Haarschwanzmaulwurf
Der Haarschwanzmaulwurf (Parascalops breweri) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Sein Verbreitungsgebiet erstreckt sich über den östlichen Teil von Nordamerika vom südlichen Kanada bis in die nordöstliche und östliche USA. Er kommt sowohl in Wäldern als auch in offenen Graslandschaften bis in mittlere Gebirgslagen vor. Die Tiere bevorzugen dabei Standorte mit lockeren, gut entwässerten Böden. Sie stellen mittelgroße Vertreter der Neuweltmaulwürfe dar, die wie alle anderen Angehörigen der Gruppe ebenfalls an eine unterirdische Lebensweise angepasst sind. Ihr Körper ist dadurch walzenförmig, der Hals kurz und die Vordergliedmaßen gleichen Grabschaufeln. Die Fellfärbung zeichnet sich durch einen dunklen Farbton aus, manchmal kommen hellere Flecken vor. Der deutschsprachige Trivialname der Art bezieht sich auf den fellbedeckten Schwanz.
| Haarschwanzmaulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
_(7748318398).jpg.webp)
Haarschwanzmaulwurf (Parascalops breweri) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Parascalops | ||||||||||||
| True, 1894 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Parascalops breweri | ||||||||||||
| (Bachman, 1841) |
Die Tiere leben unterirdisch und graben eigene Gang- und Tunnelsysteme, die sehr ausgedehnt sein können. Es bestehen mehrere Ebenen mit oberflächennahen Tunneln, die zumeist im Frühjahr und Sommer genutzt werden und der Nahrungssuche und Fortbewegung dienen. Tiefere Abschnitte werden häufig im Winter aufgesucht und schützen vorrangig vor Frost. Ebenso liegen die Nestkammern zum Rückzug und zur Aufzucht des Nachwuchses meist tiefer. Der Haarschwanzmaulwurf lebt einzelgängerisch, jedes Tier beansprucht ein eigenes Tunnelsystem. Zu bestimmten Jahresabschnitten können aber mehrere Individuen unterschiedlichen oder gleichen Geschlechts und verschiedener Altersstufen in den gleichen Gängen auftreten. Die Nahrung umfasst Regenwürmer und Insekten. Gelegentlich erfolgt die Nahrungssuche an der Erdoberfläche. Die Fortpflanzung ist auf das Frühjahr beschränkt. Der Wurf eines Weibchens besteht aus vier bis fünf Jungen, die rund einen Monat gesäugt werden.
Einzelne Berichte und Erwähnungen aus der zweiten Hälfte des 18. Jahrhunderts beziehen sich höchstwahrscheinlich auf den Haarschwanzmaulwurf. Die offizielle Erstbeschreibung datiert aber in das Jahr 1841. Der heute gültige Gattungsname wurde rund ein halbes Jahrhundert später eingeführt. Art und Gattung sind monotypisch. Es wurde aber zusätzlich ein ausgestorbener Vertreter benannt, der im Pliozän im östlichen Europa auftrat. Ebenso sind aus Nordamerika aus dieser Zeit fossile Reste bekannt. Der Bestand der heutigen Art wird als nicht gefährdet eingestuft.
Merkmale
Habitus
Der Haarschwanzmaulwurf repräsentiert einen mittelgroßen Vertreter der Neuweltmaulwürfe. Er erreicht eine Kopf-Rumpf-Länge von 12,5 bis 14,0 cm und eine Schwanzlänge von 2,6 bis 3,3 cm. Der Schwanz macht dadurch weniger als ein Viertel der Gesamtlänge aus. Das Gewicht variiert von 41,0 bis 62,8 g. Männchen sind im Durchschnitt rund 15 % größer als Weibchen, womit ein markanter Geschlechtsdimorphismus vorherrscht. Der Körperbau der Tiere zeigt wie bei allen Neuweltmaulwürfen Anpassungen an eine grabende Lebensweise. Der Rumpf ist walzenförmig, der Hals kurz und die Vordergliedmaßen sind zu Grabwerkzeugen umgebildet. Die Handflächen werden so lang wie breit und sind mit kräftigen, breiten und flachen Krallen ausgestattet. Diejenigen der Hinterfüße wirken dagegen schmal und kurz. Das Rückenfell besitzt eine schiefergraue bis schwarze Farbgebung, die Unterseite ist leicht heller getönt. Teilweise treten weiße Fleckenbildungen von rund 1 cm Durchmesser an Brust und Bauch auf. Die Füße sind fleischig und nur spärlich behaart. Den Schwanz, der an der Basis leicht einzieht, bedeckt ein dichtes Fell, außerdem besitzt er Schuppenringe. Generell ist das Fellkleid dicht und weich sowie teils seidig. Die Haare werden alle etwa gleich lang. Jungtiere zeichnen sich durch dunkelbraune Haare an den Füßen, am Schwanz und an der Schnauze aus, diese wechseln später in einen graufarbenen Ton über und werden zu fast weiß bei alten Individuen. Die Schnauze ist kurz und zugespitzt. Die Nasenlöcher liegen seitwärts und sind nach oben gerichtet. Die kleinen Augen bleiben im Fell verborgen. Sie sind in ihrer Größe stark reduziert, ihr Durchmesser beträgt rund 1 mm. Strukturell besitzen sie aber den Aufbau eines gewöhnlichen Säugetierauges. Ohrmuscheln fehlen, es kommen aber längliche Öffnungen von 3,5 mm Ausdehnung vor. Einzelne längere Haare an der Schnauze und hinter den Augen haben wohl die Funktion von Vibrissen. Die Hinterfußlänge liegt bei 1,6 bis 1,9 cm. Weibchen haben vier Paare an Zitzen, zwei davon befinden sich im Brust- und jeweils eines im Bauch- und Leistenbereich.[1][2][3][4]
Schädel- und Gebissmerkmale
Der Schädel wird 31,0 bis 33,7 mm lang und an den Warzenfortsätzen 14,0 bis 15,2 mm breit. Die Breite im Bereich der Jochbögen liegt bei 11,8 bis 12,5 mm. Allgemein ist der Schädel konisch geformt und flach, hinter den Augen tritt eine Eindellung auf. In Aufsicht zieht er hinter den Orbita etwas ein. Der Hirnschädel ist moderat breit. Das Rostrum wirkt schmal. Der Mittelkieferknochen überragt die Nasenbeine nach vorn und ist am Ende abgestumpft. Die Jochbögen sind relativ kräftig gebaut. An der Schädelbasis sind die Paukenblasen nur unvollständig entwickelt. Ein äußerer Gehörgang ist nicht sichtbar. Der schmale Gaumen endet kurz hinter dem letzten Molaren. Der horizontale Knochenkörper des Unterkiefers verläuft geschwungen, nach vorn neigt er abwärts, nach hinten richtet er sich aufwärts. Der Kronenfortsatz lehnt sich nach vorn. Er ist hoch und in Seitenansicht breit. Der Winkelfortsatz wirkt klein.[5][2][3][4]
Das Gebiss besteht aus 44 Zähnen, die Zahnformel lautet: . Manchmal treten überzählige Zähne auf. Beobachtet wurden bisher ein oder zwei zusätzliche Zahnbildungen. Sie sind nicht auf bestimmte Kieferseiten beschränkt, betreffen aber jeweils die Schneidezähne und Prämolaren. Sie ähneln in Größe und Form den regulären Zähnen.[6] Wie üblich bei den Neuweltmaulwürfen ist der vorderste obere Schneidezahn stark vergrößert. Die nachfolgenden Schneidezähne und die ersten drei Prämolaren werden nur halb so groß und weisen nur eine Spitze auf (unicuspid). Der Eckzahn ähnelt diesen, wird aber etwas größer. In der unteren Zahnreihe übertrifft der zweite Schneidezahn die anderen an Größe und hat eine eckzahnartige Gestalt (caniniform). Die übrigen Schneidezähne und die vorderen drei Prämolaren sind wiederum klein und einspitzig. Die jeweils letzte obere und untere Prämolar ist deutlich größer. Die oberen Molaren zeigen sich im Querschnitt W-förmig, die unteren M-förmig. Die Haupthöcker sind auf der Kaufläche der oberen Mahlzähne W-förmig angeordnet (dilambdodont). An der jeweiligen Zahninnenseite tritt ein niedriger Wulst auf, der aus drei Lappen besteht (trilobat), lediglich am letzten Molar sind es nur zwei Lappen (bilobat). An den unteren Molaren kommen einzelne zusätzliche Nebenhöckerchen vor. Die Länge der oberen Zahnreihe reicht von 13,0 bis 14,1 mm, die untere Prämolaren-Molarenreihe erstreckt sich über 8,9 bis 9,8 mm.[5][2][3][4]

Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 34. Er stimmt somit weitgehend mit dem der meisten anderen Neuweltmaulwürfe überein. Die Aufteilung der Autosomenpaare wird unterschiedlich angegeben, einerseits mit 12 zweiarmigen und 4 acrozentrischen, andererseits mit 15 zweiarmigen und 1 acrozentrischen. Demnach variiert auch die fundamentale Anzahl, also die Anzahl der Autosomenarme, zwischen 56 und 62.[7][8][9] Das X-Chromosom ist meta- bis submetazentrisch, das Y-Chromosom fleckenförmig.[7][2][4]
Verbreitung und Lebensraum
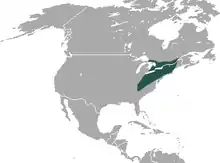
Das Verbreitungsgebiet des Haarschwanzmaulwurfs umfasst Teile des östlichen Nordamerikas. Es reicht im Norden bis in das südliche Kanada, wo er in den Provinzen Ontario und Québec auftritt. Nach Süden hin schließt es den nordöstlichen und östlichen Bereich der USA ein. Die Südgrenze liegt im Norden der US-Bundesstaaten Georgia und South Carolina. Im Osten wird ein Großteil der Küstenregionen ausgenommen, im Westen verläuft die Grenze durch das östliche Ohio, Kentucky und Tennessee. Die Tiere bewohnen verschiedene Lebensräume wie Wälder, Grasländer und Weiden, häufig in Verbindung mit mächtigen Humusschichten. Sie bevorzugen lockere und gut entwässerte, leichte Böden wie feuchte Sandlehme. In der Regel meiden sie harte, trockene oder reine Sandböden. Gelegentlich ist der Haarschwanzmaulwurf auch in Marschland oder stark tonigen Böden zu finden. Die Populationsdichte beträgt im Durchschnitt 3 Individuen auf einem Hektar, im Extremfall können es 25 bis 30 Tiere auf einer vergleichbar großen Fläche sein.[1][10] Die Höhenverbreitung reicht vom Meeresspiegelniveau bis auf 1950 m in den Great Smoky Mountains.[2][3][4]
Lebensweise und Ernährung
Territorialverhalten
Wie alle Neuweltmaulwürfe führt der Haarschwanzmaulwurf eine unterirdisch grabende Lebensweise. Er errichtet komplexe Gangsysteme unter der Erde. Ein im Oswego County im US-Bundesstaat New York vollständig untersuchtes System war 550 m lang, von denen 90 m unterhalb von Grasboden verliefen.[11] Die Gänge verteilen sich auf mehrere Ebenen. Oberflächennahe Tunnel bilden ein irreguläres Netzwerk und dienen der Nahrungssuche und Fortbewegung. Auf einer Distanz von 150 m wurden bei Untersuchungen bis zu 220 Gänge angeschnitten, was einen durchschnittlichen Abstand von etwa 67 cm entspricht. Die Tunnelröhren haben einen elliptischen Querschnitt von 3,7 bis 4,5 cm Breite und 2,5 bis 3,2 cm Höhe. Häufig verlaufen sie in rund 8 cm Tiefe, oberflächige Rippeln wie bei anderen grabenden Maulwürfen sind selten erkennbar. Teilweise orientieren sich die Gänge entlang natürlicher Objekte wie Steine oder Baumstämme. Sie besitzen regelmäßige Ausgänge, die auch von anderen Tieren wie Mäusen uns Spitzmäusen genutzt werden. Aktivitäten in den oberflächennahen Gängen erfolgen häufig im Frühjahr und Sommer. Tiefere Tunnel reichen 25 bis 45 cm in den Untergrund und sind seltener. Teilweise bedecken sie eine Fläche von 25 × 50 m. Sie fungieren als Rückzugsgebiete etwa bei frostigen Bedingungen im Winter. Ein größerer Teil der tieferen Gänge wird daher im Spätsommer und Herbst angelegt. In den tieferen Bereichen befinden sich auch Nestkammern von 15 cm Durchmesser, welche mit Pflanzenmaterial ausgestattet sind. Unter Umständen nutzt der Haarschwanzmaulwurf aber auch Tunnelerweiterungen von rund 8 cm Durchmesser als Ruheplatz. Beschädigte Bereiche der Gänge repariert ein Tier relativ zügig nach Entdeckung. Bei den Grabungsaktivitäten entstehen an der Erdoberfläche charakteristische Auswurfhügel (Maulwurfshügel). Sie weisen einen Durchmesser von 15 cm und eine Höhe von 7,5 cm auf. Auf einer Fläche von 15 m Durchmesser können bis zu 56 einzelne Hügel entstehen.[1] Unter Umständen sind die Tunnelsysteme über mehrere Maulwurfs-Generationen in Benutzung, einzelne Berichte geben bis zu acht Jahre an.[12] Der Haarschwanzmaulwurf ist generell weniger deutlich an die grabende Lebensweise angepasst als andere Vertreter der nordamerikanischen Neuweltmaulwürfe, wofür sein nicht so robuster Oberarmknochen im Vergleich etwa zu dem des Ostamerikanischen Maulwurfs (Scalopus) spricht. Dennoch ist er ein kräftiger Gräber der bis zu 370 g schwere Steine bewegen kann. Die Grabungsaktivitäten werden vornehmlich von der Bodenkonsistenz bestimmt. Die Grabgeschwindigkeit geht in härteren Substraten merklich zurück, von 37 m auf etwa 1,3 m je Stunde. Auch die Tunnellänge verringert sich. Dem gegenüber bewegt ein Tier in härteren Böden mehr Erdmaterial, was sowohl absolut als auch in Relation zum individuellen Körpergewicht und zur Arbeitsleistung je Stunde zutrifft. Der getätigte Aufwand ist teilweise um das bis zu Vierfache höher. Gleichzeitig verringert sich die kontinuierlich absolvierte Grabtätigkeit.[13][2][3][4]
Wie viele Maulwürfe hat der Haarschwanzmaulwurf keinen ausgeprägten Tag-Nacht-Rhythmus. Er ist zu beiden Tageszeiten aktiv, tendenziell häufiger jedoch tagsüber mit einer Kulmination von 08.00 bis 14.00 Uhr. Möglicherweise besteht aber ein individueller Tagesablauf. Die grabende Lebensweise bedingt, dass ein Großteil der Aktivitäten unterirdisch stattfindet. Doch kommt der Haarschwanzmaulwurf gelegentlich an die Erdoberfläche. Dies geschieht in der Regel während der Nacht, in seltenen Fällen sind auch tagsüber Oberflächenaktivitäten zu verzeichnen. Vor allem Jungtiere, die das mütterliche Nest verlassen, bewegen sich über bestimmte Strecken an der Erdoberfläche fort.[10][14] Des Weiteren verrichten die Tiere ihren Stuhlgang oberirdisch. Dazu suchen sie Latrinen auf, an denen sich bis zu einem Dutzend Kotballen ansammeln können. Die einzelnen Fäzes sind bis zu 10 mm lang, zylindrisch und an beiden Enden zugespitzt. Die typische Stellung der Vorderbeine mit nach außen gerichteten Handflächen führt zu eher unbeholfenen Bewegungen auf der Erdoberfläche. Gemessene Geschwindigkeiten liegen bei 4,6 m in 30 Sekunden, was umgerechnet etwa 550 m je Stunde entspricht.[1] Dafür ist der Haarschwanzmaulwurf ein guter Schwimmer. Hierbei bewegt er alle vier Gliedmaßen gleichzeitig, abweichend vom Ostamerikanischen Maulwurf, der nur die Hinterbeine einsetzt. Die Fortbewegung im wässrigen Milieu stellt aber eher eine Ausnahme dar.[15][2][3][4]
Im Allgemeinen lebt der Haarschwanzmaulwurf einzelgängerisch. Jedes Individuum beansprucht ein eigenes Tunnelsystem. Die Reviere der Tiere erstrecken sich über 810 m² im Durchschnitt, können im Sommer aber größer sein. Gelegentlich kommt es zum gemeinsamen Auftreten mehrerer Individuen. Dies trifft im Frühjahr vor allem auf Männchen zu. Im Sommer können Individuen beider Geschlechter und zudem Jungtiere in den gleichen Tunnelsystemen angetroffen werden. Es lassen sich nur selten Narben oder Verwundungen feststellen, so dass es möglicherweise nur in Einzelfällen zu antagonistischem Verhalten kommt.[1][2][3][4]
Ernährung
Die Nahrung des Haarschwanzmaulwurfs besteht aus Regenwürmern, verschiedenen Gliederfüßern, hier unter anderem Tausendfüßer und Hundertfüßer, sowie Insekten. Die prinzipielle Zusammensetzung der Nahrung war schon durch Untersuchungen Ende des 19. Jahrhunderts bekannt geworden.[16] Anhand von 100 analysierten Mageninhalten aus New Hampshire ließ sich ein Volumenanteil der Regenwürmer von 34 % feststellen, sie kamen in 84 % aller untersuchten Proben vor. Die entsprechenden Werte für Insektenlarven betrugen 29 % und 96 %, für ausgewachsene Insekten wiederum 18 % und 86 %. Tausend- und Hundertfüßer erreichten einen Volumenanteil von 2 % und einen Mengenanteil von 32 %. Darüber hinaus traten Schnecken, Asseln und verschiedenes pflanzliches Material wie auch Sand und Detritus auf. Unter den Insekten waren Käfer recht häufig, so beispielsweise Schnellkäfer, Weichkäfer, Blatthornkäfer, Rüsselkäfer, Laufkäfer und Schwarzkäfer. Daneben ließen sich Zweiflügler mit Schaben, Pilzmücken und Raubfliegen belegen. Untergeordnet traten zudem Schmetterlinge auf. Ameisen konnten in 57 % aller Proben dokumentiert werden. Ihre Häufigkeit nahm jeweils zu Beginn des Frühjahrs und im ausklingenden Herbst zu, möglicherweise zu einem Zeitpunkt, zu dem andere Nahrungsquellen rar sind.[1] Weitere 29 Mageninhalte aus dem Bundesstaat New York bestätigen diese Zusammensetzung weitgehend. Sie umfassten Regenwürmer, Ameisen, Käferlarven, Hundertfüßer und Pflanzenwurzeln.[17] Ähnlich verhält es sich mit rund einem Dutzend Magenproben aus Pennsylvania. Bei diesen nahmen Insekten allerdings mehr als die Hälfte und sonstige Gliederfüßer ein weiteres Viertel der Gesamtnahrung ein.[18] Gelegentliche Beobachtungen zeigen, dass einzelne Tiere Nester von Wespen plündern, wobei die Insekten durch übergeworfene Erde bedeckt werden.[19][2][3][4]
Wie alle Neuweltmaulwürfe sucht der Haarschwanzmaulwurf seine Nahrung in der Regel unterirdisch. Unter Umständen kommt er dafür jedoch auch an die Erdoberfläche, was meist in den frühen Morgen- und Abendstunden geschieht. Nach Freilandbeobachtungen in den Appalachen können diese oberflächigen Aktivitäten über eine halbe Stunde andauern. In dieser Zeit legt ein Tier bis zu 27 m zurück und gräbt kleine, bis zu 6 cm tiefe Löcher. Teilweise bewegt es sich auch durch dicke Laubschichten.[20][14] Die Regenwürmer werden durch das Abbeißen der vorderen Segmente betäubt und anschließend deponiert. Bei großen Individuen quetscht der Haarschwanzmaulwurf mit den Vorderfüßen das Erdmaterial aus den Verdauungsorganen der Würmer. In einzelnen Experimenten fraßen Tiere von 41 bis 50 g Körpergewicht täglich zwischen 66 und 159 g an Wirbellosen. Dies entspricht in etwa dem 1,3 bis 3,9-fachen ihres eigenen Körpergewichts. In menschlicher Gefangenschaft vertilgt der Haarschwanzmaulwurf auch Eier, Fleisch und Mehlwürmer, meidet jedoch pflanzliches Material wie Kartoffeln.[16][21][2][3][4]
Fortpflanzung
Die Paarung findet im Frühjahr statt und erfolgt vermutlich nur einmal jährlich. In New Hampshire wurde dies für den Zeitraum vom späten März bis frühen April beobachtet. Während dieser Zeit schwellen die Hoden der Männchen stark an, von 3 × 2 mm außerhalb der Fortpflanzungsphase auf 12 × 7 mm während der Reproduktion. Auch die Prostata vergrößert sich deutlich. Bei den Weibchen öffnet sich die Vagina, die ansonsten von einer Haut überdeckt ist. Die direkte Paarung wurde bisher nicht beobachtet, ebenso ist die Tragzeit unbekannt, es werden jedoch vier bis sechs Wochen vermutet. Während dieser Zeit legt das werdende Muttertier eine Nestkammer an. Diese befindet sich rund 25 bis 30 cm unter der Erdoberfläche. Die Kammer enthält ein rundes Nest von etwa 16 cm Durchmesser, das nahezu vollständig aus Blättern in mehreren dicht gepackten und konzentrischen Lagen besteht. Der innere Durchmesser der Kammer beträgt 8 cm. Der Wurf eines Weibchens umfasst für gewöhnlich vier oder fünf Jungtiere, es wurden aber bis zu acht Embryonen berichtet.[18] Neugeborene sind rund 71 mm lang und 10 g schwer. Sie kommen blind und nackt zur Welt, die Haut ist gefaltet und weißlich. Lediglich einzelne Vibrissen wachsen an der Schnauze. Augen- und Ohröffnungen sind erkennbar. Die Krallen haben eine weiche Struktur, das Maul ist zahnlos. Der Nachwuchs wird rund einen Monat gesäugt, danach nehmen die Jungen feste Nahrung zu sich. Zu diesem Zeitpunkt wiegen sie zwischen 34 und 51 g und erreichen somit fast das Gewicht ausgewachsener Individuen. Sie ähneln äußerlich auch diesen, sind aber etwas gräulicher in der Farbgebung des Fells. Die Geschlechtsreife setzt vermutlich nach rund zehn Monaten ein. Unter Umständen sind in diesem Alter noch Teile des Milchgebisses erhalten.[6] Anhand der Zahnabnutzung wird eine Lebenserwartung von rund vier Jahren geschätzt.[1][2][3][4]
Fressfeinde, Kommensalen und Parasiten
Der Haarschwanzmaulwurf wird nur selten von Beutegreifern erlegt. Einzelne Untersuchungen in Massachusetts ergaben einen Anteil von rund 0,75 % der Art in 131 Kotresten des Rotfuchses.[22] Ebenfalls selten fand sie sich in Rotfuchs-Fäzes in New Hampshire.[1] Reste des Haarschwanzmaulwurfs wurden darüber hinaus zusammen mit Überbleibseln von Fledertieren und Insekten im Mageninhalt eines Nordopossums registriert.[23] Weitere bedeutende Fressfeinde bilden der Bartkauz und die Schleiereule. Bei letzterer erbrachte die Analyse von mehr als 2480 Gewöllen aus Pennsylvania lediglich einen Anteil des Haarschwanzmaulwurfs von 1,2 %.[18] Unter den Reptilien sind Schlangen wie der Nordamerikanische Kupferkopf zu nennen, was Untersuchungen von entsprechenden Exemplaren in Virginia erbrachten.[24] Als mögliche Kommensalen treten die Amerikanischen Kurzschwanzspitzmäuse auf, die die Tunnel und Gänge des Haarschwanzmaulwurfes nutzen.[18] Andere assoziierte Arten gehören zu den Rotzahnspitzmäusen und den Feldmäusen.[1][2]
Es sind zahlreiche Parasiten des Haarschwanzmaulwurfs dokumentiert. Zu den häufigsten äußeren zählen Flöhe und Milben. Einzelne Individuen können bis zu drei Dutzend und mehr Parasiten tragen, besonders häufig tritt dies im Frühjahr und im Sommer in Erscheinung.[1] Eine Studie aus dem Jahr 1988 an sieben Individuen des Haarschwanzmaulwurfs erbrachte insgesamt 33 verschiedene Floh- und Milbenarten.[25] Flöhe sind mit Androlaelaps, Ctenophthalmus, Doratopsylla, Hystrichopsylla, Megabothris und Nearctopsylla belegt. Zu den Milben gehören unter anderem Cyrtolaelaps, Echinonyssus, Euryparasitus, Haemogamasus, Hirstionyssus, Hypoaspis, Laelaps und Proctolaelaps aus der Gruppe der Raubmilben. Die individuell zahlreichste Form bildet Labidophorus aus der Gruppe der Astigmata. Dem gegenüber stellt Pygmaephorus aus der Gruppe der Prostigmata die formenreichste Gattung dar, sie ist nicht als Parasit im engeren Sinne anzusehen. Außerdem wurden mit Euhaematopinus und Haematopinoides Angehörige der Läuse und mit Leptinus ein solcher der Käfer verzeichnet.[1][26][27][28][29][25] Innere Parasiten umfassen vor allem Würmer. Hier sind mit Physaloptera und Porrocaecum zwei Formen der Fadenwürmer sowie mit Macracanthorhynchus und Moniliformis zwei der Kratzwürmer zu nennen. Einigen Erhebungen zufolge befallen die Fadenwürmer bis zu 15 % aller Maulwurf-Individuen, in manchen Fällen wurden bis zu drei Dutzend Würmer in einem Magen angetroffen.[1][30] Des Weiteren ist der Haarschwanzmaulwurf Träger verschiedener Kokzidien wie Cyclospora, Eimeria und Isospora. Diese können je nach Form bei einem Viertel bis zur Hälfte aller Individuen auftreten.[31][2]
Systematik
Innere Systematik der Neuweltmaulwürfe nach Chen et al. 2021[32]
|
Der Haarschwanzmaulwurf ist eine Art aus der Gattung Parascalops, innerhalb der er deren einziges rezentes Mitglied darstellt. Die Gattung wird somit aus heutiger Sicht als monotypisch eingestuft. Art und Gattung gehören wiederum zur Familie der Maulwürfe (Talpidae) und dieser untergeordnet zusammen mit einigen weiteren in Asien und Nordamerika verbreiteten Formen zur Tribus der Neuweltmaulwürfe (Scalopini). Die Neuweltmaulwürfe umfassen, ähnlich den Eigentlichen Maulwürfen (Talpini), grabende Angehörige der Familie, die beiden Gruppen sind jedoch nicht unmittelbar verwandt, sondern entwickelten ihre Grabeigenschaften unabhängig. Andere Familienmitglieder wiederum leben nur teilweise unterirdisch, bewegen sich oberirdisch fort oder sind an eine semi-aquatische Lebensweise angepasst.[33] Besondere Kennzeichen der Neuweltmaulwürfe finden sich in dem im Vergleich zu den Eigentlichen Maulwürfen deutlich längeren Schwanz, in dem stark vergrößerten vorderen oberen Schneidezahn und in dem auftretenden Zahnwechsel. Entsprechend den Eigentlichen Maulwürfen ist an der Hand ein zusätzliches Sesambein vor dem Daumen ausgebildet, der sogenannte Präpollex („Vordaumen“), wodurch die Handfläche verbreitert wird.[34][4] Laut molekulargenetischen Untersuchungen spalteten sich die Neuweltmaulwürfe im Oberen Eozän vor rund 39 bis 35 Millionen Jahren von den anderen Triben der Maulwürfe ab.[33] Die Tribus gliedert sich in zwei Entwicklungslinien: die Parascalopina und die Scalopina, die beide anhand der Ausprägung des Metastylids am unteren zweiten Molar unterscheidbar sind. Bei den Scalopina fehlt das Metastylid, bei den Parascalopina hingegen kommt es vor.[35][36] Die beiden Linien bestehen wenigstens seit dem Unteren Miozän vor 21,4 Millionen Jahren als eigenständig. Der Haarschwanzmaulwurf ist die Charakterform der Parascalopina, als nächste Verwandte gelten der Gansu-Maulwurf (Scapanulus) und die Gattung Alpiscaptulus, die jeweils in Ostasien vorkommen. Die Diversifizierung des nordamerikanischen Haarschwanzmaulwurfs von den asiatischen Formen fand im frühen Mittleren Miozän vor gut 17 Millionen Jahren statt.[37][32]
Es werden keine Unterarten des Haarschwanzmaulwurfs unterschieden, wie die Gattung ist somit auch die Art monotypisch.[38][4][39] Es besteht aber eine weitere fossile Art:[40]
- Parascalops fossilis Skoczeń, 1993
Die Artbeschreibung basiert auf Material zweier Fundstellen in Polen. Bereits 1980 hatte Stanisław Skoczeń einen Teil der Funde unter der Bezeichnung Scapanulus agrarius vorgestellt, womit er sie in die Nähe des Gansu-Maulwurfs verwies.[41] Dies stieß aber in der Folgezeit auf einzelne Kritiken, da zu deutliche Abweichungen zum heutigen Gansu-Maulwurf vorliegen, etwa in der gegenüber letzterem schwach ausgeprägten scalopine ridge, einem charakteristischen Knochenrücken am oberen Gelenkende des Oberarmknochens.[42] Daher unterzog Skoczeń das Material einer Revision und veröffentlichte es 1993 unter dem heute akzeptierten Artnamen Parascalops fossilis. Sowohl der rezenten wie auch der fossilen Art ist die schwach ausgebildete scalopine ridge zu eigen.[43] Eine im Jahr 2001 von Thomas Dahlmann verwendete Bezeichnung Scalopoides agrarius mit Verweis auf Skoczeń 1980 für einzelne Funde aus Deutschland ist durch deren Fragmentierung schwer beurteilbar, da der Bereich der scalopine ridge bei keinem der Objekte erhalten blieb.[44] Scalopoides stellt eine nordamerikanische Fossilform des Miozäns dar, deren heute gültiger Gattungsname allerdings Mioscalops lautet.[45][36] Einzelne Autorn nehmen an, dass das Material aus Deutschland ebenfalls zu Parascalops fossilis gehört.[40]
Forschungsgeschichte


Der Haarschwanzmaulwurf wurde wahrscheinlich schon im Verlauf des 18. Jahrhunderts bekannt. Der walisische Naturforscher Thomas Pennant stellte in seiner Synopsis of Quadrupeds im Jahr 1771 einen longtailed mole („langschwänziger Maulwurf“) in Wort und Bild vor, der sich vor allem durch die kurze Behaarung auf dem Schwanz auszeichnete. Als Herkunftsgebiet wies Pennant lediglich Nordamerika aus.[46] Darauf basierend vergab im Jahr 1777 Johann Christian Polycarp Erxleben den wissenschaftlichen Namen Talpa longicaudata,[47] gleichzeitig führte Eberhard August Wilhelm von Zimmermann die Bezeichnung Talpa caudata ein.[48] Beide hoben ebenfalls Nordamerika als Herkunftsgebiet hervor, bei Erxleben mit America boreali, bei Zimmermann mit America septentrionali bezeichnet. Vor allem Erxlebens Talpa longicaudata wurde in der Folgezeit mehrfach verwendet und findet sich unter anderem bei Pieter Boddaert 1784 (als Talpa longicauda),[49] aber auch bei Anselme Gaëtan Desmarest 1819 und 1820 (als Condylura longicaudata).[50][51] Lange Zeit war unklar, auf welchen Maulwurf sich Pennants Beschreibung bezog. Die Angabe von Desmarest als Condylura und damit Vertreter der Gattung des Sternmulls ist bereits wenige Jahre zuvor bei Johann Karl Wilhelm Illiger zu finden.[52] Auch Hartley H. T. Jackson teilte im Jahr 1915 in einer Revision der nordamerikanischen Maulwürfe diese Meinung. Er begründete dies mit der irrtümlichen Feststellung, Pennant habe in seiner Beschreibung des longtailed mole auf die fingerförmigen Hautanhänge an der Schnauze hingewiesen.[5] Im Gegensatz dazu setzte Joel Asaph Allen im Jahr 1902 den longtailed mole mit dem Ostamerikanischen Maulwurf (Scalops) gleich.[53][39]
Ein weiterer früher Name für den Haarschwanzmaulwurf wird häufig mit William Bartram verbunden. Der amerikanische Naturforscher führte akribisch Tagebuch über seine Beobachtungen in der Natur und notierte darin auch eine Art Talpa americana. Diese wurde von ihm zwar nie publiziert, gelangte aber 1825 über Richard Harlans Fauna Americana an die Öffentlichkeit. Harlan erwähnte dabei zwar den typischen behaarten Schwanz, doch stellt seine gesamte Beschreibung eine mehr oder weniger originalgetreue Übersetzung von Desmarests Abhandlungen zum Europäischen Maulwurf (Talpa europaea) aus dem Jahr 1820 dar.[54][55][5] In seinem Werk unterschied Harlan außerdem Pennants longtailed mole als Condylura longicaudata.[56] Talpa americana wird daher heute als Nomen nudum angesehen.[39]


Die wissenschaftlich anerkannte Erstbeschreibung stammt von John Bachman. Häufig wird als Jahr 1842 angegeben, aber bereits ein Jahr zuvor hatte Bachman in einem Aufsatz über die nordamerikanischen Maulwürfe die Art Scalops breweri vorgestellt. Wie zuvor Pennant und Harlan lenkte Bachman besondere Aufmerksamkeit auf den Schwanz, den er als biberartig platt und behaart charakterisierte. Das Artepitheton vergab er nach Thomas Mayo Brewer, einem Naturforscher aus Boston, der ihn auf die neue Art aufmerksam gemacht hatte. Als Typuslokalität benannte Bachman mit Martha's Vineyard eine Insel vor der Küste von Neuengland, genauer des US-Bundesstaates Massachusetts.[57][58] Die Artbezeichnung Scalops breweri stellte die neue Form an die Seite des Ostamerikanischen Maulwurfs. Von Auguste Pomel stammt im Jahr 1848 der Verweis in die von ihm neu geschaffene Gattung Scapanus, die die Westamerikanischen Maulwürfe umfasst und die er aufgrund der vollständigen Bezahnung vom Ostamerikanischen Maulwurf abhob.[59] Frederick William True führte dann im Jahr 1894 die Gattung Parascalops ein. Als Begründung benannte er neben der auffallenden Schwanzbehaarung unter anderem einzelne abweichende Merkmale in der Zahn- und Beckengestaltung.[60] Der Gattungsname setzt sich aus der griechischen Vorsilbe παρά (pará) für „neben“ oder „nahe bei“ und der Gattungsbezeichnung Scalops (Scalopus) für den Ostamerikanischen Maulwurf zusammen.[61] Die Gattung wurde nachfolgend mehrfach revidiert, so von True selbst zwei Jahre später,[55] von Hartley H. T. Jackson im Jahr 1915[5] und von Neal Woodman im Jahr 2018.[39]
Stammesgeschichte
Die Neuweltmaulwürfe verfügen über einen recht umfangreichen Fossilbericht, der sich auf die Alte und Neue Welt verteilt. Dies trifft auch auf die Gattung Parascalops zu. Die ältesten Funde gehören dem Unteren Pliozän an und sind damit über 4 Millionen Jahre alt. Sie wurden an der Gray fossil site im Washington County im US-Bundesstaat Tennessee entdeckt und bestehen aus einzelnen Oberarmknochen, Teilen des Unterarms und Fingergliedern. Allgemein ähneln die Elemente jenen des heutigen Haarschwanzmaulwurfs, zeigen aber einzelne morphologische Unterschiede. Eine neue Art wird als wahrscheinlich angesehen, wurde aber noch nicht benannt. Die Tiere lebten in einer einst wasserreichen Karstlandschaft mit einer vielgestaltigen Fauna und Flora.[62] Weitere Fossilreste sind dann in Nordamerika erst wieder aus dem Oberen Pleistozän belegt. Ein nahezu vollständiger Schädel und einzelne Reste des Vorderbeins gehören wahrscheinlich zum heutigen Haarschwanzmaulwurf und kamen aus der Frankstown Cave im Blair County in Pennsylvania zu Tage. Der Felshohlraum wurde im Jahr 1907 in einem Kalkstein-Steinbruch entdeckt, er barg ebenfalls eine umfangreiche Tierwelt.[63]
Im Pliozän war die Gattung Parascalops auch in Europa präsent. Darauf verweisen einzelne Oberarmknochen und Sprungbeine sowie ein Brustbein von Podlesice und Węże, zwei Höhlenfundstellen im südlichen Polen. Die Funde sind etwas kleiner als die entsprechenden Knochen des heutigen Haarschwanzmaulwurfs und werden der Art Parascalops fossilis zugewiesen. Eventuell lassen sich auch einzelne beschädigte Oberarmknochen und ein zusätzliches Unterkieferfragment aus Wölfersheim in Hessen zu dieser Art zuordnen.[41][43][40]
Bedrohung und Schutz
Die IUCN führt den Haarschwanzmaulwurf in der Kategorie „nicht gefährdet“ (least concern). Es sind keine größeren Bedrohungen für den Bestand bekannt. Des Weiteren gilt die Art allgemein als relativ häufig. Lokal können aber Unterschiede auftreten. So ist der Haarschwanzmaulwurf in Tennessee und in Georgia gefährdet. Die Tiere kommen in mehreren Naturschutzgebieten vor.[64][4]
Literatur
- James G. Hallett: Parascalops breweri. Mammalian Species 98, 1978, S. 1–4
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 598–599) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- John O. Whitaker und William John Hamilton: Mammals of the Eastern United States. Cornell University Press, 1998, S. 64–66
Einzelnachweise
- W. Robert Eadie: A contribution to the biology of Parascalops breweri. Journal of Mammalogy 20 (2), 1939, S. 150–173
- James G. Hallett: Parascalops breweri. Mammalian Species 98, 1978, S. 1–4
- John O. Whitaker und William John Hamilton: Mammals of the Eastern United States. Cornell University Press, 1998, S. 64–66
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 598–599) ISBN 978-84-16728-08-4
- Hartley H. T. Jackson: A review of the American moles. North American Fauna 38, 1915, S. 1–100 (S. 77–82)
- W. Robert Eadie: Tooth replacement in brewer's mole. The Anatomical Record 89 (3), 1944, S. 357–360
- Terry L. Yates und David J. Schmidly: Karyotype of the Eastern mole (Scalopus aquaticus), with comments on the karyology of the family Talpidae. Journal of Mammalogy 56 (4), 1975, S. 902–905
- J. W. F. Reumer und A. Meylan: New developments in vertebrate cytotaxonomy IX, Chromosome numbers in the order Insectivora (Mammalia). Genetica 70, 1986, S. 119–151
- Shin-ichiro Kawada, Masashi Harada, Kazuhiru Koyasu und Sen-ichi Oda: Karyological note on the short-faced mole, Scaptochirus moschatus (Insectivore, Talpidae). Mammal Study 27, 2002, S. 91–94, ()
- W. J. Hamilton, Jr.: Activity of Brewer's mole (Parascalops breweri). Journal of Mammalogy 20 (3), 1939, S. 307–310
- G. C. Hickman: Burrow structure of the Talpid mole Parascalops breweri from Oswego County, New York State. Zeitschrift für Säugetierkunde 48, 1983, S. 265–269 ()
- Philip L. Wright: Parascalops tunnel in use after eight years. Journal of Mammalogy 26 (4), 1945, S. 438–439
- Y.‐F. Lin, A. Chappuis, S. Rice und E. R. Dumont: The effects of soil compactness on the burrowing performance of sympatric eastern and hairy‐tailed moles. Journal of Zoology 301 (4), 2017, S. 310–319, doi:10.1111/jzo.12418
- Erin E. Fraser und Johnston F. Miller: Diurnal Above-ground Movement in Hairy-tailed Moles, Parascalops breweri. Canadian Field Naturalist 122, 2008, S. 267
- Root Gorelick und Susan M. Bertram: Swimming Eastern Chipmunks, Tamias striatus, and Hairy-tailed Mole, Parascalops breweri, in Kawartha Highlands Provincial Park, Ontario. Canadian Field Naturalist 122, 2008, S. 73–75
- Fred E. Brooks: Notes on the habits of mice, moles and shrews. West Virginia Agricultural and Forestry Experiment Station Bulletins 113, 1908, S. 87–133
- W. J. Hamilton, Jr.: The food of small forest mammals in eastern United States. Journal of Mammalogy 22 (3), 1941, S. 250–263
- Neil D. Richmond und Harry R. Rosland: Mammal survey of Northwestern Pennsylvania. Pennsylvania Game Commission and U. S. Fish and Wildlife Service, 1949, S. 1–67 (S. 34–36) ()
- Fred E. Brooks: Moles destroy wasps' nests. Journal of Mammalogy 4 (3), 1923, S. 183
- Gary R. Graves: Crepuscular surface foraging of the Hairy-tailed mole (Parascalops breweri). Journal of the North Carolina Academy of Science 118 (3), 2002, S. 206–209
- Francis H. Fay: Quantitative experiments on the food consumption of Parascalops breweri. Journal of Mammalogy 35 (1), 1954, S. 107–109
- W. J. Hamilton, Jr., N. W. Hosley und A. E. MacGregor: Late summer and early fall foods of the Red fox in Central Massachusetts. Journal of Mammalogy 18 (3), 1937, S. 366–367
- Charles C. Sperry: Opossum and skunk eat bats. Journal of Mammalogy 14 (2), 1933, S. 152–153
- Lawrence W. Saylor: Hairy-tailed mole in Virginia. Journal of Mammalogy 19 (2), 1938, S. 247
- John O. Whitaker, Jr. und Thomas W. French: Ectoparasites and Other Arthropod Associates of the Hairy-tailed Mole, Parascalops Breweri. The Great Lakes Entomologist 21 (1), 1988, S. 39–41
- E. W. Jameson, Jr.: Notes on the habits and siphonapterous parasites of the mammals of WeIland County, Ontario. Journal of Mammalogy 24 (2), 1943, S. 194–197
- E. W. Jameson, Jr.: The external parasites of the short-tailed shrew, Blarina brevicauda (Say). Journal of Mammalog 31 (1), 1950, S. 138–145
- A. Fain und John O. Whitaker, Jr.: Phoretic hypopi of North American mammals (Acarina: Sarcoptiformes, Glycyphagidae). Acarologia 15, 1973, S. 144–170
- Terry L. Yates, Danny B. Penc und Glenn K. Launchbaugh: Ectoparasites from seven species of North American moles (Insectivora: Talpidae). Journal of Medical Entomology 16, 1979, S. 166–168
- John M. Kinsella und Vasyl V. Tkach: Checklist of helminth parasites of Soricomorpha (= Insectivora) of North America north of Mexico. Zootaxa 1969, 2009, S. 36–58
- Paulette L. Ford und Donald W. Duszynski: Coccidian Parasites (Apicomplexa: Eimeriidae) from Insectivores. VII. Six New Species from the Hairy-Tailed Mole, Parascalops breweri. Journal of Parasitology 75 (4), 1989, S. 508–513
- Zhong-Zheng Chen, Shui-Wang He, Wen-Hao Hu, Wen-Yu Song, Kenneth O Onditi, Xue-You Li und Xue-Long Jiang: Morphology and phylogeny of scalopine moles (Eulipotyphla: Talpidae: Scalopini) from the eastern Himalayas, with descriptions of a new genus and species. Zoological Journal of the Linnean Society, 2021, S. zlaa172, doi:10.1093/zoolinnean/zlaa172
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang, Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumenting the polydactyly ‚constraint‘: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- J. Howard Hutchinson: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117
- Achim Schwermann, Kai He, Benjamin J. Peters, Thorsten Plogschties und Gabrielle Sansalone: Systematics and macroevolution of extant and fossil scalopine moles (Mammalia, Talpidae). Palaeontology 62 (4), 2019, S. 661–676, doi:10.1111/pala.12422
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Neal Woodman: American Recent Eulipotyphla. Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithsonian Institution Scholary Press, 2018, S. 1–107 (S. 19–20)
- Barbara Rzebik-Kowalska: Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland. Palaeontologia Electronica 17 (2), 2014, S. 17.2.26A, doi:10.26879/457
- Stanisław Skoczeń: Scaptonychini Van Valen, 1967, Urotrichini and Scalopini Dobson, 1883 (Insectivora, Mammalia) in the Pliocene and Pleistocene of Poland. Acta zoologica cracoviensia 24, 1980, S. 411–448
- Gerhard Storch und Z. Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Stanisław Skoczeń: New records of Parascalops, Neurotrichus and Condylura (Talpinae, Insectívora) from the Pliocene of Poland. Acta Theriologica 38 (2), 1993, S. 125–137
- Thomas Dahlmann: Die Kleinsäuger der unter-pliozänen Fundstelle Wölfersheim in der Wetterau (Mammalia: Lipotyphla, Chiroptera, Rodentia). Courier Forschungsinstitut Senckenberg 227, 2001, S. 1–129 (S. 21–22)
- Gregg E. Ostrander, Assefa Mebrate und Robert W. Wilson: Type and figured specimens of fossil vertebrates in the collection of the University of Kansas Museum of Natural History. Part IV. Fossil mammals. University of Kansas Museum of Natural History Miscellaneous Publication 79, 1986, S. 1–83 (S. 9) ()
- Thomas Pennant: Synopsis of Quadrupeds. Chester, 1771, S. 1–382 (S. 314) ()
- Johann Christian Polycarp Erxleben: Systema Regni Animalis per Classes, Ordines, Genera, Species, Varietates: cvm Synonymia et Historia Animalivm: Classis I.Mammalia. Leipzig, 1777, S. 1–636 (S. 118–119) ()
- Eberhard August Wilhelm von Zimmermann: Specimen Zoologiae Geographicae, Quadrupedum Dimicilia et Migrationes Sistens. Leiden, 1777, S. 1–685 (S. 497) ()
- Pieter Boddaert: Elenchus animalium. Volume I. Rotterdam, 1785, S. 1–174 (S. 126) ()
- Anselme Gaëtan Desmarest: Sur le genre Condylure d'Illiger. Journal de physique, de chimie, d'histoire naturelle et des arts 89, 1819, S. 225–232 ()
- Anselme Gaëtan Desmarest: Mammalogie, ou, Description des espèces de mammifères. Premiere Partie. Paris, 1820, S. 1–276 (S. 158–159) ()
- Johann Karl Wilhelm Illiger: Prodromus Systematis Mammalium et Avium. Berlin, 1811, S. 1–301 (S. 125–126) ()
- Joel Asaph Allen: Zimmermann’s „Zoologiae Geographicae“ and „Geographische Geschichte“ Considered in Their Relation to Mammalian Nomenclature. Bulletin of the American Museum of Natural History 16 (2), 1902, S. 13–22
- Frederick W. True: The proper scientific name for Brewer's mole. Science, N. S. 1, 1895, S. 101
- Frederick W. True: A revision of the American moles. Proceedings of the United States National Museum 19 (1101), 1896, S. 1–112 (S. 67–77)
- Richard Harlan: Fauna americana: being a description of the mammiferous animals inhabitating North America. Philadelphia, 1825, S. 1–325 (S. 43–44) ()
- John Bachman: Observations on the genus Scalops, with descriptions of the species found in North America. Proceedings of the Boston Society of Natural History 1, 1841, S. 40–41 ()
- John Bachman: Observations on the genus Scalops, (shrew moles,) with descriptions of the species found in North America. Boston Journal of Natural History 4, 1843–1844, S. 26–35 ()
- Auguste Pomel: Etudes sur les carnassiers insectivores (Extrait). Archives des sciences physiques et naturelles, Geneve 1 (9), 1848, S. 244–251 ()
- Frederick W. True: Diagnoses of new North American mammals. Proceedings of the United States National Museum 17 (999), 1894, S. 241–243 ()
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 514 ()
- Danielle Oberg: Fossil Moles from the Gray F ossil Site, TN: Implications for Diversification and Evolution of North American Talpidae. East Tennessee State University, 2018, S. 1–95
- O. A. Peterson: The fossils of the Frankstown Cave, Blair County, Pennsylvania. Annals of the Carnegie Museum 16, 1926, S. 249–314 ()
- F. Cassola: Parascalops breweri (errata version published in 2017). The IUCN Red List of Threatened Species 2016. e.T41469A115188181 (); zuletzt aufgerufen am 16. Mai 2021
Weblinks
- Parascalops breweri in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: F. Cassola, 2016. Abgerufen am 16. Mai 2021.