Kalifornischer Maulwurf
Der Kalifornische Maulwurf (Scapanus latimanus) ist eine Säugetierart aus der Gattung der Westamerikanischen Maulwürfe innerhalb der Maulwürfe (Talpidae). Sein Verbreitungsgebiet umfasst die Westküste Nordamerikas, wo er vom südlichen Oregon bis in den Süden von Kalifornien auftritt. Dort bewohnt er Landschaften mit offener oder geschlossener Vegetation auf feuchten Böden. Die Lebensräume reichen von Tieflandgebieten bis in höhere Gebirgslagen. Es handelt sich um einen mittelgroßen Vertreter der Westamerikanischen Maulwürfe. Wie seine Verwandten ist der Kalifornische Maulwurf an eine grabende Lebensweise angepasst. Er zeichnet sich durch einen walzenförmigen Körper mit kurzem Hals und grabschaufelartigen Vordergliedmaßen aus. Das Fell ist schwarzbraun bis silbergrau gefärbt.
| Kalifornischer Maulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
_(6-8-09)_canet_rd%252C_morro_bay%252C_slo_co%252C_ca_-01_(3609050224).jpg.webp)
Kalifornischer Maulwurf (Scapanus latimanus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Scapanus latimanus | ||||||||||||
| (Bachman, 1841) |
Über die Lebensweise liegen nur bedingt Informationen vor. Die Tiere legen unterirdische Tunnel und Gänge an, die in mehreren Ebenen verlaufen und pflanzengepolsterte Nester enthalten. Die Ernährung basiert auf Wirbellosen. Der Nachwuchs kommt wahrscheinlich einmal jährlich zur Welt. Die Art wurde im Jahr 1841 wissenschaftlich eingeführt, galt aber im Verlauf des 19. Jahrhunderts als identisch mit dem Townsend-Maulwurf. Diesbezüglich gab es auch eine Zeitlang Unklarheiten über den genauen Fundort der Belegexemplare. Die Stammesgeschichte reicht bis in das Altpleistozän zurück, der größere Teil der Fossilfunde entstammt dem Jungpleistozän. Heute sind mehrere Unterarten anerkannt. Der Bestand wird als nicht gefährdet eingestuft.
Merkmale
Habitus
_(6-8-09)_canet_rd%252C_morro_bay%252C_slo_co%252C_ca_-03_(3608236851).jpg.webp)
Der Kalifornische Maulwurf ist ein mittelgroßer Vertreter der Westamerikanischen Maulwürfe. Seine Kopf-Rumpf-Länge variiert von 13,1 bis 15,3 cm. Der Schwanz wird 2,3 bis 3,8 cm lang. Das Gewicht reicht von 55,6 bis 73,9 g, sehr große Individuen können auch bis zu 85,0 g wiegen. Es gibt starke geographische Unterschiede. Tiere aus dem trockenen südlichen Verbreitungsgebiet sind deutlich kleiner als solche aus dem feuchten nördlichen und östlichen. Zudem besteht ein Geschlechtsdimorphismus mit durchschnittlich größeren männlichen gegenüber weiblichen Individuen. Im äußerlichen Erscheinungsbild gleicht der Kalifornische Maulwurf seinen Verwandten. Wie diese ist er an eine grabende Lebensweise angepasst. Dadurch zeigt sich sein Körper walzenförmig, der Hals ist kurz und die vorderen Gliedmaßen sind zu Grabwerkzeugen mit breiten, kräftigen Nägeln umgestaltet. Die Schnauze ist konisch zugespitzt und unbehaart sowie rosafarben. Sie zeichnet sich durch sichelförmige Nasenlöcher aus. Die Augen sind winzig und werden vom Fell nahezu vollständig verdeckt, äußere Ohrmuschel sind nicht erkennbar. Das kurze, seidige Fell ist schwarzbraun bis silbergrau gefärbt und weist zuweilen eine messing- oder kupferfarbene Rückenbehaarung auf. Im Winter dunkelt das Fell ab, Tiere aus dem trockenen Süden sind generell heller. Bei einigen Exemplaren kommen creme- bis orangefarbene oder weißliche Flecken am Kinn und im Gesicht vor beziehungsweise sie sind generell zimt- bis fahlgelb gefärbt. Insgesamt treten aber Individuen mit Farbanomalien eher selten auf. Den Schwanz bedeckt ein dichter silberfarbener Haarflaum. Der Hinterfuß misst 16 bis 2,5 cm in der Länge. Weibchen haben vier Paare an Zitzen, zwei in der Brust- und jeweils eins in der Bauch- und Lendengegend.[1][2]
Schädel- und Gebissmerkmale
Der Schädel wird 31,8 bis 37,4 mm lang und im Bereich der Warzenfortsätze 15,9 bis 17,7 mm breit. Die Weite auf Höhe der Augeneinschnürung beträgt 7,3 bis 8,6 mm. Der Abschnitt im Bereich der Orbita ist rechteckig gestaltet. Insgesamt ist der Schädel schlanker als im Verhältnis bei Scapanus occultus, so dass die Mastoidbreit weniger als 49 % der Gesamtlänge ausmacht.[3] Die Stirnlinie verläuft relativ gerade und flach. Im Vergleich zum Townsend-Maulwurf (Scapanus townsendii) und zum Pazifischen Maulwurf (Scapanus orarius) wirkt das Rostrum kürzer und breiter. Der Mittelkieferknochen überragt nach vorn das Nasenbein. Die Nasenöffnungen sind nach vorn gerichtet. Der Gaumen ist deutlich gestreckt. Der moderat massige Unterkiefer weist einen langgestreckten Kronenfortsatz auf. Wie bei den meisten anderen Westamerikanischen Maulwürfen besteht das Gebiss aus 44 Zähnen, die Zahnformel lautet: . Abweichend zu den weiter südlich verbreiteten Westamerikanischen Maulwürfen wie Scapanus occultus und der Baja-California-Maulwurf (Scapanus anthonyi) kommt es beim Kalifornischen Maulwurf nicht zu einer Reduktion der Zahnanzahl. Entsprechend den übrigen Neuweltmaulwürfen ist der erste obere Schneidezahn stark vergrößert, zusätzlich kommt beim Kalifornischen Maulwurf ein großer, eckzahnartiger unterer zweiter Schneidezahn vor, der die beiden anderen überragt. Die Prämolar stehen anders als beim Townsend-Maulwurf dicht gedrängt. Sie verfügen alle über jeweils eine Wurzel. Die vorderen Zähne einschließlich der ersten beiden Prämolaren der oberen und unteren Zahnreihe sind einspitzig (unicuspid), variieren in ihrer Größe aber stärker. Die beiden hinteren Prämolaren und die ersten beiden Molaren sind gegenüber den vorderen Zähnen außerordentlich groß. Die oberen Zahnreihe wird 10,2 bis 11,6 mm lang, die untere Prämolaren-Molarenreihe hingegen 9,6 bis 11,3 mm.[4][1][2]

Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 34. Es besteht aus insgesamt 16 zweiarmigen Autosomenpaaren, die Form der Geschlechtschromosomen ist unbekannt. Die Anzahl der Arme der Autosomen (fundamentale Anzahl) wird mit 60 angegeben.[5][6][2]
Verbreitung und Lebensraum
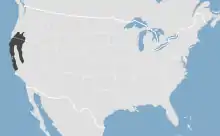
Das Verbreitungsgebiet des Kalifornischen Maulwurfs erstreckt sich mit mehreren Unterarten entlang der Pazifikküste vom südlichen Oregon durch weite Gebiete Kaliforniens bis in den Nordwesten von Baja California. Die Nordgrenze wird etwa beim Crater Lake im Klamath County und dem Fort Rock Basin im Lake County erreicht,[7] die Südgrenze liegt etwa auf Höhe von Santa Barbara und dem Yosemite Valley. Das östlichste Vorkommen erreicht den Westrand Nevadas. Eine isolierte Population besteht auf Angel Island in der Bucht von San Francisco. Ausgespart vom Vorkommen der Art bleiben Teile des San Joaquin Valleys. Der Kalifornische Maulwurf tritt bevorzugt in Gebieten mit feuchten und reichhaltigen Böden auf, die mit Wiesen, Weiden sowie Wäldern mit geringer Bodenvegetation bestanden sind. Die Höhenverbreitung reicht vom Meeresspiegelniveau bis in höhere Gebirgslagen. Hochgelegene Nachweise fanden sich mit rund 2900 m im Yosemite-Nationalpark in der Sierra Nevada sowie mit rund 2600 m in der Kaskadenkette. Im nördlichen Teil schließt der Lebensraum offene Buschgebiete mit Artemisia-Pflanzengemeinschaften ein, im Süden auch wüstenartige Landschaften. Vor allem hier suchen die Tiere bevorzugt feuchtere Bereiche etwa unter Felsblöcken auf. Generell sind felsige Regionen sowie steinige oder tonige Böden jedoch ein Hindernis für die Verbreitung der Art. Auf sehr sandigen Böden fehlt sie ebenfalls, da dort entweder nicht genügend Nahrung zur Verfügung steht oder der Boden für den Bau von unterirdischen Tunneln ungeeignet ist. Ebenso können die Tiere in Gebieten, in denen der Boden für lange Zeit gefroren bleibt, nicht überleben.[4][1][2]
Lebensweise
Territorialverhalten

Die Lebensweise des Kalifornischen Maulwurfs ist vergleichsweise ungenügend erforscht. Die Tiere leben unterirdisch und graben mit ihren kräftigen Vorderfüßen Tunnelsysteme. Diese verlaufen in mehreren Ebenen. Oberflächennahe Gänge liegen häufig nur 2 cm tief und sind oberirdisch als Rippeln erkennbar, die zahlreiche Abzweigungen aufweisen. Tiefere Tunnel verlaufen rund 25 bis 45 cm unter der Erdoberfläche. Hier befinden sich auch die Nester, die mit Pflanzen ausgepolstert werden und zusätzlich zum Schutz unter Wurzeln oder Büschen liegen. Die beim Graben anfallende überschüssige Erde wird in charakteristischen Auswurfhügeln (Maulwurfshügel) an die Oberfläche gedrückt. Da das Aushubmaterial von unten nach oben geschoben wird, sind die Tiere selten an der Oberfläche des Hügels sichtbar. Jedes Individuum beansprucht ein eigenes Gangsystem. Aufgelassene Gänge können innerhalb von eins bis zwei Tagen wieder besiedelt werden. An der zentralsüdlichen Küste von Kalifornien wurden drei Gangsysteme in einem Abstand von 1,0 bis 1,5 km zueinander dokumentiert.[8][1][2]
Der Kalifornische Maulwurf ist ein guter Schwimmer und bewegt sich dabei mit alternierenden Bewegungen der Vorder- und Hinterbeine fort. Ein Tier schwamm bis zu sechs Minuten lang ohne zu ermüden, allerdings war nach rund einer Minute das Fell so stark durchnässt, dass es den Körper nicht mehr waagerecht halten konnte. Auf festem Boden erreicht der Kalifornische Maulwurf eine Geschwindigkeit bis zu bis zu 0,6 m je Sekunde auf einer überwundenen Distanz von rund 12 m.[9][1][2]
Ernährung
Über die Ernährunsgweise wildlebender Individuen liegen aufgrund der versteckten Lebensweise unter der Erdoberfläche nur Einzelergebnisse vor. Der Mageninhalt eines einzelnen Tieres aus Oregon bestand zu 80 % aus Weichtieren und zu 20 % aus unbestimmten Käfern.[10] In Küstennähe werden wohl Sandflöhe wie Megalorchestia vertilgt.[8] Sehr sporadisch vertilgt der Kalifornische Maulwurf seine Nahrung an der Erdoberfläche, ein beobachtetes Individuum fraß dabei einen Spanner.[11] In Gefangenschaft gehaltene Tiere verzehrten sowohl Regenwürmer als auch Fleisch. Die täglich konsumierte Menge betrug zwischen 53,4 und 107,7 % des eigenen Körpergewichts. Sie war bei ausschließlicher Regenwurmnahrung größer als bei reiner Fleischnahrung. Außerdem tranken sie regelmäßig Wasser.[9][12][7][1][2]
Fortpflanzung
Die Fortpflanzung findet vermutlich einmal jährlich statt. Paarungsbereite Männchen wurden im frühen Januar beobachtet, ein nahezu ausgewachsenes Jungtier im Mai. Die Wurfgröße liegt bei zwei bis fünf, durchschnittlich vier Jungen. Sie verbleiben in einem mit Pflanzen gepolsterten unterirdischen Nest. Im Alter von zehn Tagen sind sie rund 7,2 bis 7,5 cm lang. Nach 30 bis 35 Tagen verlassen die Jungen das Nest.[13][1][2]
Fressfeinde und Parasiten
Da der Kalifornische Maulwurf nahezu ausschließlich unter der Erdoberfläche lebt, ist die Anzahl der Fressfeinde begrenzt. Gelegentlich werden Individuen jedoch von Fichtenmardern, Schleiereulen, Kornweihen oder Rotschwanzbussarden erbeutet. Nach Untersuchungen im nordöstlichen Kalifornien beträgt beim Fichtenmarder der durchschnittliche Volumenanteil der Maulwurfsart am gesamten Beutespektrum 3,6 % und der Gewichtsanteil 3,5 %. Im Herbst liegt der jeweilige Anteil mit 7,2 % beziehungsweise 6,9 % gut doppelt so hoch wie im Frühjahr mit 3,2 % beziehungsweise 3,5 %. Durchschnittlich wurde der Kalifornische Maulwurf in rund 2,6 % aller untersuchten Kotreste nachgewiesen.[14] Beobachtungen über die Jagd des Rotschwanzbussards auf die Tiere beschränken sich auf die erste Jahreshälfte, was eventuell mit dem Fortpflanzungsverhalten des Kalifornischen Maulwurfs zusammenhängt.[15][1]
Bisher wurden zumeist äußere Parasiten dokumentiert. Belegt sind unter anderm Flöhe der Gattung Corpsylla, des Weiteren Zecken der Gattung Ixodes und Milben der Gattungen Androlaelaps und Haemogamasus.[16][1]
Systematik
Innere Systematik der Westamerikanischen Maulwürfe nach Álvarez-Castañeda und Cortes-Calva 2021[3]
|
Der Kalifornische Maulwurf ist eine Art innerhalb der Gattung der Westamerikanischen Maulwürfe (Scapanus), welche insgesamt fünf Vertreter vereint. Art und Gattung wiederum gehören zur Familie der Maulwürfe (Talpidae), innerhalb derer sie gemeinsam mit einigen weiteren Gruppen aus Nordamerika und Asien die Tribus der Neuweltmaulwürfe (Scalopini) bilden. Die Neuweltmaulwürfe repräsentieren vergleichbar den Eigentlichen Maulwürfen (Talpini) grabende Angehörige der Familie, andere Vertreter leben nur teils unterirdisch, bewegen sich auf der Erdoberfläche fort oder verfolgen eine semi-aquatische Lebensweise. Molekulargenetische Untersuchungen datieren die Abspaltung der Neuweltmaulwürfe von den anderen Triben der Maulwürfe in das Obere Eozän vor rund 39 bis 35 Millionen Jahren.[17] Die Tribus kann in zwei Entwicklungslinien aufgliedert werden: die Parascalopina und die Scalopina. Beide unterscheiden sich vor allem an der Ausprägung des Metastylids am unteren zweiten Molar.[18][19] Den Scalopina fehlt das Metastylid, bei den Parascalopina hingegen kommt es vor. Beide Linien entwickelten sich bereits seit dem Unteren Miozän vor 21,4 Millionen Jahren getrennt voneinander. Die Westamerikanischen Maulwürfe gehören den Scalopina an und stehen somit in einer engeren Beziehung zum Ostamerikanischen Maulwurf (Scalopus). Die Diversifizierung der Scalopina setzte im Mittlere Miozän vor gut 14 Millionen Jahren ein, eine deutlichere Auffächerung der Westamerikanischen Maulwürfe ist seit dem Übergang vom Miozän zum Pliozän vor gut 6 Millionen Jahren zu verzeichnen.[20][21]

Die wissenschaftliche Erstbeschreibung des Kalifornischen Maulwurfs erstellte John Bachman im Jahr 1841 im Rahmen einer Abhandlung über die bis dahin bekannten nordamerikanischen Maulwürfe. Teilweise wird in der Literatur auch 1842 als Erstbeschreibungsjahr angegeben, da Bachman kurz darauf einen weiteren, vergleichbaren Aufsatz veröffentlichte. Als wissenschaftliche Bezeichnung nutzte er jeweils Scalops latimanus, womit er die neue Art in die Nähe des Ostamerikanischen Maulwurfs verwies. Bachman standen zwei Exemplare aus dem Naturkundemuseum Berlin zur Verfügung, die beide aus der Sammlung von Wilhelm Deppe stammten. Als besonderes Merkmal hob er die breite Handfläche hervor, worauf sich auch das Artepitheton bezieht (von lateinisch latus für „breit“ und manus für „Hand“). Ansonsten beschrieb er den Kalifornischen Maulwurf als intermediär zwischen dem Townsend-Maulwurf (Scapanus townsendii) und dem Ostamerikanischen Maulwurf. Die Herkunft der Belegexemplare gab Bachman nur allgemein mit „Mexiko“ an.[22][23] Gut 20 Jahre später konnte Wilhelm Peters eines der Originalexemplare ausfindig machen. In einem Aufsatz aus dem Jahr 1863 wies er darauf hin, dass die Eingangsbücher der Sammlung Deppes den Fundort mit „Santa Clara“ wesentlich genauer auswiesen, als Bachman es vermerkt hatte. Der Zeitpunkt der Auffindung war mit Oktober 1834 angegeben. Mit „Santa Clara“ vermutete Peters eine Lage in Sonora im nördlichen Mexiko. Abgesendet wurde das Exemplar laut den Eingangsbüchern von Monterey in Kalifornien.[24] Erst im Jahr 1907 klärte Wilfred Hudson Osgood auf, dass mit „Santa Clara“ keine mexikanische Lokalität, sondern die gleichnamige Stadt in Kalifornien gemeint war, welche nördlich von Monterey liegt. Ähnliches hatte bereits Frederick W. True in seiner Revision der nordamerikanischen Maulwürfe aus dem Jahr 1896 vermutet.[25] Santa Clara in Kalifornien gilt heute als Typusregion der Art.[26][27]
In seinem Aufsatz von 1863 erkannte Peters eine im Vergleich zum Ostamerikanischen Maulwurf höhere Anzahl an Zähnen. Er verwies Bachmans Belegexemplar daher zur Gattung Scapanus, die wiederum im Jahr 1848 von Auguste Pomel eingeführt worden war.[28] Gleichzeitig betrachtete Peters den Kalifornischen Maulwurf als synonym zum Townsend-Maulwurf.[24] Dies bestätigte zusätzlich True 1896. Allerdings geschah das vorläufig und unter Verweis auf eine angenommene vergleichsweise nördliche Herkunft der Belegexemplare aus Santa Clara in Kalifornien und der damit relativen Nähe zum Verbreitungsgebiet des Townsend-Maulwurfs. Als unterstützendes Argument hob True die auffallend dunklere Färbung gegenüber Maulwürfen des südlichen Kaliforniens hervor. Für diese wiederum nutzte er die Bezeichnung Scapanus californicus.[25] Die Form geht als Scalops californicus auf William Orville Ayres aus dem Jahr 1854 zurück, für die er außer einer allgemeinen Verbreitung an der Westküste keine genaue Typusfundstelle angab.[29] Osgood wiederum hob im Jahr 1907 die Synonymität des Kalifornischen Maulwurfs mit dem Townsend-Maulwurf auf und erkannte ersteren als eigenständig an. Dabei berief er sich auf die deutlich geringere Größe des Kalifornischen Maulwurfs im Vergleich zum Townsend-Maulwurf. Im selben Zug sah er Scapanus californicus als identisch mit dem Kalifornischen Maulwurf an, da sich keine abweichenden Merkmale aufzeigen ließen.[26][27]
| Jackson 1915[30] | Palmer 1937[4] Verts und Carraway 2001[1] Wilson und Reeder 2005[31] | Woodman 2018[27] Kryštufek und Motokawa 2018[2] | Álvarez-Castañeda und Cortes-Calva 2021[3] |
|---|---|---|---|
| n. def. | S. l. caurinus | S. l. latimanus | S. latimanus ssp. |
| S. l. dilatus | S. l. dilatus | ||
| S. l. alpinus | |||
| S. l. latimanus | S. l. latimanus | ||
| n. def. | S. l. insularis | S. l. insularis | |
| n. def. | S. l. parvus | S. l. parvus | |
| S. l. minusculus | S. l. minusculus | S. l. minusculus | |
| n. def. | S. l. campi | ||
| S. l. sericatus | S. l. sericatus | ||
| n. def. | S. l. monoensis | ||
| S. l. grinelli | S. l. grinelli | S. l. occultus | eigene Art |
| S. l. occultus | S. l. occultus | ||
| eigene Art | S. l. anthonyi | eigene Art | eigene Art |
Es werden mehrere Unterarten des Kalifornischen Maulwurfs unterschieden, ihre genaue Anzahl variierte aber im Laufe der Forschungsgeschichte. Während Hartley H. T. Jackson in seiner Revision der nordamerikanischen Maulwürfe aus dem Jahr 1915 noch insgesamt sieben Unterarten herausstellte,[30] führte Fletcher G. Palmer in seiner Abhandlung zum Kalifornischen Maulwurf aus dem Jahr 1937 bereits insgesamt ein Dutzend auf, von denen er aber drei selbst benannt hatte.[4] Die gleiche Anzahl gaben Robert Joe Verts und Leslie N. Carraway im Jahr 2001[1] gleichfalls wie Don E. Wilson und DeeAnn M. Reeder im Jahr 2005 an.[31] Allerdings schlossen sowohl Palmer als auch Verts und Carraway beziehungsweise Wilson und Reeder den Baja-California-Maulwurf (Scapanus anthonyi) als Unterart ein. In einer Überprüfung der nordamerikanischen Insektenfresser aus dem Jahr 2018 wies Neal Woodman insgesamt fünf Unterarten aus.[27] Dies wurde im gleichen Jahr durch Boris Kryštufek und Masaharu Motokawa für den achten Band des Standardwerkes Handbook of the Mammals of the World übernommen.[2] In beiden Systematiken war hier der Baja-California-Maulwurf als eigenständige Art gelistet. Im Jahr 2021 wurde aufgrund genetischer Befunde Scapanus occultus als südlichste Unterart ebenfalls auf Artstatus verschoben. So beträgt etwa der genetische Abstand zwischen diesem und dem Kalifornischen Maulwurf 2,5 bis 5,8 %, zum Baja-California-Maulwurf sind 7,2 bis 10,5 %.[3]
Dadurch können heute folgende Unterarten als anerkannt angesehen werden:[27][2]
- S. l. insularis F. G. Palmer, 1937; Angel Island
- S. l. latimanus (Bachman, 1842); Nominatform; nördliches und westzentrales Kalifornien, südliches Oregon und nordwestliches Nevada
- S. l. minusculus Bangs, 1899; zentralöstliches Kalifornien und zentralwestliches Nevada
- S. l. parvus F. G. Palmer, 1937; zentralwestliches Kalifornien
Fossilfunde des Kalifornischen Maulwurfs sind vergleichsweise häufig, aber auf Einzelfunde beschränkt. Sie verteilen sich hauptsächlich über Kalifornien. Ein isolierter Mahlzahn und ein Oberarmknochen von der Typusfundstelle des Irvingtoniums, einer Stufe in der nordamerikanischen Biostratigraphie, im Alameda County datieren in die jüngere Phase des Altpleistozäns vor 1,5 bis 0,7 Millionen Jahren und zählen zu den ältesten bekannten Fossilresten der. Die meisten anderen Nachweise sind aus dem Jungpleistozän belegt. Noch in dessen Beginn gehören ein ebenfalls isolierter Mahlzahn und einzelne Elemente der Vordergliedmaßen aus dem Sacramento County mit einem Alter von 103.000 Jahren. Erwähnenswert sind auch Reste aus verschiedenen Asphaltgruben von Rancho La Brea in Los Angeles, die im ausgehenden Oberpleistozän entstanden. Sie umfassen wiederum Zähne sowie Gliedmaßenknochen. Bereits den Übergang zum Holozän markieren mehrere Schlüsselbeinfunde aus dem San Bernardino County. Von Bedeutung sind ein Teilschädel und Teile des Körperskeletts von der Fundstelle Glen Abbey im San Diego County. Der recht reichhaltige küstennahe Fossilplatz, der unter anderem auch die Wandertaube und zahlreiche Kleinsäuger erbrachte, wurde im Jahr 1980 entdeckt und besitzt ein spät-jungpleistozänes Alter. Der Schädelfund des Maulwurfs wurde ursprünglich aufgrund des Fehlens des ersten Prämolaren dem Baja-California-Maulwurf zugeschrieben, der heute aber weiter südlich auftritt. Der Verlust dieses Zahns ist allerdings auch von den südlichen Populationen des Kalifornischen Maulwurfs dokumentiert, weswegen andere Autoren dazu tendieren, die Funde dieser Art zuzuschreiben.[32][33] Ein naher Verwandter des heutigen Kalifornischen Maulwurfs ist möglicherweise das ausgestorbene Scapanus malatinus, nachgewiesen über Schädelreste, isolierte Zähne und postcraniale Skelettteile. Das Typusmaterial lagerte in der Palm-Springs-Formation ebenfalls im San Diego County und lässt sich dem unteren Abschnitt des Altpleistozäns vor rund 1,9 bis 1,7 Millionen Jahren zuweisen.[32][34]
Bedrohung und Schutz
Die IUCN listet den Kalifornischen Maulwurf in der Kategorie „nicht gefährdet“ (least concern). Begründet wird dies mit der weiten Verbreitung, dem vermuteten häufigen Aufkommen und einer gewissen Toleranz gegenüber Landschaftsveränderungen. Außerdem treten die Tiere in verschiedenen Schutzgebieten auf. Größere Bestandgefährdungen sind nicht bekannt. In einzelnen Gebieten werden die Tiere als Schädlinge angesehen.[35]
Literatur
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 600) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage, Johns Hopkins University Press, Baltimore/London, 1999, ISBN 0-8018-5789-9
- B. J. Verts und Leslie N. Carraway: Land mammals of Oregon. University of California Press, Berkeley, 1998, S. 68–69
- B. J. Verts und Leslie N. Carraway: Scapanus latimanus. Mammalian Species 666, 2001, S. 1–7
Einzelnachweise
- B. J. Verts und Leslie N. Carraway: Scapanus latimanus. Mammalian Species 666, 2001, S. 1–7
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 600) ISBN 978-84-16728-08-4
- Sergio Ticul Álvarez-Castañeda und Patricia Cortes-Calva: Revision of moles in the genus Scapanus. Therya 12 (2), 2021, S. 275–281, doi:10.12933/therya-21-1174
- Fletcher G. Palmer: Geographic variations in the mole Scapanus latimanus. Journal of Mammalogy 18 (3), 1937, S. 280–314
- J. W. F. Reumer und A. Meylan: New developments in vertebrate cytotaxonomy. IX Chromosome numbers in the order Insectivora (Mammalia). Genetica 70, 1986, S. 119–151
- Shin-ichiro Kawada, Masashi Harada, Kazuhiru Koyasu und Sen-ichi Oda: Karyological note on the short-faced mole, Scaptochirus moschatus (Insectivore, Talpidae). Mammal Study 27, 2002, S. 91–94, ()
- B. J. Verts und Leslie N. Carraway: Land mammals of Oregon. University of California Press, Berkeley, 1998, S. 68–69
- Howard McCully: The Broad-handed mole, Scapanus latimanus, in an marine littoral environment. Journal of Mammalogy 48 (3), 1967, S. 480–482
- Charles A. Reed und Thane Riney: Swimming, feeding and locomotion of a captive mole. The American Midland Naturalist 30 (3), 1943, S. 790–791
- John O. Whitaker, Jr., Chris Maser und Richard J. Pedersen: Food and Ectoparasitic Mites of Oregon Moles. Northwest Science 53 (4), 1979, S. 268–273
- Vladimir Dinets: Surface foraging in Scapanus moles. Mammalia 82 (1), 2018, S. 48–53, doi:10.1515/mammalia-2016-0091
- John Norman Grim: Feeding habits of the Southern California mole. Journal of Mammalogy 39 (2), 1958, S. 265–268
- Alan C. Ziegler: Milk dentition in the Broad-footed mole, Scapanus latimanus. Journal of Mammalogy 53, 1972, S. 354–355
- William J. Zielinsky, Wayne D. Spencer und Reginald H. Barrett: Relationship between food habits and activity patterns of the Pine martens. Journal of Mammalogy 64 (3), 1983, S. 387–396
- John C. Hafner: Seasonal Predation on Moles by the Red-Tailed Hawk. The Condor 76 (2), 1974, S. 225
- Terry L. Yates, Danny B. Penc und Glenn K. Launchbaugh: Ectoparasites from seven species of North American moles (Insectivora: Talpidae). Journal of Medical Entomology 16, 1979, S. 166–168
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang, Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- J. Howard Hutchinson: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117 (S. 58–96)
- Achim Schwermann, Kai He, Benjamin J. Peters, Thorsten Plogschties und Gabrielle Sansalone: Systematics and macroevolution of extant and fossil scalopine moles (Mammalia, Talpidae). Palaeontology 62 (4), 2019, S. 661–676, doi:10.1111/pala.12422
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Zhong-Zheng Chen, Shui-Wang He, Wen-Hao Hu, Wen-Yu Song, Kenneth O. Onditi, Xue-You Li und Xue-Long Jiang: Morphology and phylogeny of scalopine moles (Eulipotyphla: Talpidae: Scalopini) from the eastern Himalayas, with descriptions of a new genus and species. Zoological Journal of the Linnean Society, 2021, S. zlaa172, doi:10.1093/zoolinnean/zlaa172
- John Bachman: Observations on the genus Scalops, with descriptions of the species found in North America. Proceedings of the Boston Society of Natural History 1, 1841, S. 40–41 ()
- John Bachman: Observations on the genus Scalops, (shrew moles,) with descriptions of the species found in North America. Boston Journal of Natural History 4, 1843–1844, S. 26–35 ()
- Wilhelm Peters: Über neue Eichhornarten aus Mexico, Costa Rica und Guiana, so wie über Scalops latimanus Bachmann. Monatsberichte der Koniglichen Preussischen Akademie der Wissenschaften zu Berlin 1863, S. 652–656 ()
- Frederick W. True: A revision of the American moles. Proceedings of the United States National Museum 19 (1101), 1896, S. 1–112 (S. 52)
- Wilfred H. Osgood: Some unrecognized and misapplied names ofAmerican mammals. Proceedings of the Biological Society of Washington 20, 1907, S. 43–52 ()
- Neal Woodman: American Recent Eulipotyphla. Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithsonian Institution Scholary Press, 2018, S. 1–107 (S. 25–27)
- Auguste Pomel: Etudes sur les carnassiers insectivores (Extrait). Archives des sciences physiques et naturelles, Geneve 1 (9), 1848, S. 244–251 ()
- William Orville Ayres: The ground mole. Proceedings of the California Academy of Natural Sciences 1, 1854, S. 54 ()
- Hartley H. T. Jackson: A review of the American moles. North American Fauna 38, 1915, S. 1–100 (S. 54–75)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- J. Howard Hutchinson: Moles of the Scapanus latimanus group (Talpidae, Insectivora) from the Pliocene and Pleistocene of California. Contributions in Science, Natural History Museum of Los Angeles County 386, 1987, S. 1–15
- C. Paul Majors: Preliminary report on a Late Pleistocene vertebrate assemblage from Bonita, San Diego County, California. PaleoBios 15 (4), 1993, S. 63–77
- J. Howard Hutchinson: Late Pliocene (Blancan) Scapanus (Scapanus) (Talpida: Mammalia) from the Glenns Ferry Formation of Idaho. PaleoBios 12 (45), 1987, S. 1–7
- J. Matson, N. Woodman, I. Castro-Arellano und P. C. de Grammont: Scapanus latimanus (errata version published in 2017). The IUCN Red List of Threatened Species 2016. e.T41473A115188559 (); zuletzt aufgerufen am 4. April 2021
Weblinks
- Scapanus latimanus in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: J. Matson, N. Woodman, I. Castro-Arellano & P. C. de Grammont, 2016. Abgerufen am 4. April 2021.