sklerotisierte bzw. gehärtete Platte zwischen den Laden an der Vorderseite des Sternums) besitzt eine rotbraune Grundfärbung, wobei der apikale (an der Spitze gelegene) Rand ebenfalls heller gefärbt ist. Auch das Sternum (Brustschild des Prosomas) erscheint heller und ist mit drei deutlich erkennbaren Paaren dunkler Flecken auf lateraler Fläche versehen.[3]
Die Formel der Beinpaare des Männchens der Sechspunkt-Jagdspinne nach absteigender Länge lautet wie bei vielen Spinnen 4-1-2-3. Alle Beinsegmente erscheinen ventral (unterhalb) blass und sind dorsal (oberhalb) mit unregelmäßigen grauen Längslinien versehen. Die Femora (Schienen) des vierten Beinpaares weisen beim Männchen je einen stacheligen Tuberkel (Hügel) apikal auf der ventralen Seite auf.[4]
Das Opisthosoma (Hinterleib) des Männchens hat eine dunkelrotbraune bis hellbraune Grundfarbe, wobei die Dorsalfläche eher dunkelbraun und beim Herzbereich deutlich heller gefärbt ist. Außerdem befinden sich auf diesem Körperabschnitt spiegelsymmetrisch zur Mittelachse angeordnete Paare von weißen Flecken befinden. Dazu zählen beidseitig neben dem Herzbereich befindliche Paare von anterioren weißen Flecken, deren Anzahl zwei beträgt. Posterior davon sind vier Paare weißer Flecken seitlich zu drei Paaren kleinerer weißer Flecken ausgeprägt. Zwischen diesen Fleckenpaaren verlaufen transversal unklare, dunkle Bänder. Die Flanken des Opisthosomas sind jeweils mit einem unregelmäßigen, weißen und netzförmigen Band versehen, das an den dunklen Bereich der Dorsalseite des Opisthosomas angrenzt. Die wie die anderen Körperregionen an dieser Stelle hellere Ventralseite des Opisthosomas wird durch unregelmäßige dunkle Flecken charakterisiert. Außerdem befinden sich hier median verlaufende Längsbänder. Daneben befinden sich dunkle Seitenbereiche, die anterior beinahe an die Spinnwarzen gelangen.[4]
_(1).jpg.webp) Dorsalansicht eines Männchens
Dorsalansicht eines Männchens_(2).jpg.webp) Frontalansicht eines Männchens
Frontalansicht eines Männchens Lateralansicht eines Männchens
Lateralansicht eines Männchens Rückansicht eines Männchens mit zwei fehlenden Beinen
Rückansicht eines Männchens mit zwei fehlenden Beinen Detailansicht eines Männchens
Detailansicht eines Männchens
Weibchen

Das Prosoma des Weibchens hat bei einer gesamten Körperlänge von 16,63 ± 2,73 Millimetern eine Länge von 7,38 ± 1 und eine Breite von 6,3 ± 0,83 Millimetern.[5] Die Grundfarbe des Carapax des Weibchens ist grünlich grau bis braun. Im Unterschied zum Männchen ist beim Weibchen jedoch ein kleiner Bereich um jedes einzelne Auge dunkler gefärbt. Die weiteren Farbmuster sind ähnlich wie beim Männchen, doch ist beim Weibchen beim Clypeus ein medianer, heller Fleck zwischen den beiden Grauzonen ausgeprägt. Die Marginalbänder sind beim Weibchen außerdem dunkler. Die Cheliceren verfügen beim Weibchen über eine dunkelrotbraune Färbung. Die anteriore Fläche ist beim Weibchen überdies mit langen und unterschiedlich hellen und dunklen Setae bedeckt. Die Laden sind wie beim Männchen rotbraun gefärbt, dabei jedoch apikal heller. Das Labium des Weibchens weist im Gegensatz zu dem vom Männchen eine dunkelrotbraune Grundfarbe auf, ist dafür aber wie beim Männchen am apikalen Rand erhellt. Die Erscheinung des Sternums gleicht der des Sternums vom männlichen Gegenstück.[4]
Anders als beim Männchen haben beim Weibchen die Beine des ersten und des zweiten Paares eine identische Länge. Wie beim Männchen ist dagegen das vierte Beinpaar länger und das dritte kürzer als die anderen. Auch das sonstige Erscheinungsbild der Beine entspricht dem des Männchens mit Ausnahme der beim Weibchen fehlenden Tuberkel.[4]
Auch die Grundfarbe des Opisthosomas ist beim Weibchen wie beim Männchen dunkelrotbraun bis hellbraun. Dorsal ist die Farbe ebenfalls eher dunkelbraun, doch ist beim Weibchen der Herzbereich weniger hell als beim Männchen gefärbt. Posterior der Herzregion sind beim Weibchen wie beim Männchen vier Paare weißer Flecken angelegt, die bei diesem allerdings von einem dunklen Ring umgeben sind. Neben den weißen Flecken befinden sich bei den Weibchen zudem vier weitere Paare kleinerer weißer Flecken auf lateraler Ebene zu den größeren. Das auch beim Weibchen vorhandene Netzband an beiden Flanken des Opisthosomas gleicht dem des Männchens an selbiger Stelle. Ähnlich wie beim Männchen ist die helle Ventralseite des Opisthosomas beim Weibchen mit dunklen Längsbändern bestückt. Diese sind allerdings weniger deutlich ausgeprägt und auch medianer als die des Männchens angelegt. Beim Weibchen besitzt die Ventralseite zudem unregelmäßige dunkle Flecken.[4]
 Dorsalansicht eines Weibchens
Dorsalansicht eines Weibchens Frontalansicht eines Weibchens
Frontalansicht eines Weibchens Lateralansicht eines Weibchens
Lateralansicht eines Weibchens Rückansicht eines Weibchens
Rückansicht eines Weibchens Detailansicht eines Weibchens mit einer anderen Spinne als Beute
Detailansicht eines Weibchens mit einer anderen Spinne als Beute
Genitalmorphologische Merkmale
_(3).jpg.webp)
Die Tibien (Schienen) der Pedipalpen (umgewandelte Extremitäten im Kopfbereich) sind beim Männchen der Sechspunkt-Jagdspinne länger als breit.[2] Ein einzelner Bulbus (männliches Geschlechtsorgan) der Art zeichnet sich innerhalb der Uferjäger (Dolomedes) dadurch aus, dass die apikale (am Ende gelegene) Spitze der Tibiaapophyse (chitinisierter Fortsatz) gerundet und erweitert ist und sich außerdem über das distale (von der Mitte entfernt liegende) Ende der Tibia des Pedipalpus hinaus erstreckt.[6] Daneben gibt es noch eine weitere Apophyse auf der retrolateralen (seitlich rückliegenden) Fläche der Tibia. Diese ist von der Form her lang und gerundet sowie basal schlank und außerdem distal erweitert. Eine dritte Apophyse ist median angelegt. Diese ist schlank in der basalen und breiter in der distalen Hälfte. Außerdem ist diese Apophyse nicht gebogen. Das Verhältnis zwischen der Länge des Cymbiums (erstes Sklerit, bzw. Hartteil des Bulbus) zur Länge der Tibia vom Pedipalpus beträgt 1,6 bis 2,6.[2]
Die Epigyne (weibliches Geschlechtsorgan) der Art wird innerhalb der Gattung dadurch charakterisiert, dass deren posteriorer (vorderer) Medianrand in dorsale (obere) sowie anteriore (vorhergehende) Richtung gekrümmt ist und somit den größten Teil des Kopulationsapparats bedeckt. Durch die eher leichte Pigmentierung können die Kanten undeutlicher erscheinen. Das mediane Septum (Trennwand) ist an den Seiten konvex und am posterioren Ende deutlich nach innen gedreht.[2] Die Befruchtungskanäle sind relativ schmal und im dorsalen Bereich der Epigyne leicht geschlungen, wobei die Kanäle in ihrem Verlauf jeweils drei Schleifen bilden.[4] Außerdem sind die Kanäle vergleichsweise kurz. Die Spermatheken (Samentaschen) sind eher klein und sackförmig und die Befruchtungskanäle lang und gewickelt.[2] In der anterioren Hälfte des dorsalen Bereichs befinden sich Samenklappen.[4] Das Verhältnis zwischen der Höchstlänge der Epigyne und der maximalen Höhe der Samenklappe, gemessen zwischen der apikalen Spitze der Samenklappe und dem hinteren Ende der Epigyne, beläuft sich auf über 1,9.[7]
Jungtiere

Die Jungtiere der Sechspunkt-Jagdspinne sehen den adulten Individuen sehr ähnlich, unterscheiden sich jedoch durch die hellere und grünere Grundfärbung. Die dunklen Ringe und die weißen Flecken auf dem Opisthosoma stechen bei jüngeren Exemplaren deutlich hervor. Individuen in ganz frühen Häutungsstadien weisen ein undeutliches Muster und quer verlaufende Markierungen auf dem Opisthosoma auf.[4]
Geographische Variationen


Die Sechspunkt-Jagdspinne ist vermutlich die am variabelsten auftretende Art aller in der Nearktis vorkommenden Uferjäger (Dolomedes) und weist sowohl geographisch als auch intrapopular (innerhalb der gleichen Populationen) eine Vielzahl an Variationen auf.[8] Dies zeichnet sich auch in der Größe der Spinne aus. Die im Süden vorkommenden Individuen der Art sind im Regelfall größer als die im Norden ihres Verbreitungsgebiets vorkommenden.[9]
Die Populationen der Art im Osten der Vereinigten Staaten unterscheiden sich sowohl in der Ausgeprägtheit als auch in der Verteilung des Farbmusters auf dem gesamten Körper. Insbesondere bei Weibchen scheinen bereits dunkle Musterungen im fortschreitenden Alter noch dunkler zu werden. Während jüngere und hellere Individuen grünlich gebräunt erscheinen, wechselt die Farbgebung bei älter werdenden Exemplaren zu einem dunkleren Braun. Auf der Dorsalseite scheinen die dunklen Ringe um die weißen Flecken den Mittelpunkt der fortschreitenden Verdunklung zu bilden. Dadurch können diese Ringe von schmalen Ringen bis zu breiten und ineinander verschmelzenden dunklen Längsbändern ausgeprägt sein.[9]
Ebenfalls kann bei den im Osten der Vereinigten Staaten vorkommenden Individuen der Sechspunkt-Jagdspinne die Breite der Bänder des Carapax variieren. Das Band kann variieren und durch Zwischenglieder auch geteilt werden. Es kann von mehreren schmalen submarginalen bis zu einem einzelnen breiten Band ausgebildet sein, welches nahezu die seitlichen Ränder des Carapax erreicht und über den Clypeus hinausreicht. Individuen der Sechspunkt-Jagdspinne mit breiteren Bändern auf dem Carapax sind lediglich im Osten des Verbreitungsgebiets der Art zu finden, während sie im Südosten häufiger sind. Von Nordosten nach Westen ändert sich das Erscheinungsbild dieser Bänder, indem sie zunehmend medianer und gerader werden, bis die Spinnen schließlich vermehrt denen der nah verwandten Art Dolomedes striatus ähneln.[9]
Die Ausprägung der Flecken auf dem Sternum scheint deutlich mehr von der geographischen Lage der jeweiligen Population der Sechspunkt-Jagdspinne beeinflusst zu werden. Bei im Südosten des Verbreitungsgebiets der Art vorkommenden Individuen sind diese Flecken deutlich voneinander separiert, während bei den im Nordwesten auftretenden Exemplaren die Flecken mehr oder weniger miteinander verschmelzen und dabei auch lateral angelegte Bänder bilden können. Vermutlich existiert eine stark ausgeprägte Änderung dieser Variation von Süden nach Norden.[9]
Die Tibiaapophyse des Männchens der Sechspunkt-Jagdspinne scheint das einzige genitalmorphologische Merkmal der Art zu sein, das je nach geographischer Lage unterschiedlich ausgebildet ist. Davon ist insbesondere der Scheitelpunkt der Apophyse betroffen, dessen Ausmaß insbesondere von der Gesamtgröße der Apophyse selber beeinflusst wird. Dafür ist wiederum die Körperlänge der Spinne ausschlaggebend, da die am weitläufigsten ausgeprägten Ausdehnungen dieser Merkmale bei größeren Männchen vorhanden sind. Zusätzlich ist insbesondere bei Männchen der im Nordwesten und -osten gelegenen Populationen die Tibiaapophyse derart gedreht, dass diese in einem Winkel von gut 90° zur Ebene der Basis gelegen ist.[9]
Ähnliche Spinnen
Eine der Sechspunkt-Jagdspinne in ihrem Farbmuster sehr ähnliche und teilweise in den gleichen Gebieten vorkommende Art ist die ebenfalls zu den Uferjägern (Dolomedes) zählende Art D. striatus. Beide Arten besitzen weiße Bänder auf dem Carapax und dem Opisthosoma sowie auf letzterem Körperabschnitt zusätzlich weiße Flecken, während das Sternum beider Spinnen mit dunklen Flecken versehen ist. Bei der Sechspunkt-Jagdspinne sind die Bänder auf dem Opisthosoma jedoch dorsaler angelegt, deutlicher erkennbar und einfarbig, während das dunkle Medianband am Rand und neben den weißen Bändern dunkler ist.[10]
Eine weitere der Sechspunkt-Jagdspinne ähnliche Art ist die Gerandete Jagdspinne (D. fimbriatus), die zur gleichen Gattung gehört, jedoch in Europa verbreitet ist.[11] Die Sechspunkt-Jagdspinne wird außerdem gelegentlich mit größeren Wolfsspinnen (Lycosidae) verwechselt, die wie die Raubspinnen (Pisauridae) mitsamt der Sechspunkt-Jagdspinne ebenfalls zur Überfamilie Lycosoidea zählt. Unter den Wolfsspinnen wird die Sechspunkt-Jagdspinne häufig mit den ebenfalls schwimmfähigen, jedoch wesentlich kleineren Piratenspinnen (Pirata) verwechselt.[12]
 Weibchen von Dolomedes striatus
Weibchen von Dolomedes striatus Weibchen der Gerandeten Jagdspinne (D. fimbriatus)
Weibchen der Gerandeten Jagdspinne (D. fimbriatus) Weibliche Wolfsspinne der Art Hogna carolinensis
Weibliche Wolfsspinne der Art Hogna carolinensis Männliche Piratenspinne (Pirata sp.)
Männliche Piratenspinne (Pirata sp.)
Vorkommen
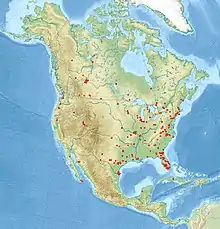
Die Sechspunkt-Jagdspinne kommt auf dem amerikanischen Kontinent vor und besitzt das größte Verbreitungsgebiet aller in der Nearktis vertretenen Uferjäger (Dolomedes). Es reicht nach bisherigen Kenntnissen in Richtung Norden bis zum kanadischen Bundesstaat Ontario und in Richtung Osten bis zum Bundesstaat Maine der Vereinigten Staaten.[13] Einzelne Funde der Art aus dem kanadischen Territorium Nunavut sollen jedoch ebenfalls erfolgt sein.[11] Während das Verbreitungsgebiet der Sechspunkt-Jagdspinne nach Westen bis zum Alaska Panhandle reicht, erstreckt es sich in Richtung Süden über die US-Staaten Florida und Texas, bis es im mexikanischen Bundesstaat Chiapas auf der Halbinsel Yucatán endet.[13] Darüber hinaus soll es auch Sichtungen der Art in Zentral- und Südamerika gegeben haben.[1]
Lebensräume und gemeinsames Auftreten mit anderen Uferjägern

Die Sechspunkt-Jagdspinne lebt wie alle Uferjäger (Dolomedes) gerne in der Nähe von Gewässern und weist dabei von allen nearktischen Arten der Gattung die größte Gebundenheit an stehende Gewässer auf. Sie nimmt als Habitate (Lebensräume) etwa Ränder von Seen oder Teichen,[9] jedoch auch stillstehende Bereiche in Bächen oder Flüssen an.[14] Darüber hinaus ist die Spinne in krautiger Vegetation und gelegentlich auf kleinen Büschen und Bäumen entlang der Wasserlinie anzutreffen.[15]
Die Sechspunkt-Jagdspinne kann innerhalb ihrer Lebensräume neben D. striatus mit weiteren Uferjägern (Dolomedes) gemeinsam angetroffen werden, die ähnliche Ansprüche an ihre Habitate haben. Dabei handelt es sich um die Arten D. albineus, D. okefinokensis und D. tenebrosus, deren Verbreitungsgebiete sich teilweise mit dem der Sechspunkt-Jagdspinne überlappen.[9]
Einflussfaktoren der Häufigkeit
Die Sechspunkt-Jagdspinne gilt in ihrem Verbreitungsgebiet aufgrund ihrer Anpassungsfähigkeit allgemein als häufig. Die Populationsdichte der Art wird jedoch sowohl durch die Anzahl an Prädatoren (Fressfeinden) als auch durch die Verfügbarkeit von geeigneten Beutetieren beeinflusst, was dazu führt, dass die Sechspunkt-Jagdspinne an einigen Gewässern reichlich vorfindbar ist, an anderen jedoch sehr selten vorkommt oder sogar fehlt. Sind größere Fische, die die Rolle als Fressfeinde der Spinne einnehmen, in einem Mikrohabitat nicht vorhanden, wird die Häufigkeit vor allem durch das Nahrungsangebot bestimmt. Je höher dieses im jeweiligen Mikrohabitat ausfällt, desto größer sind dort auch die Populationen der Sechspunkt-Jagdspinne.[16]
Lebensweise

Die Sechspunkt-Jagdspinne ist tagaktiv und verbleibt überwiegend reglos.[1] Wie andere Uferjäger (Dolomedes) hält sich auch diese Art bevorzugt in direkter Wassernähe oder auf der Wasseroberfläche auf. Dabei wird wie bei den anderen Vertretern der Gattung auch der Sechspunkt-Jagdspinne die Fortbewegung auf dem Wasser durch ein dichtes Kleid aus Setae ermöglicht, das der Spinne hydrophobe (wasserabweisende) Eigenschaften und somit einen Auftrieb verleiht. Dadurch kann sich die Spinne auch schnell auf der Wasseroberfläche fortbewegen.[17]
Wird die Sechspunkt-Jagdspinne gestört, taucht sie unter die Wasseroberfläche und versteckt sich dort unter den Blättern von Seerosen oder auf der Vertikalseite des untergetauchten Bereichs von Blättern anderer Pflanzen, die in das Wasser geraten sind.[9] Auch dies wird der Spinne durch ihre dichte Behaarung ermöglicht. Dafür wird Sauerstoff im Haarkleid gespeichert, was der Spinne einen Aufenthalt unter Wasser für eine Dauer von 45 Minuten ermöglicht.[1] Andere Quellen sprechen von anderthalb Stunden maximaler Aufenthaltsdauer unter Wasser.[18]

Auf der Wasseroberfläche neigt die Sechspunkt-Jagdspinne bei starker Luftbewegung zu einem „Segelverhalten“, das daraus besteht, dass die Spinne das zweite Beinpaar anhebt. Damit wird vermutlich die Fähigkeit zum Abfangen von Windbewegungen erhöht, während die basale Auftriebsfläche der Spinne und somit gleichzeitig ihre Stabilität auf dem Wasser unbeeinflusst bleibt. Dadurch spart die Spinne Energie, kann schnell über die Wasseroberfläche gegen die Strömung gleiten und ist gleichzeitig vor wehendem Laub geschützt. Da dies mit minimaler Beinbewegung erlangt wird, wird auch die Bewegung auf dem Wasser reduziert. Dadurch wird ebenso die Erkennbarkeit der Spinne gegenüber Prädatoren verringert, da durch die minimierte Sichtbarkeit auf nicht getarnten Oberflächen insbesondere die Wahrscheinlichkeit, dass Fische und Wasservögel die Spinne erbeuten, verkleinert werden kann.[19]
Jagdverhalten
Die Sechspunkt-Jagdspinne lebt wie alle Spinnen räuberisch und jagt wie alle Raubspinnen (Pisauridae) sowie die meisten Spinnen der Überfamilie der Lycosoidea ohne ein Spinnennetz. Stattdessen nutzt sie, dem Prinzip des Beutefangs der Uferjäger (Dolomedes) entsprechend, das Wasser für den Beuteerwerb. Dabei kann die Sechspunkt-Jagdspinne beim Fang von Beutetieren auf unterschiedliche Methoden zurückgreifen.
Beutefang und Jagderfolg

Zu den Jagdmethoden der Sechspunkt-Jagdspinne zählt unter anderem das stille Verharren an einer geeigneten Position und somit die Methodik der Lauerjagd. Dafür positioniert sich die Spinne kopfüber in der Nähe der Wasserlinie oder hält die vorderen Extremitäten auf die Wasseroberfläche, während die hinteren den Halt der Spinne an einem Blatt oder Zweig sichern.[9] Die Spinne kann über mehrere Stunden in dieser Position verweilen. Alternativ kann die Sechspunkt-Jagdspinne auch aktiv laufend Beute auf der Wasseroberfläche aufsuchen. Daneben ist die Sechspunkt-Jagdspinne dafür bekannt, mit den vorderen Extremitäten auf der Wasseroberfläche zu planschen, um unter Wasser befindliche Beutetiere anzulocken.[1]
Die Sechspunkt-Jagdspinne nutzt dabei wie andere Uferjäger (Dolomedes) auch ihren sensiblen Vibrationssinn und damit ihre Trichobothria (Sinneshaare), um Bewegungen an oder unter der Wasseroberfläche wahrzunehmen. Dabei ist es der Spinne möglich, Beutetiere auf eine Entfernung von bis zu 18 Zentimetern zu orten. Ist ein solches nah genug an die Spinne gelangt, taucht die Spinne für den Fang des Beutetieres sturzartig unter Wasser und kann dabei ebenfalls 18 Zentimeter tief unter die Wasseroberfläche tauchen. Dabei nutzt die Spinne ihren gut entwickelten Sehsinn, um die Beutetiere unter Wasser genau zu orten.[1]
Bei geglückter Jagd wird die Beute dann oberhalb der Wasseroberfläche verzehrt. Allerdings ist dies nur bei gut 9 % der Jagdversuche der Sechspunkt-Jagdspinne der Fall. Die vergleichsweise geringe Erfolgsquote ist wahrscheinlich dem Sehsinn der Spinne verschuldet, der es dieser nicht ermöglicht, Beutetiere genau von anderen im Wasser treibenden Elementen, etwa Blättern oder Zweigen, zu unterscheiden. Solche fehlerhaften Jagdversuche machen 16 % aller Jagdversuche der Sechspunkt-Jagdspinne aus.[1]
Variabilität des Beutespektrums und Menge an Beutetieren
Die Sechspunkt-Jagdspinne weist wie andere Uferjäger (Dolomedes) aufgrund ihrer Größe ein für Spinnen vergleichsweise großes Beutespektrum auf, das sich sowohl aus anderen Wirbellosen, jedoch auch aus kleineren Wirbeltieren zusammensetzt. Überdies kann die Art an einem Tag das Fünffache ihres Eigengewichts zu sich nehmen. Das Beutespektrum der Spinne ist jedoch aufgrund der Jagdmethodik auf unter Wasser oder auf der Wasseroberfläche befindliche Beutetiere beschränkt. Gängige Beutetiere der Sechspunkt-Jagdspinne sind Larven von verschiedenen Insekten, Kaulquappen und kleinere Fische.[1] Wie andere Uferjäger kann die Art theoretisch auch Kröten, Salamander, Eidechsen und Mäuse erbeuten.[20] Besonders häufig werden jedoch neben amphibischen Insekten auch Spinnen mit dieser Lebensweise erbeutet. Zu den geläufigen Beutetieren der Sechspunkt-Jagdspinne zählen außerdem der Nachtfalter Malacosoma disstria aus der Familie der Glucken (Lasiocampidae) sowie Zwergzikaden (Cicadellidae) aus den Gattungen Draeculacephala und Oncometopia. Diese Insekten ernähren sich von Pflanzen, die an den Ufern von Teichen wachsen, weshalb Begegnungen dieser Tiere mit der Sechspunkt-Jagdspinne häufig stattfinden können.[21]

Die Sechspunkt-Jagdspinne wird, anders als viele andere Spinnen, nicht durch hartschalige Exoskelette oder Abwehrsekrete von verschiedenen Insekten abgeschreckt, was es der Art ermöglicht, auch regelmäßig wehrhafte amphibische Vertreter dieser Klasse, etwa Schwimmkäfer (Dytiscidae), Wasserläufer (Gerridae) oder Rückenschwimmer (Notonectidae), zu erbeuten.[21] Daneben ist die Sechspunkt-Jagdspinne auch dafür bekannt, die Imagines (ausgewachsene Stadien) von Libellen und anderen an Gewässer gebundenen Insekten zu erbeuten.[9] Kleinlibellen etwa werden oftmals während der Eiablage oder generell während des Landens von der Spinne gefangen. Aufgrund dessen wird vermutet, dass die Sechspunkt-Jagdspinne vor allem aktive auf der Wasseroberfläche befindliche Beutetiere erlegt, was sie auch etwa von Wasserläufern unterscheidet, die überwiegend verendete Insekten auf der Wasseroberfläche jagen oder solche, die kurz vor dem Tod stehen.[21]

Im Allgemeinen sind Individuen der Art beim Verzehr von Beutetieren eher selten zu finden. Die Verfügbarkeit an Beutetieren wird für die Spinne anscheinend von saisonalen Schwankungen beeinflusst. Teilweise und gänzlich amphibische Wanzen stellen insbesondere während der eisfreien Winterzeit eine ausreichend verfügbare Nahrungsquelle dar, während etwa verschiedene Libellen, Zweiflügler, Käfer und auch andere Spinnen durch deren Phänologie (Aktivitätszeit) starke Häufigkeitsschwankungen zeigen und dadurch als Nahrung für die Sechspunkt-Jagdspinne teilweise nur sporadisch verfügbar sind. Die saisonalen Schwankungen der Verfügbarkeit von Nahrung der Spinne werden zumeist durch die eintretenden Veränderungen der Häufigkeit von verschiedenen Beutetieren erklärt. Eine andere Theorie ist, dass durch Jagderfolge der Spinne selbst die Häufigkeit einzelner Beutetiere beeinflusst wird.[21]

Die Größe der Spinne selbst ist weniger ausschlaggebend für die mögliche Größe, die ein zu erlegendes Beutetier aufweisen kann. Obwohl die Größe der Beutetiere von Jungtieren wesentlich kleiner ausfällt als die der ausgewachsenen Spinnen und insbesondere der Weibchen, so wurden auch kleinere Exemplare der Art gesichtet, die vergleichsweise große Beutetiere erlegt hatten. Das lässt vermuten, dass auch die kleineren Individuen der Sechspunkt-Jagdspinne nicht unbedingt auf kleinere Beutetiere angewiesen sind. Im Gegensatz dazu scheinen ausgewachsene Weibchen sehr kleine Tiere zu meiden, wofür es zwei sich nicht gegenseitig ausschließende Theorien gibt. Eine besagt, dass diese kleinen Tiere zu geringe Vibrationen erzeugen, als dass diese den Jagdtrieb des Weibchens auslösen. Die andere besagt, dass die weiblichen Spinnen diese Lebewesen zwar wahrnehmen, aber das Erlegen für sie in diesem Fall keinen Nutzen erbringen würde, da durch diese der Nahrungsbedarf nicht gedeckt werden könnte. Da die verfügbare Energie gerade bei verpaarten Weibchen durch die Produktion der Eier begrenzt zu sein scheint, müssen diese ihren Nahrungsbedarf umso mehr decken, dürfen dabei jedoch nicht zu viel Energie aufbringen. Die Spinnen versuchen dann in dem Fall eine maximale Nahrungsausbeute mit minimalem Aufwand zu erreichen.[21]
Auswirkung der Sechspunkt-Jagdspinne auf die Bestände ihrer Beutetiere
.jpg.webp)
Die Jagdunternehmungen der Sechspunkt-Jagdspinne können insbesondere durch die saisonal variierende Dichte sowohl von Jungtieren als auch ausgewachsenen Individuen der Art die Menge einzelner Taxa (Gruppen von Lebewesen) an Beutetieren verschieden beeinflussen. Insbesondere Wasserläufer und darunter etwa die Art Limnoporus dissortis sowie insbesondere Gerris buenoi scheinen unter den Wanzen den Großteil der Beute der Sechspunkt-Jagdspinne auszumachen, was dazu führt, dass deren Anzahl in Gebieten mit Populationen der Spinne drastisch reduziert werden kann. Letztere Art wird vermutlich entweder selektiv von der Sechspunkt-Jagdspinne erbeutet oder ist besonders anfällig für deren Jagdversuche. Insbesondere fallen viele Nymphen (in Jugendstadien befindliche Individuen) von G. buenoi der Sechspunkt-Jagdspinne zum Opfer. Sie werden im Spätsommer vermehrt von Jungtieren der Art erbeutet, wodurch die Häufigkeit der Nymphen in einer kurzen Zeitspanne drastisch reduziert werden kann. Demnach können Jagden der Sechspunkt-Jagdspinne die allgemeine Häufigkeit von den genannten Wasserläuferarten in Habitaten, in denen die Wasserläufer und die Spinne gemeinsam vorkommen, saisonal deutlich beeinflussen. Sollten die Bestände der Wasserläufer knapp werden, wechselt die Sechspunkt-Jagdspinne zu einer anderen primären Nahrungsquelle, etwa Ruderwanzen (Corixidae). Auch sind Libellen, besonders beim Landen und deren Weibchen, besonders wenn sie sich bei der Eiablage befinden, durch die Sechspunkt-Jagdspinne gefährdet. Ebenso sind männliche Libellen, die die Weibchen während der Eiablage begleiten, häufige Beutetiere der Art. Insgesamt kann die Sechspunkt-Jagdspinne durch die erläuterten Aspekte auch die Phänologie und die Brauchbarkeit von Lebensräumen verschiedener Beutetiere beeinflussen.[21]
Alle vorherigen Punkte machen eine Räuber-Beute-Beziehung zwischen der Sechspunkt-Jagdspinne und den erwähnten Vertretern der Wanzen deutlich. Diese leben und entwickeln sich gemeinsam mit der Spinne in den gleichen Habitaten, wobei die unterschiedliche Phänologien und Lebensraumnutzungen der Tiere adaptive Reaktionen auf räumlich-zeitliche Variationen gegenseitiger Erbeutungen sein können. Insbesondere der Lebenszyklus der Sechspunkt-Jagdspinne deutet auf eine derartige Relation. Jungtiere der Art, die sich im ersten Häutungsstadium befinden, bewegen sich nur selten aktiv auf der Wasseroberfläche fort, wenn größere Jungtiere der Spinne und größere Nymphen von Wasserläufern sowie Ruderschwimmern maximale Häufigkeiten aufweisen. Kleine Individuen der Sechspunkt-Jagdspinne besiedeln Oberflächen von Gewässern überwiegend ab Mitte Juni und damit ab dem Zeitpunkt, ab dem die Anzahl größerer Exemplare der Spinnen deutlich zurückgegangen ist und Vertreter beider zuvor genannten Wanzenfamilien lediglich in Form von sich entwickelnden Jungtieren vorhanden sind. Die Nahrungskonkurrenz dieser Tiere kann sich demnach gravierend auf deren Koevolution in Gebieten, in denen sie gemeinsam vorkommen, auswirken.[22]
Neigung zu Kannibalismus und seine Auswirkungen
Kannibalismus ist bei der Sechspunkt-Jagdspinne zur Erhaltung eines Gleichgewichts der Populationen wie auch bei anderen Spinnentieren von großer Bedeutung. Dabei tritt dies insbesondere seitens größerer Jungtiere auf, was für die Entwicklung stadienspezifischer Muster der Nutzung von Lebensräumen aus selektiver Sicht wichtig sein kann. Kleinere Individuen der Art meiden die unmittelbare Uferzone sowohl in der Spätsaison als auch im Frühling und halten sich stattdessen mehrere Meter vom Wasser entfernt auf.[21]
Ausgewachsene Weibchen zeigen insbesondere während der Paarungszeit eine höhere Neigung zu kannibalistischem Verhalten und erbeuten dabei häufig Männchen. Sie sind somit ein beeinflussender Faktor für deren Häufigkeit. Ein Kannibalismus nach der Paarung seitens des Weibchens gegenüber dem Männchen, wie er bei einigen anderen Spinnen vorkommen kann, ist im Falle der Sechspunkt-Jagdspinne jedoch unwahrscheinlich, da das Männchen nach der Paarung zumeist vor dem Weibchen flieht. Ferner ist es durch den hohen Nährstoffanteil, den das Weibchen für das Anfertigen seiner Eikokons benötigt, unwahrscheinlich, dass ein Männchen seine reproduktive Fitness durch eine derartige Selbstaufopferung steigern kann.[22]
Lebenszyklus und Phänologie
Der Lebenszyklus der Sechspunkt-Jagdspinne wird wie bei anderen in den gemäßigten Klimazonen vorkommenden Spinnen von den Jahreszeiten maßgeblich beeinflusst. Ausgewachsene Individuen beider Geschlechter der Art sind anscheinend das ganze Jahr über anzutreffen, wobei diese im Norden ihres Verbreitungsgebiets vor allem in den wärmeren Jahreszeiten vorfindbar sind. In Florida können adulte Exemplare der Sechspunkt-Jagdspinne in der Zeitspanne zwischen Dezember und April sogar massenhaft auftreten.[9]
Im Zentrum der kanadischen Provinz Alberta hingegen ähnelt der Lebenszyklus dem der gattungsverwandten Gerandeten Jagdspinne (D. fimbriatus) in Mittel- und Nordeuropa. Dadurch ergibt sich, dass die Sechspunkt-Jagdspinne flexible Regulationen bezüglich ihres Lebenszyklus aufweist. Die bei der Art auftretende Dormanz in Form von Überwinterungsperioden ist nicht zeitlich festgelegt, sodass die Sechspunkt-Jagdspinne unter passenden Bedingungen auch in den nördlichen Gebieten ihres Verbreitungsgebiets in Form von univoltinen (einjährigen) Generationen erscheinen kann. In Alberta sind ausgewachsene Männchen der Art nur innerhalb einer kurzen Zeitspanne zur Reproduktion fähig, während die Anzahl adulter Weibchen für gewöhnlich Mitte Juni ihren Höchstwert erlangt.[23]
Präkopuläres Verhalten
Der eigentlichen Paarung geht ein sogenanntes präkopuläres Verhalten voraus, das sich vor allem aus dem Zusammenfinden beider Geschlechtspartner und der darauf folgenden Balz zusammensetzt.
Auffinden eines Weibchens mittels von diesem abgesonderten Pheromonen
Ein geschlechtsreifes Männchen sucht aktiv ein ebenso geschlechtsreifes Weibchen auf. Das Aufsuchverhalten des Männchens lässt sich in drei Phasen gliedern, bei denen es sich um die Bewegungs-, die sensorische und die eigentliche Balzphase handelt. Die Bewegungsphase, in der das Männchen ein Weibchen aktiv aufsucht, dient dazu, die Wahrscheinlichkeit eines Zusammentreffens beider Geschlechtspartner zu erhöhen, da dem Männchen die Suche nach Weibchen durch die eher unüberschaubaren Habitate der Sechspunkt-Jagdspinne erschwert wird. Außerdem befindet sich das Weibchen der Art nie an einer festen Position, obgleich es eher ortsgebunden ist. Ein Männchen der Art zeigt dabei kein Territorialverhalten gegenüber anderen Männchen.[24]
Ein paarungswilliges Weibchen nutzt arteigene Pheromone, die es über das Integument (äußere Körperhülle) aussondert, um geschlechtsreife Männchen anzulocken. Diese Pheromone breiten sich aufgrund dessen vermutlich auch auf der Wasseroberfläche aus, wie es ebenso bei einer un- oder nur schwach polaren Verbindung, etwa Lipiden oder Steroiden, der Fall ist.[25] Jedoch kann das Männchen der Art auch den Wegfäden von Weibchen sowohl auf trockenem Bodensubstrat als auch auf der Wasseroberfläche folgen. Deshalb wird als dritte Theorie bezüglich der Verbindungen in Betracht gezogen, dass es sich bei dieser um eine unpolare Bindung handelt. Ob und inwiefern die Wirkung der Pheromone abhängig von deren Platzierung variiert, ist noch ungeklärt. Ein Männchen kann jedoch Wegfäden von Weibchen auch auf der Wasseroberfläche folgen, indem es sich an dem Faden entlang zieht und sich durch Ruderbewegungen dabei fortbewegt. Es kann gelegentlich vorkommen, dass Männchen der Sechspunkt-Jagdspinne den Wegfäden anderer Männchen, eines subadulten Weibchens oder eines Weibchens anderer Uferjäger (Dolomedes) folgen, dabei jedoch keinerlei sexuelle Reaktion zeigen.[26]
Da das Weibchen der Sechspunkt-Jagdspinne mitunter über mehrere Tage seine Position nicht wechselt und an einer gleichen Stelle auch gerne für mindestens zwei Stunden reglos verbleibt, besteht die Möglichkeit, dass sich im Wasser um das Weibchen herum die von diesem abgesonderten Pheromone ansammeln. Dadurch ist es einem auf der Suche nach einer Geschlechtspartnerin befindlichen Männchen möglich, dieses bereits zu orten, bevor es dessen Wegfaden gefunden oder visuellen Kontakt mit ihm aufgenommen hat.[25]
Bei der zweiten Phase des Aufsuchprozesses des Männchens, der sensorischen Phase, peilt das Männchen ein Weibchen direkt an. Beide Phasen werden vom Männchen oftmals simultan ausgeführt. Das auf der Wasseroberfläche suchende Männchen kann durch die vom Weibchen abgesonderten Pheromone erkennen, ob es sich in seiner Nähe befindet. Dies lässt darauf schließen, dass das Männchen in der sensorischen Phase zielorientierter nach Geschlechtspartnern sucht, da es nicht einfach auf diese stößt.[24]
Balz
Ist es einem Weibchen gelungen, ein Männchen anzulocken, vollführt letzteres, wie bei anderen der Überfamilie der Lycosoidea zugehörigen Spinnenarten üblich, ein charakteristisches Balzverhalten. Die Balz wird als dritte Phase des Aufsuchverhaltens des Männchens klassifiziert und dient der Identifizierung des Männchens als arteigener Geschlechtspartner gegenüber dem Weibchen, sodass dieses dadurch nicht als Beute in Erscheinung treten soll.[24] Das Männchen der Sechspunkt-Jagdspinne umwirbt dabei sowohl nicht begattete als auch bereits zuvor verpaarte Weibchen.[22] Letzteres erscheint eigenartig, da das Männchen, sollte es zwischen begatteten und unbegatteten Weibchen differenzieren können, einen selektiven Vorteil genießen würde.[27]
Das Balzverhalten ähnelt stark dem Suchverhalten des Männchens und ist kein einzelner Teil für sich, sondern ein Bestandteil des zusammenhängenden Reproduktionsverhaltens, weshalb Ähnlichkeiten zwischen den Verhaltensweisen vorkommen. Dies ist von Vorteil, da ein bereits vorhandenes Verhalten an ein folgendes angleichbar ist und nicht ein gänzlich neues entwickelt werden müsste. Die Anzahl der Ähnlichkeiten von Verhaltensweisen eines einzelnen Aspekts der Fortpflanzung zu einem anderen wird zu einem großen Teil durch die Funktion des jeweiligen Verhaltens bestimmt.[28]
Die Balz besteht unter anderem aus dem schon beim Suchverhalten des Männchens beginnenden Klopfbewegungen auf der Wasseroberfläche. Diese Bewegungen, die dann während der eigentlichen Balz schneller werden, sind ein Beweis für geringfügige Verhaltensänderungen. Klopft das Männchen auf den Körper des Weibchens, kräuselt letzteres seine Beine, was auch bei der ebenfalls zu den Raubspinnen (Pisauridae) zählenden Art Pisaurina mira vorkommt. Dieser Prozess führt außerdem zu einem an Katalepsie oder Starrsucht erinnernden Zustand des Weibchens, das dann auf andere Reize keine Reaktionen zeigt.[28]
Daneben beinhaltet die Balz ein Einspinnen des Weibchens seitens des Männchens, das jedoch anscheinend weniger der Immobilisierung des Weibchens, sondern vielmehr ebenfalls dem Erreichen von dessen katalepsieartigem Zustand dient. Das Männchen, das fast halb so klein wie das Weibchen ist, befestigt an diesem nur selten genug Spinnseide, die das Weibchen an einer Befreiung aus dem Gespinst hindern. Oftmals werden nur drei bis vier Seidenfäden verlegt oder vom Männchen auch lediglich die für das Einspinnen notwendigen Bewegungen ausgeführt. Aufgrund dessen wird vermutet, dass sich der Prozess des Einspinnens im Laufe der Evolution zu einem ritualartigen Paarungsverhalten entwickelt hat, das das Weibchen zusätzlich in den katalepsieartigen Zustand versetzen soll. Ein ähnliches Verhalten ist anscheinend bei der zu den Kräuselspinnen (Dictynidae) zählenden Art Dictyna volucripes vorhanden, bei der das Männchen ebenfalls während der Balz sein Opisthosoma bewegt, während es über das Weibchen läuft. Dies deutet darauf hin, dass auch das Männchen dieser Art das Weibchen bei der Balz einspinnt. Ähnlich wie bei der Sechspunkt-Jagdspinne handelt es sich wohl auch im Falle von D. volucripes bei diesem Verhalten um ein ritualartiges Verhalten und weniger um eine Prävention des Männchen vor Kannibalismus seitens des Weibchens.[29]
Zum Zwecke der Balz führt das Männchen außerdem ruckartige Bewegungen mit dem ganzen Körper aus, wobei es sich dabei um eine Folge sexueller Erregung seitens des Männchens handelt, was sich anhand der dadurch ausgelösten konzentrischen Wellen der Wasseroberfläche bemerkbar macht.[30] In Gefangenschaft ließ sich überdies nachweisen, dass Männchen der Art das Vibrieren auch ohne durch Weibchen ausgelöste Reize ausübt.[31] Sollte die Konzentration der Pheromone höher sein, dann gehen die Balzbewegungen des Männchens in ein rapides Klopfen auf die Wasseroberfläche über. Dies ist insbesondere dann der Fall, wenn die Pheromone vom Männchen nahe der Ruheposition des Weibchens registriert werden.[30] Das Ruckeln seitens des Männchens ist also auch beim Suchverhalten des Männchens präsent und wird überdies auch dann ausgeführt, sollte sich das Männchen auf dem Weibchen befinden. In den ruckartigen Balzbewegungen werden mehrere Zwecke vermutet. Dazu zählen unter anderem die Identifizierung seitens des Männchens gegenüber dem Weibchen als arteigener Geschlechtspartner oder die Demonstration seines Zustands. In Anbetracht dessen, dass das Ruckeln zwischen den Klopfphasen ausgeführt wird, besteht die Theorie, dass ersteres ebenfalls dazu dient, das Weibchen in den katalepsieartigen Zustand zu versetzen.[32]
Während der Balz winkt das Männchen außerdem mit den Beinen.[30] Erwidert das Weibchen die Paarungswilligkeit des Männchens, signalisiert es seine Bereitschaft durch ein Winken der beiden vorderen Beinpaare. Daraufhin üben beide Partner mehrere lang anhaltende Runden der Beinbewegungen aus.[30] Es ließ sich mehrfach nachweisen, dass sich Männchen der Sechspunkt-Jagdspinnen auf andere balzende Männchen zubewegten, was insbesondere dann der Fall war, wenn letztere das ruckartige Bewegungsmuster vollführten.[24]
Das Balzverhalten der Sechspunkt-Jagdspinne kann dabei nach bisherigen Beobachtungen stark variieren. Es wird vermutet, dass einige für die Arterkennung notwendigen Elemente der Balz nicht erforderlich sind, wenn in einem von der Sechspunkt-Jagdspinne bewohnten Mikrohabitat neben dieser keine weitere Art der Uferjäger vorhanden ist.[32] Das gesamte präkopuläre Verhalten mitsamt der Balz wird den erläuterten Aspekten nach durch chemische Reize ausgelöst und beinhaltet außerdem vibratorische und taktische Signale, wobei die Beinbewegungen womöglich auch optischen Reizen dienen. Das präkopuläre Verhalten der Sechspunkt-Jagdspinne ähnelt dem der ebenfalls zu den Uferjägern (Dolomedes) zählenden Art D. scriptus, bei der jedoch das Männchen währenddessen keine ruckartigen Bewegungen mit dem Körper ausführt. Die durch das Beinwinken ausgeübten Signale der Spinne ähneln außerdem denen von Wasserläufern.[30]
Verstärkte Balz bei frisch gehäuteten Weibchen und Exuvien und Theorien zur Kohabitation beider Geschlechter
Das Männchen der Sechspunkt-Jagdspinne zeigt deutlich aktivere Balzsignale gegenüber frisch gehäuteten Weibchen oder solchen, die kurz vor der Häutung stehen. Dies beinhaltet Bewegungen in Form von vermehrten Klopf-, Streich- und Ruckperioden. Darüber hinaus reagiert das Männchen intensiver auf Exuvien (nach der Häutung abgeworfene Exoskelette bzw. Chitinpanzer) von sub- oder präsubadulten Weibchen sowie anderen subadulten Männchen. Der Anreiz für dieses Verhalten dürfte auf das Wahrnehmen von einer mit der Häutung in Verbindung stehenden Chemikalie zurückzuführen sein. Dies dürfte auch erklären, dass das Männchen die Suche scheinbar dann intensiviert, wenn es diese Chemikalie wahrnimmt. Das Männchen könnte somit also frisch gehäutete Weibchen lokalisieren. Ein nach einem Weibchen suchendes Männchen übt ein Suchverhalten generell jedoch auch ohne einen Anreiz dafür aus.[31]
Obwohl Beobachtungen belegen, dass das Männchen der Sechspunkt-Jagdspinne mit dem Weibchen in einer Assoziation steht, lässt sich nicht genau bestimmen, ob es sich bei diesem gemeinsamen Vorkommen um ein direktes Zusammenleben handelt. Erschwert wird die Bildung von befürwortenden Thesen durch das aktive Verhalten des Weibchens der Art bis kurz vor dessen Häutung zum adulten Stadium. Die Verbindung beider Geschlechter scheint der anderer Spinnen der Teilordnung der Entelegynae zu entsprechen. Ein Männchen sucht also die Nähe von Weibchen auf und zeigt bei dieser Art zusätzlich die bereits erwähnte und erhöhte Anziehung auf Weibchen, die sich kurz vor der Häutung befinden. Die Verbindung von subadulten Individuen beider Geschlechter zueinander ist unbekannt.[33]
Kannibalismus seitens des Weibchens bei der Balz und Verhaltensänderung des Weibchens nach der Paarung
Aufgrund der hohen Bereitschaft des Weibchens der Sechspunkt-Jagdspinne zum Kannibalismus fällt ein sich annäherndes Männchen der Art nicht selten dem Weibchen zum Opfer. Dies ist insbesondere dann der Fall, wenn das Weibchen hungrig ist oder das Männchen seine Balzbewegungen falsch ausführt.[1] Dabei lassen sich allerdings seitens des Weibchens Verhaltensunterschiede feststellen, die bei einer Jagd auf andere Beuteobjekte nicht vorkommen. Anstelle direkt auf die Vibrationsquelle zuzuschnellen, wie es ansonsten bei der Jagd der Fall ist, wartet das Weibchen zumeist, bis das Männchen sich in der Nähe befindet. Dieses Verhalten, das vor allem bei zuvor begatteten Weibchen vorkommt, ließ sich auch bei der zur gleichen Gattung zählenden Gerandeten Jagdspinne (D. fimbriatus) belegen. Eine Möglichkeit wäre, dass das Weibchen, um sich einen Jagdvorteil zu verschaffen, eine Unentschlossenheit gegenüber dem Männchen vortäuscht.[22] Das Männchen der Sechspunkt-Jagdspinne kann anscheinend nicht ein bereits begattetes von einem unbegatteten Weibchen unterscheiden, sodass es jedes Weibchen als möglichen Geschlechtspartner betrachtet und sich diesem annähert sowie es umwirbt. Eine Primärfunktion der Balz des Männchens als Prävention gegen Kannibalismus seitens des Weibchens wäre demnach ausgeschlossen.[27]
Es ist belegt, dass das Weibchen der Sechspunkt-Jagdspinne dazu in der Lage ist, balzende Männchen von Beutetieren zu unterscheiden, was dazu dienen soll, dass die Männchen als Geschlechtspartner und nicht als Beuteobjekte betrachtet werden. Da sich das Verhalten des Weibchens gegenüber balzenden Männchen nach der Paarung dennoch ändert, lässt sich vermuten, dass das Weibchen besagte Fähigkeit in diesem Falle nutzt, um die Männchen als zusätzliche Nahrungsquelle nutzen zu können. Dies ließe sich damit erklären, dass sich ein verpaartes Weibchen oft auf ein balzendes Männchen fokussiert, jedoch wartet, bis es sich nähert und dann schlussendlich überwältigt.[34]
Es ist durchaus möglich, dass die eigentliche Begattung bei dem Weibchen in einer Unempfänglichkeit resultiert, die wiederum mit kannibalistischem Verhalten gegenüber Männchen korreliert. Mögliche Bildungen dieser Unempfänglichkeit können entweder durch eine chemische Übertragung von Männchen auf Weibchen gewährleistet sein oder durch das Hervorrufen taktiler Reize ermöglicht werden. Unabhängig von der Entstehung der Unempfänglichkeit wird dem Weibchen somit die Nutzung von Männchen als weitere Nahrungsquelle gesichert.[27]
Eine weitere Möglichkeit wäre, dass ein Weibchen durch sein kannibalistisches Verhalten gegenüber den Männchen auch die Anzahl potentieller Partner für andere Weibchen verringert und somit diese an einer Konkurrenz hindert. Nach bisherigen Kenntnissen kann die arteigene Ausbeute eines Weibchens der Sechspunkt-Jagdspinne zu 37 % aus ebenfalls weiblichen Artgenossen bestehen. Auch damit verringert das Weibchen das Auftreten von konkurrierenden Nachkommen.[35]
Der bei der Sechspunkt-Jagdspinne gelegentlich auftretende Kannibalismus spielt vermutlich auch beim Reproduktionserfolg eine entscheidende Rolle. Da sexueller Kannibalismus bei der Art während der Paarung jedoch selten ist und zumeist Männchen von Weibchen erbeutet werden, die bereits begattet wurden, erbringt das Männchen der Sechspunkt-Jagdspinne für das Weibchen keinen Nutzen durch Selbstaufopferung zwecks einer gesteigerten Reproduktionsrate. Dies wäre als Voraussetzung für einen nutzvollen Kannibalismus vonnöten. Bedingt durch das versetzte Auftreten beider Geschlechter ist es jedoch möglich, dass ein Weibchen der Art mögliche Konkurrenz sowohl für sich selbst als auch für seine Nachkommen auf längere Sicht dezimiert, da es dadurch die Wahrscheinlichkeit verringert, dass später auftretende Weibchen begattet werden können.[36]
Paarung
Die eigentliche Paarung verläuft in der für freijagende Echte Webspinnen (Araneamorpahe) typischen Position III, die hier jedoch wahrscheinlich zwecks der auf der Wasseroberfläche stattfindenden Kopulation abgewandelt ausgeführt wird. Für gewöhnlich besteigt das Männchen bei dieser Position das Weibchen frontal, sodass beide übereinander befindliche Geschlechtspartner jeweils in die entgegengesetzten Richtung blicken. Außerdem würde das Männchen in diesem Falle je einen Bulbus auf einer Seite in die Epigyne des Weibchens einführen, indem es dessen Opisthosoma auf gleicher Seite mit dem jeweiligen Bulbus dafür anhebt. Mangels fester Stützungsmöglichkeiten ist dies im Falle der Paarung bei der Sechspunkt-Jagdspinne nicht möglich und das Weibchen beginnt während der Kopulation auf der Wasseroberfläche zu rollen. Auf ähnliche Art würde auch das Männchen in diesem Falle den Bodengrund nutzen, um sich während seiner Begattungsmethodik zu verfestigen. Die Tuberkel an den Femoren des vierten Beinpaares beim Männchen und der Fakt, dass das Weibchen in dem katalepsieartigen Zustand verbleibt, hilft dem Männchen, das Weibchen auf den Rücken zu legen, was bei dieser Art für die Begattung notwendig ist.[37]
Die verlängerte Tibiaapophyse des Männchens dient vermutlich selbigem Zweck. Bedingt durch die amphibische Biologie der Sechspunkt-Jagdspinne kann das Weibchen der Art eine sehr versiegelte Epigyne aufweisen. Demnach müsste das Männchen ein Organ aufweisen, dass ihm einen Zugang zum Genitaltrakt des Weibchens verschafft, was durch die Tibiaapophyse ermöglicht werden könnte. Die Apophyse würde dieser Annahme nach dazu dienen, den Embolus zwischen sich selbst und der Basis des Cymbiums der Palpe in den Genitaltrakt des Weibchens zu leiten.[38]
Dass das Weibchen sich im Gegensatz zu Männchen nur einmalig verpaart, liegt wahrscheinlich daran, dass das Weibchen schon bei einer einzelnen Kopulation die notwendige Menge an Sperma für das Befruchten aller seiner Eier enthält.[39] Da das für Raubspinnen (Pisauridae) typische Brutgespinst, das ein begattetes Weibchen einige Zeit nach der Paarung anlegt, diesem viel Zeit, Kraft und Aufwand abverlangt, sollte sich das Weibchen theoretisch möglichst bald nach der Häutung in das Adultstadium verpaaren, um somit die dafür verfügbare Zeit zu maximieren.[40]
Kokonbau und Eiablage

Nach der Paarung beginnt das begattete Weibchen einen für Spinnen typischen Eikokon anzufertigen, in den auch die Eier abgelegt werden. Das zeitliche Auftreten der Kokons variiert je nach geographischer Lage. Während in Florida bereits Eikokons der Sechspunkt-Jagdspinne im April vorfindbar sind, ist dies in den nördlicheren Breiten erst im Zeitraum zwischen Juni und September der Fall.[9]
Die Dotterproduktion erfolgt wie bei den meisten anderen Spinnen über zwei Phasen. In der ersten noch vor der Paarung stattfindenden Phase aggregieren beziehungsweise vereinigen sich Dotterpartikel in der sich bildenden Eizelle zu feinkörnigen Elementen. In der zweiten nach der Paarung folgenden Phase bildet sich eine weitere Ansammlung aus Dotterpartikeln, die hier die Gestalt deutlich größerer Körner annehmen. In dieser Zeit und somit nach der Paarung nimmt das Weibchen deutlich mehr Nahrung zu sich als gewöhnlich und zeigt eine höhere Bereitschaft zu Kannibalismus, da die zweite Phase der Dotterproduktion nur gewährleistet werden kann, wenn dem Weibchen ausreichend Energie zur Verfügung steht. Außerdem schwillt das Opisthosoma des Weibchens durch die darin heranreifenden Eier sichtbar an. Aufgrund dessen ist Periode der Dotterbildung von sich wiederholenden Futtersuchen des Weibchens geprägt.[41]
Brutpflege

Die Sechspunkt-Jagdspinne betreibt wie alle Raubspinnen (Pisauridae) Brutpflege. Der Eikokon wird vom Weibchen anfangs für drei bis vier Wochen mithilfe seiner Cheliceren an sich geheftet permanent getragen. In dieser Zeit nimmt es keine Nahrung zu sich und bewegt sich kaum. Sobald der Schlupf der Jungtiere bemerkbar wird, fertigt das Weibchen ein für Vertreter dieser Familie übliches Brutgespinst an, das in unmittelbarer Nähe gefertigt wird. Der Standort dafür wird allem Anschein nach noch vor der Produktion des Eikokons ausgewählt. Faktoren, die die Wahl des Standorts beeinflussen, können neben der notwendigen Stabilität des Standortes das Auftreten von Prädatoren einschließlich Artgenossen und die am Standort herrschenden Witterungen sein.[42]
Unbekannt ist der Grund für die währenddessen verstärkte Aktivität des Weibchens, das nicht durch die zuvor erläuterten Faktoren beeinflusst werden kann. Sollte das Weibchen instinktiv durch die Suche nach geeignete Aufenthaltsorten angetrieben werden, so müsste dies auch in beliebigen Zeiträumen nach der Paarung der Fall sein. Demnach müssten einige Weibchen einen Tag nach seiner letzten Häutung für die Adulthäutung geeignete Aufenthaltsorte finden und an diesen verbleiben, um Ressourcen zu sparen, und dies insbesondere dann, wenn nur wenige für den Aufenthalt geeignete Positionen verfügbar wären. Stattdessen bewegt sich das Weibchen der Sechspunkt-Jagdspinne bis zur Herstellung eines Eikokons frei umher.[42] Das Weibchen bewacht den Eikokon auch weiterhin, sobald er sich im Brutgespinst befindet.[1]
Faktoren des Reproduktionserfolgs
_(14591910267).jpg.webp)
Der Erfolg der Reproduktion der Sechspunkt-Jagdspinne wird wie bei Spinnen üblich maßgeblich von der Verfügbarkeit an Beutetieren für das Weibchen mitbestimmt. Davon auch betroffen sind das Wachstum der Individuen und die Populationsdichte der Art. Sollte das Angebot an Nahrung für ein begattetes Weibchen nicht knapp sein, so fällt die Reproduktionsleistung bei größeren Weibchen in dem Fall höher aus als bei kleineren. Bei geringer Nahrungsverfügbarkeit zeigen große Weibchen oft eine geringe Reproduktionsleistung oder ein Reproduktionsversagen. Kleinere Weibchen hingegen benötigen weniger Energie für eine erfolgreiche Reproduktion. Daraus erschließt sich, dass kleinere Weibchen einen gleichwertigen oder größeren Fortpflanzungserfolg erzielen als große Weibchen, sollte ein geringes Nahrungsangebot im jeweiligen Habitat der Fall sein. Ein sehr starker Mangel an Beutetieren führt jedoch fast immer zu einer nicht abschließbaren Reproduktion. Ein ausreichendes Nahrungsangebot muss also für die Bildung von zumindest einem Eikokon gegeben sein. Die Körpergröße der Individuen der Sechspunkt-Jagdspinne hängt vom Nahrungsangebot der Jungtiere ab, womit dies auch später den Reproduktionserfolg beeinflusst.[16] Die Verfügbarkeit der Beutetiere ist somit insbesondere für größere Weibchen der Sechspunkt-Jagdspinne ein beeinflussender Faktor für deren Häufigkeit. Durch die Auswirkung des Nahrungsaufkommens auf größere und kleinere Weibchen wird der Selektion zur Erhöhung der Größe weiblicher Tiere eine weitere Selektion nach der Fähigkeit entgegengewirkt, das begrenzte Nahrungsangebot in einem beliebigen Habitat zu nutzen.[43]
Bei einem ausreichend großen Nahrungsangebot legt ein verpaartes Weibchen ein größeres Brutgespinst sowie einen schwereren Eikokon an und hat eine stärker ausfallende Gewichtszunahme infolge der heranreifenden Eier, als es bei einem kleineren Weibchen der Fall ist. Allerdings übt die Größe des Weibchens keinen Einfluss auf die Anzahl der Tage, die für die Herstellung eines Eikokons aufgebracht werden müssen, und die Masse eines einzelnen Eis aus. Dies trifft in dem Fall auch auf das Nahrungsangebot zu. Insbesondere erstgenannter Aspekt legt der Vermutung nahe, dass die Größe der Eier der Sechspunkt-Jagdspinne immer konstant bleibt und ein Weibchen im Falle eines geringeren Nahrungsangebots weniger Eier legt, bei denen jedoch keine Größenunterschiede vorliegen. Auch bei anderen Spinnenarten ließ sich eine Relation zwischen dem Nahrungsangebot und der Menge an abgelegten Eiern belegen. Zwar ist neben der Größe auch das Gewicht der Eier bei der Sechspunkt-Jagdspinne relativ einheitlich, allerdings kann dafür deren Qualität abhängig vom Nahrungsangebot stark variieren.[43]
Schlupf und Heranwachsen der Jungtiere
Die nach einiger Zeit schlüpfenden Jungtiere verweilen anfangs noch in dem Brutgespinst und werden von ihrer Mutter überwacht. Dies endet mit dem Beginn derer Zerstreuung, die im Regelfall eine Woche nach dem Schlupf eintritt.[1] Die Zerstreuung der Jungtiere findet bei der Sechspunkt-Jagdspinne vermutlich mithilfe des sogenannten Spinnenflugs im Herbst statt.[44]
Die Jungtiere der Sechspunkt-Jagdspinne wachsen wie bei Gliederfüßern üblich über mehrere Häutungsstadien heran. Dabei kann die Dauer des Heranwachsens variieren und wird von verschiedenen Faktoren beeinflusst. Besonders gut ist dies bei Populationen der Art in Alberta erforscht worden. Dort benötigen die Jungtiere der Sechspunkt-Jagdspinne für die Entwicklung allgemein zwei Jahre und durchlaufen dabei 12 Stadien. Innerhalb dieser Provinz überwintern die Jungtiere dabei zweimalig. Die erste Überwinterung findet in den Stadien drei bis fünf und die zweite in den Stadien acht bis zehn statt. Die meisten kleineren Jungtiere, die nach der ersten Überwinterung in Alberta anzutreffen sind, erreichen im gleichen Jahr noch nicht die Geschlechtsreife, obwohl innerhalb dieser Zeit auch das Auftreten mittelgroßer Jungtiere der Art belegt ist.[23]
Individuen der Sechspunkt-Jagdspinne, die in Alberta Mitte Juni ihre Geschlechtsreife erlangen würden, wären jedoch der nachteiligen Situation ausgesetzt, nur begrenzte Möglichkeiten für eine Paarung zu erhalten. Außerdem könnten diese Tiere gesundheitlichen Einbußen ausgesetzt sein, wobei die Wahrscheinlichkeit davon bei begatteten Weibchen, noch einen zweiten Eikokon zu produzieren zu können, erhöht wird. Daher wäre die zweite Überwinterung für die Populationen der Sechspunkt-Jagdspinne in Alberta enorm von Vorteil. Da Spinnen in den arktischen Regionen im Regelfall mehr als zwei Jahre für das Erlangen des ausgewachsenen Stadiums benötigen, wird dies auch bei der Sechspunkt-Jagdspinne vermutet.[23]
Wachsen die Jungtiere an einem Gewässer mit einem reichlichen Nahrungsangebot auf, erreichen diese im ausgewachsenen Stadium eine höher ausfallende Endgröße. Jungtiere, die sich an Gewässern mit geringer Dichte an Beutetieren entwickeln, haben zwei Möglichkeiten, um dann im ausgewachsenen Stadium eine erfolgreiche Reproduktion zu gewährleisten. Sie nehmen in diesem Stadium entweder eine kleinere Endgröße an oder wechseln zu einem anderen Lebensraum mit einer größeren Menge an Beutetieren. Die ausgewachsenen Spinnen wechseln kaum noch ihr Habitat. Daher wird die Fähigkeit der Sechspunkt-Jagdspinne für Anpassungen an einen jeweiligen Lebensraum mitsamt der anzunehmenden Körpergröße und eventuell die Annahme eines potentiell geeigneten Habitats vor allem in der Ausbreitungstätigkeit der Jungtiere gesehen.[45]
Habitatwechsel
Die Jungtiere der Sechspunkt-Jagdspinne nehmen eine standorttreue Lebensweise fast das ganze Jahr über an.[46] Dennoch nutzen sie innerhalb ihrer Saison höchstwahrscheinlich die Wasseroberfläche und die unmittelbare Uferzone. Diese Theorie wird durch die Tatsache gestützt, dass ein stetiges Wachstum von Jungtieren im Frühjahr und Sommer beobachtet werden kann, auch wenn bei Jungtiere zwischen den Stadien fünf bis sieben eine Absenz zu vermerken ist. In der ersten Überwinterung der Jungtiere innerhalb des dritten und fünften Stadiums können diese auch in Arealen fernab von Gewässern anzutreffen sein. Es ist möglich, dass die Jungtiere in diesem Fall innerhalb dieser Zeit zu einer Lebensweise an Land wechseln und unmittelbar nach der Überwinterung eine oder zwei weitere Häutungen durchführen. Außerdem besteht die Theorie, dass die Jungtiere entweder den direkten Bereich nahe dem Gewässerufer verlassen oder geschützte Überwinterungsplätze in der Einstreu oder in Klumpen von Schlitten und einzelnen Pflanzen suchen. Ähnliche Verhaltensweisen sind bei den Jungtieren der ebenfalls zu den Uferjägern (Dolomedes) zählenden Gerandeten Jagdspinne (D. fimbriatus) ausgeprägt. Im Gegensatz zu denen der anderen Art legen mittelgroße Jungtiere der Sechspunkt-Jagdspinne keinerlei Gespinste an.[47]
Allgemein werden für räuberische Tiere und darunter gerade für Spinnen Faktoren, wie die Struktur des jeweiligen Habitats und dessen Mikroklima, die Menge an Beutetieren und die Vermeidung von Prädatoren sowie Kannibalismus, als ausschlaggebend für Habitatwechsel gesehen. Die ontogenetisch (ein einzelnes Lebewesen betreffende) Entwicklung der Sechspunkt-Jagdspinne dient dementsprechend der Vermeidung eines Zusammenkommens mit ebenfalls auf dem Wasser jagenden Prädatoren und Nahrungskonkurrenz genauso wie Kannibalismus mit Artgenossen. Jungtiere der Art sind gerade vor und nach ihrer ersten Überwinterung verschiedenen Risiken und Gefahren ausgesetzt. Dazu zählen unter anderem neben größeren Artgenossen auch semi- und aquatische Prädatoren, wie Wasserläufer und Ruderwanzen, die jedoch wiederum auch größeren Individuen der Sechspunkt-Jagdspinne zum Opfer fallen können. Vertreter beider Wanzenfamilien sind insbesondere im Juli und im August häufig in den gleichen Lebensräumen wie die Sechspunkt-Jagdspinne vorfindbar und erbeuten kleine und mittelgroße Jungtiere der Art. Im Frühjahr und im Juni wären kleinere Jungtiere häufig von größeren und in der prä-subadulten Phase befindlichen Jungtieren bedroht, zumal letztere sich schneller entwickeln.[47]
Regulationen des Lebenszyklus
Wie bei Spinnen üblich, so üben auch bei der Sechspunkt-Jagdspinne Faktoren wie Temperatur, Tageslänge, endogene (auf innere Ursachen zurückzuführende) Mechanismen und die Anzahl an verzehrten Beutetieren einen starken Einfluss auf den Lebenszyklus und das Heranwachsen der Jungtiere der Art aus. So kann etwa die Fähigkeit zur Anpassung der Entwicklungsdauer der Jungtiere als adaptive Fähigkeit gesehen werden. Im Herbst hindert die kürzere Tageslänge kleinere Jungtiere, die noch aktiv auf der Suche nach Beutetieren befindlich sind, an Häutungen zum sechsten oder siebten Stadium. Diese Tiere können möglicherweise nicht überwintern. Wärmere Temperaturen im Spätherbst hingegen regen bereits fortgeschrittenere Jungtiere zu weiteren Häutungen an. Dadurch wird im Falle dieser für die Fortpflanzung nicht optimalen Umweltbedingungen ein Erlangen der Geschlechtsreife verhindert. In Gegenden mit langanhaltenden Eisperioden, etwa Alberta, kann dort nach dieser typischen Periode von Tagen mit kurzzeitigem Tageslicht die sich fortsetzende Entwicklung bereits größerer Jungtiere der Sechspunkt-Jagdspinne bei relativ niedrigen Temperaturen scheinbar sogar gefördert werden. Da für die Sechspunkt-Jagdspinne etwa in Alberta optimale Bedingungen für Paarungen zeitlich begrenzt sind, könnte das gleichzeitige Auftreten der Entwicklung auf das Erwachsenenstadium und anderen bereits etwas größeren Jungtieren unter den jeweiligen Witterungen im Frühling die bedeutendste Folge der Adaptionsfähigkeit der Entwicklungsrate sein.[23]
Die Überwinterung erfolgt bei bereits größeren Jungtieren in Form einer Ruhephase. Die Überwinterung wird in diesem Falle durch die Witterungen und weniger durch die bereits vorausgegangene Überwinterung beeinflusst. Die Anpassung auf derartige Überwinterungsstadien sollten ohne große Probleme für die Jungtiere umsetzbar sein, sollten sie zu den Populationen der Sechspunkt-Jagdspinne in den Gebieten zählen, in denen eine semivoltine (über ein Jahr dauernde) Entwicklung vorausgesetzt ist. Bei der ersten Überwinterungsperiode ist dies jedoch deutlich komplexer, da sich niedrige, zyklische oder sinkende Temperaturen genauso wie kurze Tageslängen allein für die Beeinflussung der Resistenz gegenüber kälteren Witterungen bei den Jungtieren in diesem Fall nicht als wirksam erweisen. Obwohl nach bisherigen Kenntnissen die Entwicklung der Jungtiere der Sechspunkt-Jagdspinne anscheinend sowohl von der Tageslänge als auch von der Temperatur beeinflussbar zu sein scheint, zeigen sich bei Beobachtungen auch Kurztagperioden gemeinsam mit zyklischen Temperaturen sowie einer Kurzlichtzeit mit Temperaturen von 4 °C einflusslos auf die Entwicklung der Jungtiere.[48]
Verhaltensänderungen
Bei der Sechspunkt-Jagdspinne sind je nach Stadium und Zustand der jeweiligen Individuen große Verhaltensunterschiede zu vermerken. Die Jungtiere der Sechspunkt-Jagdspinne zeigen eine verglichen mit den ausgewachsenen Spinnen deutlich geringere Bereitschaft für Nahrungssuchen und bevorzugen stattdessen Lauerjagden. Der Grund dafür ist nicht gänzlich geklärt, zumal sich eine verringerte Nahrungsaufnahme bei jüngeren Spinnen negativ auf deren spätere Fruchtbarkeit auswirken kann. Dies dürfte wiederum auf die nachlassende Fähigkeit zum erfolgreichen Tragen der Eier zurückzuführen sein. Eine Vermutung wäre, dass die Jungtiere durch die Lauerjagden ihren Energieverbrauch insbesondere dann, wenn es sich letztendlich um kleinere Beutetiere handelt, deutlich minimieren können. Junge Weibchen der Sechspunkt-Jagdspinne, die erst kürzlich das Adultstadium erreicht haben, gehen zu einer aktiveren Lebensweise über, in der häufige Fortbewegungen zu vermerken sind. In der Periode zwischen der Adulthäutung und dem Fertigen der Eikokons wächst der Nahrungsbedarf des Weibchens der Art generell aufgrund der für die Herstellung der Eidotterbestandteile notwendigen Ressourcen.[49]
Es wird vermutet, dass das unterschiedliche Verhalten der Jungtiere und der ausgewachsenen Exemplare der Sechspunkt-Jagdspinne mitunter durch jeweils verschiedene Prädatoren beeinflusst wird, obgleich die Sterberate einzelner Individuen der Art durch das Erbeuten von Prädatoren unbekannt ist. Das Erbeuten der Sechspunks-Jagdspinne durch größere Fische konnte belegt werden, als sich Exemplare der Art auf der offenen Wasseroberfläche fortbewegten. Ebenso kommen Wegwespen und insbesondere der Blaureiher (Egretta caerulea), die beide mithilfe ihres Sehsinns jagen, als bedeutende Feinde der Sechspunkt-Jagdspinne infrage. Sowohl Jungtiere als auch ausgewachsene Weibchen der Sechspunkt-Jagdspinne kommen möglicherweise als Beute verschiedener Wirbeltiere, etwa von Fischen, Fröschen oder verschiedenen Vögeln aus der Ordnung der Regenpfeiferartigen (Charadriiformes), in Frage. Die Endgröße adulter Weibchen dürfte diese allerdings zu einem deutlich ungeeigneteren Beuteobjekt für andere an Gewässern lebende Wirbellose werden lassen. Ferner ist die Gefahr durch Kannibalismus insbesondere für Jungtiere höher. Der Wechsel von einer stationären zu einer aktiveren Lebensweise erfolgt nicht graduell während des Heranwachsens, sondern findet nach der Adulthäutung statt. Außerdem wird durch das stationäre Verhalten der Jungtiere vom Zeitraum zwischen August und Oktober, wo nur noch wenige ausgewachsene Weibchen auf den Gewässern verbleiben, vermutet, dass diese sogar während der Absenz von Fressfeinden Bewegungen vermeiden.[50]
Adaptive Anpassungen bei Verhaltensänderungen
Das Weibchen der Sechspunkt-Jagdspinne zeigt eine geringere Scheu, wenn Nahrung verfügbar ist, und verbringt dann mehr Zeit auf der Wasseroberfläche als darunter. Gleiches trifft auch zu, sollte sich in der Nähe eines Weibchens ein balzendes Männchen befinden, das für das Weibchen dann entweder einen Geschlechtspartner oder eine Nahrungsquelle darstellen kann. Die Scheu seitens des Weibchens ist noch geringer, sollte es zu einem beliebigen Zeitpunkt Nachwuchs bewachen. Letzteres kann jedoch auch aus dem Energieverbrauch des Weibchens für die Produktion der Eier oder einem gegebenenfalls vorkommenden und längeren unter Wasser stattfindenden Aufenthalt mitsamt dem Kokon resultieren.[51]
Entgegen der Hypothese, dass die ausgewachsenen Spinnen durch ihre höher ausfallende Endgröße einem geringeren Risiko des Erbeutens von Prädatoren ausgesetzt sind und deshalb weniger scheu als ihre jüngeren Verwandten sein sollten, zeigen alle Individuen der Sechspunkt-Jagdspinne unabhängig vom Alter keine signifikanten Unterschiede bezüglich ihres wenig scheuen Verhaltens. Dies ist in Anbetracht dessen bemerkenswert, dass gerade die ausgewachsenen Individuen einer größeren Nahrungsknappheit ausgesetzt und demzufolge aktiver sowie weniger scheu sein können. Eine mögliche Erklärung für die nahezu gleichermaßen geringe Scheu, die bei der Sechspunkt-Jagdspinne stadienübergreifend vorhanden ist, könnte der für Jungtiere aufwendigere Aufenthalt unter Wasser sein, die diese somit zu einer geringeren Aktivität veranlasst.[51]
Variationen und Entwicklung der Scheuheit
Die geringe Scheuheit der Sechspunkt-Jagdspinne kann selber variabel und abhängig vom Stadium und der aktuellen Situation ausgeprägt sein. Beim Weibchen der Art ist dieses Verhalten abgesehen vom Falle der Brutpflege, wo es erhöht wird, nahezu konstant ausgeprägt. Während der Nahrungssuche ist die Scheu bei der Spinne allgemein gering. Umgekehrt, aber ähnlich konstant, verhielt es sich bei 2007 stattgefundenen Experimenten in Gefangenschaft, bei denen Exemplare der Sechspunkt-Jagdspinne auf ihre Reaktionen untersucht wurden, wobei die Resultate hier weniger einheitlich waren und die Versuchstiere teilweise unterschiedlich reagierten.[51] Auf ausgelöste Störungen reagierten die allesamt weiblichen Exemplare größtenteils mit einer Flucht unter Wasser.[52] Die geringe Scheuheit ist bei ausgewachsenen Individuen der Sechspunkt-Jagdspinne vor allem bei der Nahrungssuche und der Balz sowie bei allgemeiner Untätigkeit bemerkbar, jedoch nicht bei Weibchen, die Brutpflege betreiben.[51]
Bei Jungtieren der Sechspunkt-Jagdspinne ist im Gegensatz zu den ausgewachsenen Tieren generell eine höhere Scheuheit vorhanden. Trotzdem scheint es nicht selten vorzukommen, dass juvenile Individuen der Art ihren Ausdruck an Scheuheit auf das Adultstadium übertragen. Sollte die Scheuheit bei einem einzelnen Individuum in einem Ausmaß vorhanden sein, das über die Entwicklungsphasen des Individuums hinweg konstant bleibt, dann ist dieses Individuum in diesem Falle durch die gegebenen Umweltbedingungen in einer Anpassung dieses Verhaltens eingeschränkt. Mangels Ergebnissen für vollständige Verhaltensentwicklung einzelner Exemplare der Sechspunkt-Jagdspinne ist diese Entwicklung jedoch auch hinsichtlich möglicher Hindernisse sowie Korrelationen nicht genau beurteilbar.[51]
Die verringerte Scheuheit bei Weibchen mit Eikokons
Die bereits erläuterte und noch weiter verringerte Scheuheit beim Weibchen der Sechspunkt-Jagdspinne, sofern es einen Eikokon trägt, lässt sich damit begründen, dass die Herstellung des Kokons dem Weibchen ein hohes Maß an Ressourcen abverlangt. Außerdem kann das Weibchen in dem Falle, sofern es sich unter Wasser befindet, auch schneller als gewöhnlich wieder auftauchen und sich weiterhin über Wasser aufhalten. Dabei handelt es sich wahrscheinlich um eine Strategie des Weibchens gegen Prädatoren im Falle des Tragen eines Eikokons, da das Weibchen durch dessen Tragen hinsichtlich seiner Verfügbarkeit an Energie und/oder seiner Bewegungsfähigkeit eingeschränkt ist. Diese Theorie wird dadurch gestützt, dass dieses Verhalten auch in Wechselbeziehung mit dem körperlichen Zustand des Weibchens steht.[51]
Demnach wäre nur Weibchen mit ausreichender Kondition ein längerer Aufenthalt unter Wasser möglich und physische Einbußen würden einem Weibchen in dem Fall die oben erläuterte Strategie gegen Prädatoren erschweren, was wiederum das Tier in dem Fall in Anbetracht des bereits erbrachten Aufwands für die Brutpflege weniger scheu erscheinen ließe. Dies verdeutlicht, dass diese Verhaltensmerkmale der Sechspunkt-Jagdspinne nicht für sich einzeln stehen, sondern als Neigung zu einem seitens der Spinne gleichmäßigen Gesamtverhalten betrachtet werden können.[53]
Theorien zur Entwicklung der Scheuheit
Eine direkte Ausprägung einer geringen Scheuheit fehlt bei den Jungtieren der Sechspunkt-Jagdspinne im Gegensatz zu den ausgewachsenen Individuen, was darauf schließen lässt, das dessen Ausprägung über die Entwicklung der Jungtiere hinweg nicht stabil ist. Das für Spinnen übliche und unstetige Wachstum wird dafür als möglicher Grund gesehen, da durch dieses deutliche Umstellungen zwischen den verschiedenen Häutungsstadien mitsamt dem subadulten und dem adulten Stadium erkennbar werden. Dies könnte auch eine maßgebliche Umstrukturierung des Verhaltens einschließlich dessen Mechanismen ermöglichen, womit das Vorhandensein der verringerten Scheuheit bei ausgewachsenen Individuen der Sechspunkt-Jagdspinne erklärt werden könnte.[54]
Systematik
Die klassische Systematik befasst sich im Bereich der Biologie sowohl mit der taxonomischen (systematischen) Einteilung als auch mit der Bestimmung und mit der Nomenklatur (Disziplin der wissenschaftlichen Benennung) von Lebewesen einschließlich der Sechspunkt-Jagdspinne.
Der Artname triton bezieht sich auf den Meeresgott Triton der griechischen Mythologie und somit auf die amphibische Lebensweise der Art.[55]
Beschreibungsgeschichte und Synonymie
Charles Athanase Walckenaer gab der Sechspunkt-Jagdspinne bei der Erstbeschreibung 1837 die Bezeichnung Lycosa triton und ordnete sie somit als eine zur Familie der Wolfsspinnen (Lycosidae) zählende Art innerhalb der Gattung Lycosa ein.[56] Der Artname triton geriet jedoch in Vergessenheit, sodass die 1845 von Nicholas Marcellus Hentz angewandte Bezeichnung Dolomedes sexpunctatus für lange Zeit verwendet und die Art somit damals bereits in die Gattung der Uferjäger (Dolomedes) transferiert wurde.[56][57] Dies hielt bis zu einer 1924 von Sherman Chauncey Bishop durchgeführten Revision (Überarbeitung der Systematik) an. Derselbe Autor beschrieb dabei neben der Nominatform D. t. triton die Unterart D. t. sexpunctatus und nutzte für die Differenzierung zwischen der Nominatform und der Unterart die Verbreitungsgebiete dieser. Während letztere sowohl im Norden des Verbreitungsgebiets der Sechspunkt-Jagdspinne und in den Südatlantikstaaten beispielsweise im Okefenokee Swamp verbreitet sein soll, so soll die Nominatform lediglich im Süden vorkommen. Der Status von D. t. sexpunctatus wurde 1973 unter James E. Carico aufgehoben. Ein Grund dafür ist die Sympatrie (Überlappung der Verbreitungsgebiete) der Unterart und der Nominatform, die der Voraussetzung für eine Unterart in Form der Allopatrie (nicht stattfinde Überlappung der Verbreitungsgebiete) mehrerer Unterarten einer Art unter Einbeziehung der Möglichkeit zur Kreuzung dieser widerspricht. Außerdem konnte im Nachhinein bestätigt werden, dass Bishop für die Individuen der Sechspunkt-Jagdspinne, die innerhalb der für die Unterart und die Nominatform genannten Gebieten gefunden wurden, variable Merkmale angab, zumal intrapopulationale Überschneidungen und die von Bishop angegebenen Merkmale bei Exemplaren beider angeblicher Formen im Norden des Verbreitungsgebiets der Sechspunkt-Jagdspinne nachgewiesen wurden. Von Bishop besonders hervorgehoben wurden die Breite des Carapax bei den jeweiligen Individuen und die variierende Größe der Männchen.[57]
Bei der Revision beschrieb Bishop auch die heute mit der Sechspunkt-Jagdspinne synonymisierte Art Dolomedes albiclavius und gab für diese unter anderem drei sehr voneinander entfernte Verbreitungsgebiete an. Davon ist eines die im US-Bundesstaat Missouri gelegene Stadt Springfield und die anderen beiden der Große Salzsee im Norden des US-Bundesstaats Utah sowie Billy’s Island inmitten des Okefinokee Swamps. Außerdem gab Bishop an, dass es sich bei D. albiclavius um eine große und ausgeprägt gezeichnete Art handelt, die mit der Sechspunkt-Jagdspinne nah verwandt sei. Von letzterer Art unterscheidet sich D. albiclavius laut Bishop durch den Aufbau der Epigyne und durch die insgesamt dunkler ausfallende Farbgebung sowie durch das Fehlen der für die Sechspunkt-Jagdspinne typischen und weißen Fleckenpaare auf dem Opisthosoma. Ferner soll D. albiclavius sowohl proportional als auch tatsächlich längere Beine als die Sechspunkt-Jagdspinne aufweisen, während der Körper überdies robuster als bei der anderen Art gebaut sein sollte. Der Von Bishop verwendete weibliche Holotypus (für die Erstbeschreibung angewandtes Exemplar) und überdies das einzige überlieferte Exemplar, das der Art D. albiclavius angehörig sein soll, besitzt zwar tatsächlich eine dunkle Färbung und überdies fehlende Punktpaare, doch ist letzterer Aspekt das Ergebnis starker und schon vor längerer Zeit geschehener Abreibung. Zuletzt entsprechen der Aufbau der Epigyne sowie die Form der Beine vom untersuchten Holotypus denen der Sechspunkt-Jagdspinne. Unter diesen Anhaltspunkten hat Carico 1973 den Artstatus von D. albiclavius aufgehoben.[57]
Allgemein waren die Konzepte dieser Taxa (biologische Einheiten) seitens Bishop für lange Zeit gültig und lediglich eine Abwandlung von einem seiner Werke wurde bis zu den grundlegenden Überarbeitungen der Taxa unter Carico im Jahr 1973 veröffentlicht. Ralph Vary Chamberlin und Wilton Ivie entwarfen 1946 das Konzept einer nach der Sechspunkt-Jagdspinne benannten Artengruppe, die neben dieser noch drei weitere Arten aus der Gattung der Uferjäger umfasst, wobei Merkmale der Tibiaapophysen bei Männchen als Differenzierungsmerkmale herangezogen wurden.[57] Eine dieser vorgeschlagenen Arten wäre die von den gleichen Autoren im selbigen Jahr erstbeschriebene Art D. scopularis, deren Bezeichnung auf einer 1847 von Carl Ludwig Koch für die Sechspunkt-Jagdspinne verwendete Bezeichnung für die Sechspunkt-Jagdspinne basiert.[57][56] Von D. scopularis wurden lediglich Männchen beschrieben, bei denen laut Autoren die Tibiaapophyse nach oben um 40° gedreht sei. Die zweite Art war D. sexpunctatus auf Basis der Beschreibungen von Hentz und außerdem die einzige, deren Beschreibung die Kriterien für einen vollständigen Artstatus erfüllt. Als Eigenmerkmal der angeblichen Art nutzte man die schmale Spitze der Tibiaapophyse, während die Sechspunkt-Jagdspinne selber anhand der intermediären (mittig liegenden) Erweiterung der Tibiaapophyse von den anderen vermeintlichen Arten abgegrenzt wurde. Für die Sechspunkt-Jagdspinne gaben Chamberlin und Ivie lediglich Vorkommen im US-Bundesstaat Georgia an.[58]
Die vierte Art dieser Gruppe wäre D. spatulatus, die von Chamberlin und Ivie anhand eines in der im US-Bundesstaat Tennessee gelegenen Stadt Kingston gefundenen Holotypus erstbeschrieben wurde. In der sehr kurzen Beschreibung dieser Art ist besonders die breite Erweiterung der Tibiaapophyse hervorgehoben. Bei vierzehn männlichen und in zwei Tagen an derselben Stelle gefundenen Exemplaren, bei denen es sich vermutlich um die von Chamberlin und Ivie verwendeten Paratypen (bei der Erstbeschreibung mitverwendeten Exemplare) von D. spatulatus handelt, wies deren apikale Erweiterung je nach Individuum eine sehr unterschiedliche Breite auf. Die für D. sexpunctatus schmaleren und die für D. spatulatus breiteren Apophyse sowie die der Sechspunkt-Jagdspinne, deren Maße zwischen denen der beiden anderen liegt und von Chamberlin und Ivie als Eigenmerkmale dieser drei Arten dienen, sind überlappend in den Verbreitungsgebieten aller drei Arten überlappend vorhanden, sodass D. sexpunctatus und D. spatulatus kein Artstatus zugesprochen werden kann. Ferner ist die Beschreibung von D. scopularis durch die dafür angewandten und sehr variablen Charaktermerkmale für eine Anerkennung als Art unzureichend, sodass diese Art zusammen mit D. sexpunctatus und D. spatulatus aufgrund der bereits erläuterten Gründe mit der Sechspunkt-Jagdspinne ebenfalls unter Carico 1973 synonymisiert wurden.[59]
Evolution und äußere Systematik

Die evolutionäre Entwicklung der Sechspunkt-Jagdspinne ist ebenso wie die der ebenfalls zu den Uferjägern (Dolomedes) zählenden und genauso in der Nearktis vorkommenden Arten D. albineus und D. striatus unzureichend erforscht, doch es ist anzunehmen, dass die großen Verbreitungsgebiete dieser Arten eine signifikante Rolle bezüglich deren Evolution einnehmen. Da das der Sechspunkt-Jagdspinne jedoch das umfangreichste aller Uferjäger innerhalb dieses Gebiets ist, wären gewisse Differenzen bei den verschiedenen Populationen zu erwarten. Allerdings wurden bisher außerhalb des Ostens der Vereinigten Staaten nur Jungtiere und seltener ausgewachsene Individuen der Art analysiert, die allesamt keine deutlichen Unterschiede zueinander aufwiesen. Ein Untersuchen weiterer Exemplare der Sechspunkt-Jagdspinne innerhalb dieser Areale könnte Aufschluss über mögliche, bisher unbekannte Variationen der Art geben.[60]
Ein Versuch, das verwandtschaftliche Verhältnis der nearktischen Vertreter der Uferjäger zu erforschen, ist 1973 unter Carico geschehen. Dazu wurden die Bulbi der Männchen der jeweiligen Arten als Indikator für die außensystematische Stellung zueinander genutzt, wobei darunter besonders der Grundaufbau der Median- und der Tibialapophyse als hilfreich für diese Analyse betrachtet werden können. Auch relevant dürften nach Carico der Aufbau des Tegulums und die Gesamtlänge des Cymbiums und der Tibia bei den männlichen Tieren der nearktischen Uferjägern mitsamt der Sechspunkt-Jagdspinne sein.[61]
Die neun nearktischen Arten der Uferjäger lassen sich nach Carico in zwei Artengruppen zusammenfassen. Davon ist eine die der Art D. tenebrosus und umfasst neben dieser die Art D okefinokensis.[62] Die übrigen Arten und darunter auch die Sechspunkt-Jagdspinne gehören zu der Artengruppe der Gerandeten Jagdspinne (D. fimbriatus), die jedoch selber nicht in Amerika vorkommt und somit auch nicht selber zu der Artengruppe zählt. Stattdessen wurden die genitalmorphologischen Merkmale dieser Art, die sehr denen der gleichnamigen Artengruppe der nearktischen, gattungsverwandten Arten ähnelt, als Basis für die Bildung dieser Artengruppe genutzt.[63]
Bei den Männchen der Artengruppe der Gerandeten Jagdspinne mitsamt der Sechspunkt-Jagdspinne ist bei einem einzelnen Bulbus jeweils die mediane Apophyse in der basalen Hälfte anders als im Falle der Artengruppe von D. tenebrosus nicht eindeutig am breitesten. Die Länge vom Cymbium und der Tibia beläuft sich bei beiden Artengruppen auf 2,7 Millimeter. Bei den Weibchen beider Artengruppen ist am posterioren Rand der Epigyne je eine mittig platzierte Lappung vorhanden. Bei der Artengruppe der Sechspunkt-Jagdspinne ist allerdings noch zusätzlich ein konkaver Verlauf im Medianbereich vorhanden, der bei der anderen Artengruppe fehlt.[64] Folgendes Kladogramm verdeutlicht die bisher ermittelten Stellungen der einzelnen Arten innerhalb der Gattung der Uferjäger:[62]
| Uferjäger (Dolomedes) |
| |||||||||||||||||||||||||||||||||
Sechspunkt-Jagdspinne und Mensch

Die Art gilt beispielsweise aufgrund ihres Verbreitungsgebiets als geeignetes Forschungsobjekt für den Lebenszyklus von Spinnen, die in arktischen Regionen präsent sind.[23] Außerdem wird der Sechspunkt-Jagdspinne auch wie allen Spinnen die wichtige Funktion in Ökosystemen zugesprochen, ein dortiges Massenauftreten von Insekten zu verhindern.[1] Gleichzeitig ist bekannt, dass die Art ebenso große Mengen an in Fischbrutbetrieben gehaltenen Fischen erbeuten und somit auch als Schädling in Erscheinung treten kann.[18]
Einzelnachweise
- Lindsay Lane: Dolomedes triton. In: Animal Diversity Web. University of Michigan, abgerufen am 13. Juni 2021.
- Charles D. Dondale, James H. Redner: The insects and arachnids of Canada, Part 17. The wolf spiders, nurseryweb spiders, and lynx spiders of Canada and Alaska, Araneae: Lycosidae, Pisauridae, and Oxyopidae. In: Agriculture and Agri-Food Canada (Hrsg.): Research Branch Agriculture Canada Publication. Band 1856, Nr. 1, 1990, ISBN 0-660-13628-7, S. 337.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 482–483.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 483.
- Charles D. Dondale, James H. Redner: The insects and arachnids of Canada, Part 17. The wolf spiders, nurseryweb spiders, and lynx spiders of Canada and Alaska, Araneae: Lycosidae, Pisauridae, and Oxyopidae. In: Agriculture and Agri-Food Canada (Hrsg.): Research Branch Agriculture Canada Publication. Band 1856, 1990, ISBN 0-660-13628-7, S. 338.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 482.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 451.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 483–484.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 484.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 479.
- Richard A. Bradley: Common Spiders of North America. University of California Press, 2013, ISBN 978-0-520-27488-4, S. 181.
- Troy Bartlett: Species Dolomedes triton - Six-spotted Fishing Spider. In: BugGuide. Iowa State University, 16. Februar 2004, abgerufen am 13. Juni 2021.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 485.
- Charles D. Dondale, James H. Redner: The insects and arachnids of Canada, Part 17. The wolf spiders, nurseryweb spiders, and lynx spiders of Canada and Alaska, Araneae: Lycosidae, Pisauridae, and Oxyopidae. In: Agriculture and Agri-Food Canada (Hrsg.): Research Branch Agriculture Canada Publication. Band 1856, Nr. 1, 1990, ISBN 0-660-13628-7, S. 339.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 438.
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 44–45, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 436.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 437.
- G. S. Deshefy: ‘Sailing’ behaviour in the fishing spider, Dolomedes triton (Walckenaer). In: Animal Behavior. Band 29, Nr. 3, 1981, S. 965, doi:10.1016/S0003-3472(81)80042-5 (fdocuments.in [abgerufen am 22. Juni 2021]).
- Manfred Zimmermann, John R. Spence: Prey Use of the Fishing Spider Dolomedes triton (Pisauridae, Araneae): An Important Predator of the Neuston Community. In: Oecologia. Band 80, Nr. 2. Springer Science+Business Media, August 1989, S. 191, doi:10.1007/BF00380149.
- Manfred Zimmermann, John R. Spence: Prey Use of the Fishing Spider Dolomedes triton (Pisauridae, Araneae): An Important Predator of the Neuston Community. In: Oecologia. Band 80, Nr. 2. Springer Science+Business Media, August 1989, S. 192, doi:10.1007/BF00380149.
- Manfred Zimmermann, John R. Spence: Prey Use of the Fishing Spider Dolomedes triton (Pisauridae, Araneae): An Important Predator of the Neuston Community. In: Oecologia. Band 80, Nr. 2. Springer Science+Business Media, August 1989, S. 193, doi:10.1007/BF00380149.
- Manfred Zimmermann, John R. Spence: Phenology and life-cycle regulation of the fishing spider Dolomedes triton Walckenaer (Araneae, Pisauridae) in central Alberta. In: University of Alberta (Hrsg.): Canadian Journal of Zoology. Band 76, Nr. 2, Februar 2011, S. 307, doi:10.1139/z97-200 (researchgate.net [PDF; abgerufen am 22. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 13, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Chantal Roland, Jerome S. Rovner: Chemical and Vibratory Communication in the Aquatic Pisaurid Spider Dolomedes Triton. In: Ohio University (Hrsg.): The Journal of Arachnology. Band 11, Nr. 1, 1983, S. 83.
- Chantal Roland, Jerome S. Rovner: Chemical and Vibratory Communication in the Aquatic Pisaurid Spider Dolomedes Triton. In: Ohio University (Hrsg.): The Journal of Arachnology. Band 11, Nr. 1, 1983, S. 84.
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 27, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 18, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 18–19, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Chantal Roland, Jerome S. Rovner: Chemical and Vibratory Communication in the Aquatic Pisaurid Spider Dolomedes Triton. In: Ohio University (Hrsg.): The Journal of Arachnology. Band 11, Nr. 1, 1983, S. 82.
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 14, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 19, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 14–15, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 26, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 27–28, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 47–48, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 21–22, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 22, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 22–23, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 23, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Nancy Kreiter, David H. Wise: Age-related changes in movement patterns in the fishing spider, Dolomedes triton (Araneae, Pisauridae). In: Journal of Arachnology. Band 24, Nr. 1, Januar 1996, S. 29.
- Nancy Kreiter, David H. Wise: Age-related changes in movement patterns in the fishing spider, Dolomedes triton (Araneae, Pisauridae). In: Journal of Arachnology. Band 24, Nr. 1, Januar 1996, S. 30.
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 46, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 440.
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. 47, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF; abgerufen am 13. Juni 2021]).
- Nancy Kreiter, David H. Wise: Age-related changes in movement patterns in the fishing spider, Dolomedes triton (Araneae, Pisauridae). In: Journal of Arachnology. Band 24, Nr. 1, Januar 1996, S. 28.
- Manfred Zimmermann, John R. Spence: Phenology and life-cycle regulation of the fishing spider Dolomedes triton Walckenaer (Araneae, Pisauridae) in central Alberta. In: University of Alberta (Hrsg.): Canadian Journal of Zoology. Band 76, Nr. 2, Februar 2011, S. 306, doi:10.1139/z97-200 (researchgate.net [PDF; abgerufen am 17. Juni 2021]).
- Manfred Zimmermann, John R. Spence: Phenology and life-cycle regulation of the fishing spider Dolomedes triton Walckenaer (Araneae, Pisauridae) in central Alberta. In: University of Alberta (Hrsg.): Canadian Journal of Zoology. Band 76, Nr. 2, Februar 2011, S. 307–308, doi:10.1139/z97-200 (researchgate.net [PDF; abgerufen am 17. Juni 2021]).
- Nancy Kreiter, David H. Wise: Age-related changes in movement patterns in the fishing spider, Dolomedes triton (Araneae, Pisauridae). In: Journal of Arachnology. Band 24, Nr. 1, Januar 1996, S. 28–30.
- Nancy Kreiter, David H. Wise: Age-related changes in movement patterns in the fishing spider, Dolomedes triton (Araneae, Pisauridae). In: Journal of Arachnology. Band 24, Nr. 1, Januar 1996, S. 30–31.
- James Chadwick Johnson, Andrew Sih: Fear, food, sex and parental care: A syndrome of boldness in the fishing spider, Dolomedes triton. In: Animal Behaviour. Band 74, Nr. 5, November 2007, S. 1136, doi:10.1016/j.anbehav.2007.02.006.
- James Chadwick Johnson, Andrew Sih: Fear, food, sex and parental care: A syndrome of boldness in the fishing spider, Dolomedes triton. In: Animal Behaviour. Band 74, Nr. 5, November 2007, S. 1133, doi:10.1016/j.anbehav.2007.02.006.
- James Chadwick Johnson, Andrew Sih: Fear, food, sex and parental care: A syndrome of boldness in the fishing spider, Dolomedes triton. In: Animal Behaviour. Band 74, Nr. 5, November 2007, S. 1136–1137, doi:10.1016/j.anbehav.2007.02.006.
- James Chadwick Johnson, Andrew Sih: Fear, food, sex and parental care: A syndrome of boldness in the fishing spider, Dolomedes triton. In: Animal Behaviour. Band 74, Nr. 5, November 2007, S. 1137, doi:10.1016/j.anbehav.2007.02.006.
- Charles Athanase Walckenaer: Histoire naturelle des insectes. Aptères. Tome premier. In: Librairie encyclopédique de Roret. Paris 1837, S. 340.
- Naturhistorisches Museum der Burgergemeinde Bern: World Spider Catalog – Dolomedes triton. Abgerufen am 22. Juli 2021.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 486.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 486–487.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 487.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 443.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 443–444.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 444.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 448.
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 451.
Literatur
- Richard A. Bradley: Common Spiders of North America. University of California Press, 2013, ISBN 978-0-520-27488-4 (288 S.).
- James Edwin Carico: The Nearctic species of the genus Dolomedes (Araneae: Pisauridae). In: Bulletin of the Museum of Comparative Zoology. Band 144, Nr. 7. Harvard University Press, 13. März 1973, S. 435–488.
- G. S. Deshefy: ‘Sailing’ behaviour in the fishing spider, Dolomedes triton (Walckenaer). In: Animal Behavior. Band 29, Nr. 3, 1981, S. 965, doi:10.1016/S0003-3472(81)80042-5 (fdocuments.in).
- Charles D. Dondale, James H. Redner: The insects and arachnids of Canada, Part 17. The wolf spiders, nurseryweb spiders, and lynx spiders of Canada and Alaska, Araneae: Lycosidae, Pisauridae, and Oxyopidae. In: Agriculture and Agri-Food Canada (Hrsg.): Research Branch Agriculture Canada Publication. Band 1856, Nr. 1, 1990, ISBN 0-660-13628-7, S. 1–383.
- James Chadwick Johnson, Andrew Sih: Fear, food, sex and parental care: A syndrome of boldness in the fishing spider, Dolomedes triton. In: Animal Behaviour. Band 74, Nr. 5, November 2007, S. 1131–1138, doi:10.1016/j.anbehav.2007.02.006.
- Nancy Kreiter, David H. Wise: Age-related changes in movement patterns in the fishing spider, Dolomedes triton (Araneae, Pisauridae). In: Journal of Arachnology. Band 24, Nr. 1, Januar 1996, S. 24–33.
- Chantal Roland, Jerome S. Rovner: Chemical and Vibratory Communication in the Aquatic Pisaurid Spider Dolomedes Triton. In: Ohio University (Hrsg.): The Journal of Arachnology. Band 11, Nr. 1, 1983, S. 77–85.
- Jack P. Wojcicki: A description of the reproductive biology of the fishing spider Dolomedes triton (Walck.) (Araneae:Pisuridae) in central Alberta. Hrsg.: Library and Archives Canada. Ottawa 1992, ISBN 0-315-77142-9, S. i-65, doi:10.7939/R3R49GD71 (era.library.ualberta.ca [PDF]).
- Manfred Zimmermann, John R. Spence: Prey Use of the Fishing Spider Dolomedes triton (Pisauridae, Araneae): An Important Predator of the Neuston Community. In: Oecologia. Band 80, Nr. 2. Springer Science+Business Media, August 1989, S. 187–194, doi:10.1007/BF00380149.
- Manfred Zimmermann, John R. Spence: Phenology and life-cycle regulation of the fishing spider Dolomedes triton Walckenaer (Araneae, Pisauridae) in central Alberta. In: University of Alberta (Hrsg.): Canadian Journal of Zoology. Band 76, Nr. 2, Februar 2011, S. 295–309, doi:10.1139/z97-200 (researchgate.net [PDF]).
Weblinks
- Dolomedes triton im World Spider Catalog
- Dolomedes triton bei Global Biodiversity Information Facility.
- Lindsay Lane: Dolomedes triton. In: Animal Diversity Web. University of Michigan, abgerufen am 23. Juni 2021.
- Troy Bartlett: Species Dolomedes triton - Six-spotted Fishing Spider. In: BugGuide. Iowa State University, 16. Februar 2004, abgerufen am 23. Juni 2021.