Vegetation
Vegetation (mittellateinisch vegetātio „Wachstumskraft“, „Grünung“ und spätlateinisch vegetāre „leben, wachsen“) – im Deutschen auch Pflanzendecke oder Pflanzenkleid – nennt man im Allgemeinen die Gesamtheit der Pflanzenformationen einer Landfläche. Im Gegensatz zur Flora, bei der vorrangig die Pflanzensippen und -arten betrachtet werden, bezieht sich der Begriff Vegetation auf gleichartige Verteilungsmuster, Gestalt- und Wuchsformen (Offenland, Wald, Tundra, Steppe, Tropischer Regenwald u. dgl.). In Geobotanik, Biogeographie und Ökologie werden neben den Pflanzen alle Lebensformen zur Vegetation gerechnet, die sich nicht aktiv fortbewegen können, also auch Flechten und Pilze.[1] In der Medizin versteht man unter Vegetation die Besiedlung mit Bakterien beziehungsweise Biofilm, zum Beispiel auf Herzklappen bei infektiöser Endokarditis. Das Verb vegetieren bedeutet „untätig“ zu leben wie eine Pflanze.[2]

.jpg.webp)

Sofern die Bedingungen für Leben (Luft, Licht, Wasser, Nährstoffe) – zumindest zeitweise – gegeben sind, stellt sich auf jeder Oberfläche im Laufe der Zeit eine standorttypische Vegetation ein. In extrem kalten oder trockenen Regionen kann sie aus sehr wenigen Arten bestehen – etwa ausschließlich Flechten. Die Zeit, in der der Bewuchs wachsen kann, heißt Vegetationsperiode. Je wärmer und feuchter es ist und je länger diese Bedingungen anhalten, desto komplexer ist das entstehende Ökosystem. Bis zu seiner optimalen Ausgestaltung – ohne wiederkehrende Störungen Klimaxvegetation genannt – durchläuft der Bewuchs eine erprobte Abfolge von Stadien mit ganz unterschiedlicher Artenzusammensetzung. Diese Entwicklung nennt man Sukzession. Im Laufe sehr viel längerer Zeiträume führt die zwangsläufige Entstehung von Böden – durch die fortlaufende Umwandlung anorganischer Nährstoffe in organische Verbindungen sowie die zunehmende Ablagerung toter Biomasse – zu einem langsamen Wandel der Vegetation. Wenn sich währenddessen die klimatischen Bedingungen ändern und sich in der permanenten Evolution neu entstandene Lebensformen oder vom Menschen eingebrachte Neobiota als Bestandteil des Ökosystems langfristig bewähren und anpassen, können völlig neue Vegetationstypen mit zum Teil gänzlich neuen Eigenschaften entstehen, die vorher nicht existierten. Im Idealfall beeinflusst und stabilisiert die Vegetation selbst die Umweltbedingungen in einer für sie zuträglichen Weise (etwa die Aufrechterhaltung des feuchtheißen Klimas im tropischen Regenwald durch den Wald).
Die Pflanzenwelt stellt durch ihre Photosyntheseleistung (Erzeugung von Zucker aus Wasser und Kohlenstoffdioxid unter Verwendung von Lichtenergie) die mit Abstand größte Menge von Primärproduzenten für die Biomasse an Land und bildet damit die entscheidende Lebensgrundlage für nahezu jedes Ökosystem.
Wachstum nach dem „Optimalprinzip“
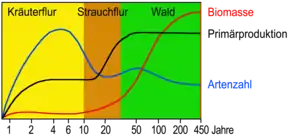

Obwohl die Vegetation weit voneinander entfernter und nicht miteinander verbundener Regionen zu einem großen Teil aus nicht direkt verwandten Arten besteht, strebt die Natur immer nach einer möglichst großen Umwandlung der vorhandenen abiotischen Faktoren in Biomasse. Häufig entstehen unabhängig voneinander die gleichen Überlebensstrategien (analoge Entwicklungen wie beispielsweise die Wasserspeicherfähigkeit von Sukkulenten oder der Laubfall als Kälteschutz), die sich aufgrund gemeinsamer Strategien gegen vorherrschende extreme Umweltbedingungen (Trockenheit oder Nässe, Hitze oder Kälte, regelmäßige Klimaschwankungen im Tages- oder Jahresgang, Feuer, Fressfeinde usw.) evolutionär entwickelt haben.
Die natürliche Sukzession des Artenwandels der Pflanzendecke strebt im voll entwickelten Schlussstadium immer und überall zu einem optimalen Zustand der Vegetation – nach Gustav Wendelberger (1978) zur „Optimalgesellschaft“.[3] Seine Gesellschaft höchster Lebensfülle – die in der Waldökologie auch „Optimalphase“[4] genannt wird – kann nach heutigen Erkenntnissen als Vegetationstyp verstanden werden, der je nach den vorherrschenden Umweltfaktoren (insbesondere Klima u. Bodenbeschaffenheit) in einer Region folgende Zustände optimal verbindet:[5][6][7]
- ein ideal angepasstes Artenspektrum,
- Fließgleichgewicht zwischen konkurrierenden Arten
- und Besatz möglichst jeder ökologischen Nische,
- sowie eine unter diesen Bedingungen maximale Menge an Biomasse.[8]
Auch Artenvielfalt und Biodiversität befinden sich auf einem hohen Niveau: Allerdings besteht die maximaler Vielfalt nicht in der Schlussgesellschaft, sondern während der gesamten Sukzessionsfolge in unterschiedlicher Zusammensetzung. So zeigte etwa eine 20-jährige Studie aus dem Raum Bremen, dass die Pflanzenvielfalt zu Beginn der natürlichen Besiedlung einer Deponie am höchsten war, während die Vielfalt der Tierarten viel geringer war als zum Ende des Zeitraumes. So führte die große Zahl potenzieller Pionierpflanzen zu einer schnellen und sicheren Besiedlung aller Nischen des Gebietes, ohne dass zu viele Pflanzenfresser diesen Prozess verlangsamten. Die größte Biodiversität entsteht bei einem möglichst großen Nebeneinander verschiedenster Sukzessionsstadien und nicht innerhalb einer voll entwickelten Schlussgesellschaft.[9]
Diese Betrachtung erklärt auch die höhere Artenvielfalt bäuerlicher Kulturlandschaften im Gegensatz zum geschlossenen, natürlichen Laubwald. Die Kriterien der vorgenannten „Optimalgesellschaft“ erreichen von Menschen geprägte Ersatzgesellschaften nicht nachhaltig. So ist etwa die photosynthetische Effizienz (als Vorbedingung für die Primärproduktion) bei einem natürlichen Wald wesentlich höher als bei einem landwirtschaftlichen Acker (im gleichen Klima). Die Menge der Biomasse aus lebenden und toten Pflanzen und Tieren (Boden) lag bei einem Vergleich im Buchenwald bei 211 t C/ha und im Maisfeld bei 62,5 t C/ha.[10] Sofern intensive Düngung oder Monokulturen zu einer höhere Biomasseproduktion führen, wäre dies unter natürlichen Bedingungen nur vorübergehend der Fall, da die anderen Kriterien (optimales Zusammenspiel der Arten) keine langfristige Existenz garantieren.

Die Anpassungsstrategien der Natur, um dauerhaft eine möglichst hohe Primärproduktion zu erreichen, sind evolutionär in sehr langen Zeiträumen entstanden: Wir sehen sie heute etwa im Blattflächenindex (optimale Ausnutzung des Sonnenlichts durch die Anordnung und Fläche des Laubes), in der „Wuchshöhenregel“ nach Reiner Schwarz (je feuchter, wintermilder, sommerwärmer, strahlungsreicher das Klima bei eher nährstoffarmen Böden, desto höher reicht das Kronendach eines Waldes)[8] oder der an den Einfallswinkel der Sonnenstrahlen ausgerichteten Wuchsform von Bäumen (zum Beispiel schlanke Nadelbäume mit offenem Kronendach im tiefen Sonnenstand der borealen Zone – sodass die Strahlen möglichst viele Nadeln erreichen – gegenüber deutlich breiteren Kronen in Breitengraden mit höheren Sonnenständen). Die besten Bedingungen für einen üppigen Pflanzenwuchs sind ein immerfeuchtes, kühles, (fast) frostfreies Klima mit sehr großen Niederschlagsmengen und hoher Luftfeuchtigkeit, gedämpfte Sonneneinstrahlung und staunasse Böden: Im Redwood-Nationalpark an der kalifornischen Pazifikküste sind diese Bedingungen offenbar ideal, denn nirgends sonst auf der Erde erzeugt die Vegetation so viel Biomasse pro Flächeneinheit und ist der Blattflächenindex so hoch wie bei den dortigen, über 80 m hohen Küstenmammutbaumwäldern.[11]
Trotz vielfältiger Strategien zur optimalen Energienutzung und Produktion ist auch die Optimalgesellschaft nur ein Modell: Sobald beispielsweise jährlich regelmäßige Buschbrände auftreten (typisch etwa für Savannen) oder riesige Herden großer Pflanzenfresser (etwa die früheren Bisonherden der Prärien Nordamerikas) die Übergangsbereiche zu Waldgebieten baumfrei halten, kann die Sukzession nicht bis zum optimalen Klimax führen.
Vegetationskonzepte
Es wird zwischen verschiedenen Vegetationskonzepten unterschieden:
Ursprüngliche natürliche Vegetation
Die ursprüngliche natürliche Vegetation ist die – v. a. mittels Gesteinsanalysen, Klimamodellen sowie Pollenanalysen und Analysen anderer organischer Funde – rekonstruierte Vegetation, die vor dem Erscheinen von menschlicher Kultur in einem Gebiet und einem bestimmten naturgeschichtlichen Zeitraum ausgebildet gewesen sein könnte.[12] Da die ursprüngliche natürliche Vegetation anderen klimatischen Bedingungen unterworfen war und aus einem anderen Artenpool gebildet wurde, unterscheidet sie sich in der Regel mehr oder weniger deutlich nicht nur von der realen heutigen Vegetation, sondern auch von der heutigen potenziell natürlichen Vegetation.
Reale Vegetation
Die reale Vegetation ist die in einem Gebiet tatsächlich vorkommende Vegetation. Diese ist häufig durch den Menschen beeinflusst, zum Beispiel durch Eingriffe land- und forstwirtschaftlicher Art. Falls kulturelle Einflüsse zu erheblichen Veränderungen der Vegetation führten, handelt es sich um Ersatzgesellschaften, welche die natürlichen Pflanzengesellschaften abgelöst haben.[13]
Potenzielle natürliche Vegetation
Der Begriff „potenzielle natürliche Vegetation“ (pnV) bezeichnet den Endzustand der Vegetation, den man im jeweiligen Gebiet erwarten würde, sofern dort fortan keine menschlichen Eingriffe mehr erfolgen.[14] „Dabei sollen etwaige Änderungen abiotischer Faktoren, die im Laufe einer hypothetischen Sukzession eintreten könnten … gedanklich ausgeschlossen sein.“[15] Die pnV ist also die aufgrund der aktuellen natürlichen und ggf. anthropogen veränderten Standortfaktoren erwartete Vegetation, wenn der menschliche Einfluss beendet würde. Die pnV ist daher zumeist nicht mit jener Vegetationsform identisch, die man erwarten würde, wenn der Mensch nie eingegriffen hätte (= Rekonstruierte natürliche Vegetation).[16] Man kann den Begriff der pnV auf verschiedene Zeiten beziehen. Bezieht man ihn auf die heutige Zeit, so spricht man von der heutigen potenziell natürlichen Vegetation (hpnV). Wenn keine Zeitangabe gemacht wird, meint pnV die hpnV.[17]
Der Begriff der potentiellen natürlichen Vegetation baut primär auf dem Konzept einer Schlussgesellschaft von Pflanzen, der sogenannten Klimaxvegetation, auf.[18] Damit wird eine längerfristig stabile Vegetationsform bezeichnet, die sich an einem Wuchsort nach einer Sukzessionsreihe, während der sich die Standorteigenschaften des Wuchsortes verändern können, einstellt. Die Klimaxvegetation selbst unterliegt in sehr langen Zeiträumen einem allmählichen Wandel durch klimatische, geologische und floristische Veränderungen an einem Wuchsort (z. B. Vegetationsentwicklung im Holozän). Ein alternatives Konzept zur Klimaxvegetation ist das Mosaik-Zyklus-Konzept.
Rekonstruierte natürliche Vegetation
Die rekonstruierte natürliche Vegetation ist die rekonstruierte Vegetation, die man in einem Gebiet erwarten würde, wenn der Mensch dort niemals erschienen wäre. Sie berücksichtigt im Gegensatz zur potentiellen Vegetation also keine anthropogenen Veränderungen, sondern schließt diese durch Extrapolierungen aus.[18] Die rekonstruierte natürliche Vegetation wird für den Naturschutz im Prozessschutz und für das Wildniskonzept relevant.
Weblinks
Einzelnachweise
- Jörg S. Pfadenhauer und Frank A. Klötzli: Vegetation der Erde. Springer Spektrum, Berlin/Heidelberg 2014, ISBN 978-3-642-41949-2. S. 2.
- Zeno.org, Vegetieren. Abgerufen am 11. April 2021.
- Gustav Wendelberger: Das vierdimensionale Vegetationsgefüge der Erde, in Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien. Früher: Verh. des Zoologisch-Botanischen Vereins in Wien. seit 2014 "Acta ZooBot Austria", Band 116–117, 1978, pdf. abgerufen am 11. Februar 2022, S. 150–151.
- Richard Pott: Allgemeine Geobotanik. Biogeosysteme und Biodiversität. Springer, Berlin, Heidelberg, New York 2005, ISBN 3-540-23058-0. S. 459.
- Wolfgang Frey, Rainer Lösch: Geobotanik. Pflanze und Vegetation in Raum und Zeit. 3. Auflage. Spektrum Akademischer Verlag, Heidelberg 2010, ISBN 978-3-8274-2335-1, S. 276, 307–309.
- Zitat: „Sowohl bei der primären wie der sekundären Sukzession ändert sich im Laufe der Zeit in den verschiedenen Sukzessionsstadien die Artenzusammensetzung. Die Entwicklung strebt zu einem stabilen Endzustand, der Schlussgesellschaft oder dem Klimaxstadium, entgegen. Dort ändert sich die Artzusammensetzung kaum oder nicht mehr. Höchste Akkumulation an Biomasse.“ aus Albert-Ludwigs-Universität Freiburg: Sukzession Zusammenfassung, Kurs Biologie und Ökologie, Vorlesungsteil Vegetationskunde, 2. Fachsemester Umweltnaturwissenschaften auf studocu.com, online, abgerufen am 25. Februar 2022. S. 6/2.
- Zitat: „Ein aus Biodiversitätssicht optimaler Zustand wird dann annähernd erreicht, wenn der Wald solche Baumarten aufweist, welche der jeweiligen „potenziellen natürlichen Vegetation“ entsprechen und deren genetische Zusammensetzung vom Menschen möglichst unbeeinflusst ist, genügend Totholz und Veteranenbäume aufweist, sich ohne negativen Wildeinfluss und Waldweide natürlich verjüngen kann […]“ in Bundesforschungszentrum für Wald: BFW.Praxisinformation: Biodiversität im Wald, Nr. 37, Wien 2015, ISSN 1815-3895, pdf, abgerufen am 25. Februar 2022. S. 7.
- Reiner Schwarz: Die Wuchshöhenregel der ökologischen Pflanzengeographie, in: Erdkunde, 53, 1999. S. 245–247.
- Hartmut Koehler u. Josef Müller: Entwicklung der Biodiversität während einer 20 jährigen Sukzession als Grundlage für Managementmaßnahmen, Universität Bremen, Zentrum für Umweltforschung und Umwelttechnologie, Bremen 2003, pdf. abgerufen am 11. Februar 2022. S. 98, 180
- Umweltbundesamt: Der "gute ökologische Zustand" naturnaher terrestrischer Ökosysteme - ein Indikator für Biodiversität?, Tagungsband zum Workshop in Dessau 19./20.9.2007, Texte 29/08, Dessau/Roßlau 2008, ISSN 1862-4804, pdf. S. 24.
- Robert Van Pelt, Stephen C. Sillett, William A. Kruse, Russell Kramer: Emergent crowns and light-use complementarity lead to global maximum biomass and leaf area in Sequoia sempervirens forests, in Forest Ecology and Management 375 (2016), DOI: 10.1016/j.foreco.2016.05.018S. S. 279–308
- Otti Wilmanns: Ökologische Pflanzensoziologie. Heidelberg 1993: 56. Vgl. Reinhold Tüxen: Die Grundlagen der Urlandschaftsforschung. Ein Beitrag zur Erforschung der Geschichte der anthropogenen Beeinflussung der Vegetation Mitteleuropas. Niedersächsisches Jahrbuch für Landesgeschichte 8, 1931: S. 59–105.
- Otti Wilmanns: Ökologische Pflanzensoziologie. Heidelberg 1993: 55.
- Reinhold Tüxen: Die heutige potentielle natürliche Vegetation als Gegenstand der Vegetationskartierung. Angew. Pflanzensoz. 13, 1956: S. 5–42
- Otti Wilmanns: Ökologische Pflanzensoziologie. Heidelberg 1993: 55.
- Otti Wilmanns: Ökologische Pflanzensoziologie. Heidelberg 1993: 56.
- Otti Wilmanns: Ökologische Pflanzensoziologie. Heidelberg 1993: 55f.
- Alessandro Chiarucci, Miguel B. Araújo, Guillaume Decocq, Carl Beierkuhnlein & José María Fernández-Palacios: The concept of potential natural vegetation: an epitaph? Journal of Vegetation Science 21: 1172–1178, 2010.