Trichobothrium
Trichobothria (Einzahl Trichobothrium; altgriechisch θρίξ thrix, Gen. τριχός trichós, deutsch ‚Haar‘ und altgriechisch βοθρίον bothríon ‚Grübchen‘) sind sehr lange Tasthaare (Setae) bei einigen Gliederfüßern.[1] Sie sind beweglich in Aussparungen der Cuticula eingelenkt und dienen als mechanosensitive Sinnesorgane, welche besonders auf Luftschwingungen ansprechen und ein Hören ermöglichen.[1] Vor allem treten sie an den Beinen und Pedipalpen von Spinnentieren auf.[1] Aber auch einige Insekten (wie Käfer[2][3] und Schnabelkerfe[4]) tragen Trichobothria. Im Laufe der Evolution sind Trichobothria wahrscheinlich dreimal parallel entstanden.[4] Die Entwicklung von Trichobothria kann bei Skorpionen bis in die Kreidezeit zurückverfolgt werden.[5]

Die Orientierung durch Trichobothria heißt Orthobothriotaxis.[5] Eine andere, viel kleinere Form von Oberflächensensoren der Cuticula stellen Sensilla dar.
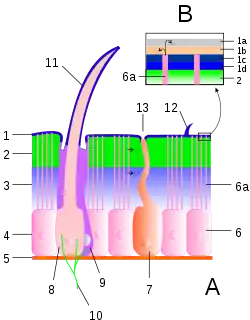
1 Epicuticula, B Details der Epicuticula:
1a Cementum, 1b Wachsschicht, 1c Epicuticula externa, 1d Epicuticula interna
2 Exocuticula, 3 Endocuticula
2 + 3 Procuticula
4 Epithelschicht, 5 Basalmembran
6 Epithelzelle, 6a Porenkanäle
7 Drüsenzelle
8 Cuticulagrube (Bothrion)
9 Wärmerezeptive Zellen
10 Nervenenden
11 Trichobothrium
12 Sensillum
13 Drüsenöffnungen
Struktur und Funktion
Da Gliederfüßer einen harten und starren Panzer aus Sklerotin und Chitin besitzen, müssen mechanosensitiven Ionenkanälen in Nervenmembranen mechanische Außenreize durch das Exoskelett zugeführt werden.
Auf Strömungen des Mediums (meistens Luftbewegungen) reagieren Trichobothria hochsensibel, weil die Haargebilde aus Chitinfäden einer Cuticulagrube entspringen. Die größte bislang bei einer Spinne festgestellte Zahl an Trichobothria beträgt bei der mittelamerikanischen nachtaktiven, stillsitzend auf Insekten lauernden Jagdspinne Cupiennius salei etwa 100 Trichobothria je Bein. Die feinen, im Durchmesser 5 bis 15 µm dünnen Fäden ragen nur um 100 bis 1500 µm aus der Cuticulaoberfläche heraus. Damit sie sich überhaupt durch geringste Luftbewegungen auslenken lassen, sind sie durch eine dünne, gespannte Membran in die Cuticula eingebracht. Mit wachsender Fadenlänge nimmt die Erregungsschwelle ab, sie liegt bei Luftgeschwindigkeiten von weniger als 1 mm/s. Der Auslenkungswinkel wird durch den Rand der Cuticulagrube auf maximal 25 bis 35° begrenzt.
Käfer können verschieden lange Trichobothria unterschiedlicher Typen am Hinterleib tragen und damit hören, dies kann der Kommunikation untereinander dienen.[2] Schnabelkerfe wie Wanzen tragen Trichobothria an ihren Antennen.[4]
Wahrnehmung
Fliegende Insekten erzeugen Luftvibrationen von Frequenzen bis etwa 600 Hz, und da sich Trichobothria wie Tiefpassfilter verhalten, reagieren sie auf solch tiefe Frequenzen. Je länger die Fadenhaare sind, desto besser werden niedere Frequenzen übertragen. So lässt sich die Schwingungsempfindlichkeit an den für ihre Lebensweise wichtigen Frequenzbereich anpassen. An jeder überströmten Fläche bildet sich ein Film unbewegter Luft. Aus dieser sogenannten Grenzschicht müssen die Haare herausragen, damit die Trichobothria richtig reagieren können. Mit abnehmender Strömungsfrequenz wächst die Dicke dieser Grenzschicht. Die Haarlänge ist optimal angepasst, wenn sie umgekehrt proportional zur Wurzel aus der Reizfrequenz ist. So wie lange Trichobothria sich fast gleich gut in alle Richtungen auslenken lassen, gibt es stattdessen bei den kurzen Richtungsselektivität, die nicht durch die asymmetrischen Ränder der Cuticulagruben hervorgerufen wird, sondern durch die Eigenschaften der Gelenkmembran. An der Insertion von drei oder vier mechanosensitiven Dendriten werden die Trichobothria innerviert, dabei antwortet jeder Dendrit auf einen bestimmten Auslenkungssektor. Also entsteht die Richtungsempfindlichkeit erst beim Transduktionsprozess durch die Lage der Dendriten im Gelenk. Es ist noch nicht genau erforscht, ob der Druck auf die Dendritenmembran der adäquate Reiz ist.
Die Kraftübertragung von den Luftpartikeln auf das Haar geschieht viskös, der Faden wird durch die Reibung der Luftpartikel am Haarschaft mitgenommen. Durch Reibung befördern feine Fiederungen am Haar diese Kraftübertragung. Bei den meisten Trichobothria beträgt das Amplitudenverhältnis zwischen Haarauslenkung und Luftpartikelbewegung 1:2 bis 1:1. Es gibt auch einzelne Trichobothria, bei denen die Haarauslenkung größer ist als die Luftpartikelbewegung. Die Haarauslenkung korreliert mit der Luftpartikelbeschleunigung und der Geschwindigkeit, letzteres nennt man auch Schallschnelle. Die Schallschnelle ist nur im Nahfeld bemerkbar, da sie mit der Entfernung sehr schnell abnimmt, realistisch sind Werte von bis zu 30 cm. Dieser sensorisch erfasste Raum ist bezogen auf die Körpergröße einer Spinne ausreichend.
Die Jagdspinne Cupiennius salei kann mit diesen Vibrationssensoren, die durch unterschiedliche Haarlängen insgesamt einen ausreichend weiten Bereich von Luftschwingungsstärken und -frequenzen rezipieren, ihre Beute ausmachen. So kann sie sogar fliegende Insekten aus der Luft fangen. Beispielsweise kann sie eine summende Fliege aus 30 cm und eine mit den Flügeln schlagende Fliege aus 70 cm Entfernung bemerken. Die Hauptnahrung der Spinnen sind auf den Blättern laufende Insekten, diese erzeugen in ihrem Umkreis von 20 mm Luftströmungen, auf die die Trichobothria reagieren, wodurch sich die Spinne in Richtung Beute orientiert.
Literatur
- Rainer F. Foelix: Biology of Spiders. Harvard University Press, Cambridge MA u. a. 1982, ISBN 0-674-07431-9.
- Pierre-P. Grassé: Traité de Zoologie. Anatomie, Systématique, Biologie. Tome 6: Onychophores – Tardigrades – Arthropodes – Trilobitomorphes – Chélicérates. Masson et Cie, Paris 1968.
- Jeffrey W. Shultz: Evolutionary Morphology And Phylogeny of Arachnida. In: Cladistics. Vol. 6, Nr. 1, 1990, ISSN 0748-3007, S. 1–38.
Weblinks
- Der Lufthauch, der die Fliege ankündigt, Bericht von Veronika Schmidt, Die Presse, 26. Mai 2012
Einzelnachweise
- Peter Görner: A proposed transducing mechanism for a multiply-innervated mechanoreceptor (trichobothrium) in spiders. In: Cold Spring Harbor Symposia on Quantitative Biology. Band 30. Cold Spring Harbor Laboratory Press, 1965, doi:10.1101/SQB.1965.030.01.010.
- Kazimir V. Drašlar: Functional properties of trichobothria in the bug Pyrrhocoris apterus (L.). In: Journal of Comparative Physiology 84, Nr. 2, 1973, S. 175–184.
- Aleš Škorjanc, Samo Batagelj, Kazimir Drašlar: Thermal acclimatization does not affect the resting activity of type T 1 trichobothrium in the firebug (Pyrrhocoris apterus; Heteroptera). In: Acta Biol Slov 50, Nr. 2, 2007, S. 85–92.
- J. Zrzavý: Antennal trichobothria in Heteroptera: a phylogenetic approach. In: Acta Entomologica Bohemoslovaca 87, Nr. 5, 1990, S. 321–325.
- Michael E. Soleglad, Victor Fet: Evolution of scorpion orthobothriotaxy: a cladistic approach. In: Euscorpius 2001, Nr. 1, 2013, S. 1–38.