Confuciusornis
Confuciusornis ist eine urtümliche Vogelgattung, deren fossile Reste in der chinesischen Provinz Liaoning in Sedimentgesteinen der unteren Kreidezeit entdeckt wurden. Die Vollständigkeit zahlreicher zu Tage geförderter Skelette und gut erhaltene Gefieder-Abdrücke machen Confuciusornis zu einem der am besten bekannten Vögel des Erdmittelalters.
| Confuciusornis | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Lebendrekonstruktion von Confuciusornis sanctus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Untere Kreidezeit | ||||||||||||
| 125 bis 110 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Confuciusornis | ||||||||||||
| Hou, Zhou, Gu & Zhang, 1995 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Geschichte der Entdeckung und Erforschung

Noch vor den Funden gefiederter Dinosaurier, welche die Volksrepublik China in den Mittelpunkt paläontologischer Forschung rückten, wurden im Jahr 1993 in der Gegend um Sihetun und Jianshangou nahe der Stadt Beipiao die fossilen Überreste eines etwa 30 Zentimeter großen Vogels entdeckt und 1995 von einer chinesischen Forschergruppe um Lianhai Hou als „Confuciusornis sanctus“ („heiliger Konfuzius-Vogel“) beschrieben.[1] Besonders die Fundstelle bei Sihetun erwies sich seither als sehr reich an Vogelfossilien:
Mehr als 1000 Exemplare der Gattung Confuciusornis sind dort bis zur Jahrtausendwende geborgen worden. Confuciusornis sanctus blieb nicht die einzige Art der Gattung Confuciusornis: Die Forschergruppe benannte in den Jahren 1997 und 1999 insgesamt drei weitere Arten, von denen zwei jedoch umstritten sind. Mit Liaoningornis im Jahr 1996 und Changchengornis im Jahr 1999 kamen von derselben Fundstätte auch weitere neue Vogelgattungen hinzu. Bedauerlicherweise gingen Ende der 1990er-Jahre der Wissenschaft durch die Tätigkeit von Raubgräbern der paläontologischen Forschung viele Vogelfossilien verloren. Bis es gelang, die wichtigsten Fundstellen um Beipiao vor illegalen Fossilienjägern und -händlern zu schützen, waren hunderte Exemplare ausgegraben und verkauft, ohne wissenschaftlich untersucht worden zu sein.
Bei der Erstbeschreibung von Confuciusornis glaubte man, die Gesteinseinheiten der Jehol-Gruppe, zu denen die Fundschichten bei Sihetun und Jianshangou gehören, hätten ein vergleichbares Alter wie die Solnhofener Plattenkalke, aus denen der Urvogel Archaeopteryx stammt. Im Jahr 1999 veröffentlichte Ergebnisse radiometrischer Datierungen der Gesteine belegen jedoch für die gesamte Jehol-Gruppe ein geringeres Alter (Unterkreide gegenüber Oberjura). Der im Vergleich zu Archaeopteryx anatomisch modernere Vogel Confuciusornis war demnach auch der eindeutig jüngere.
Confuciusornis’ Anatomie im Vergleich
Seit den 1980er-Jahren, als Archaeopteryx zeitlich und morphologisch von den anderen damals bekannten fossilen Vögeln isoliert war, wurden zahlreiche Mosaikformen entdeckt, welche die bestehende Lücke füllten, Confuciusornis ist eine davon. Dem Urvogel ähnlich, zeigt das Erscheinungsbild von Confuciusornis ein auffälliges Mosaik aus urtümlichen Merkmalen, also solchen, die bereits bei den Vorfahren der Vögel (den theropoden Dinosauriern) ausgebildet waren, und abgeleiteten Merkmalen, die im Verlauf der Vogelevolution neu hinzukamen. Sie sind in Tabelle 1 aufgelistet. „Nichtvogeltheropode“ ist ein aus der englischen Sprache kommender Begriff für theropode Dinosaurier im herkömmlichen Sinne ohne Einbeziehung ihrer Nachfahren, der Vögel.
| Tabelle 1: Urtümliche und abgeleitete Merkmale bei Confuciusornis im Vergleich zu anderen Gruppen | ||||
| anatomisches Merkmal | theropode Dinosaurier („Nichtvogeltheropoden“) | Archaeopteryx | Confuciusornis | Höhere Vögel (Ornithothoraces) |
|---|---|---|---|---|
| Zähne | bei vielen | + | − | bei vielen Kreide-Vögeln |
| zwei Hinterhauptsöffnungen (diapsider Schädel) | + | − | + | - |
| knöcherner Schwanz verkürzt, letzte Schwanzwirbel zu Pygostyl verschmolzen | bei wenigen | − | + | + |
| Gabelbein (Furcula) | bei einigen (bumerangförmig) | bumerangförmig | bumerangförmig | spitzwinkelig (V-förmig, gabelförmig) |
| Rabenbein (Coracoid) ist lang, strebenförmig | − | − | + | + |
| Brustbein (Sternum) mit Brustbeinkiel (Carina) | − | − | − (aber bei einigen mit niedrigem Kamm) | + (bei flugfähigen Vögeln) |
| Sternalrippen (Rippen, die mit Brustbein verbunden) | bei einigen | − | + | + |
| Bauchrippen (Gastralia) | + | + | + | - |
| Rippen mit Hakenfortsatz (Processus uncinatus) | bei einigen | − | + | bei vielen |
| Oberarm mit einem ausgedehnten perforierten Deltopectoralkamm (autapomorph) | − | − | + | - |
| Mittelhandknochen I–III zu Carpometacarpus verschmolzen | − | − | + (Verschmelzung nur nahe der Handwurzel) | + |
| Mittelfußknochen II–IV sind zu Laufbein (Vogellauf, Tarsometatarsus) verschmolzen | bei einigen | + | + | + |
| Mittelfußknochen V vorhanden | + | + | + | - |
| asymmetrische Schwungfedern | bei einigen | + | + | + |
| Daumenfittich (Alula) | bei wenigen | − | − | + |
Der Schädel zeigt eine ungewöhnliche Merkmalskombination: Der Schnabel von Confuciusornis war wie bei heute lebenden Vögeln unbezahnt, während zeitgleiche modernere Vogelgattungen wie Liaoningornis und Yanornis noch Zähne besaßen. Hingegen war die Schläfenregion so wie bei diapsiden Reptilien ausgebildet: Zwei paarige Schläfenfenster (das Infratemporal- und das Supratemporalfenster) lagen hinter der Augenöffnung (Orbita). Bei allen anderen Vögeln einschließlich Archaeopteryx sind solche Schläfenfenster nicht nachgewiesen.
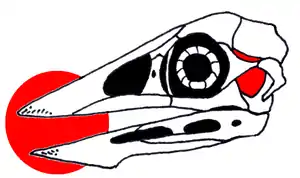
Der Flugapparat von Confuciusornis war gegenüber dem von Archaeopteryx fortgeschritten: Das Brustbein (Sternum) zeigt bei manchen Exemplaren einen flachen Kamm, der an den Brustbeinkiel (Carina) moderner Vögel erinnert. Als Ansatzstelle für die wichtigsten Flugmuskeln zum Flügelauf- und -abschlag ist bei allen rezenten flugfähigen Vögeln ein ausgedehnter Brustbeinkiel entwickelt. Das im Vergleich zu Archaeopteryx verlängerte Rabenbein (Coracoid) zeigt ebenfalls den Umbau der Schultermuskulatur in Richtung moderner Vögel an.
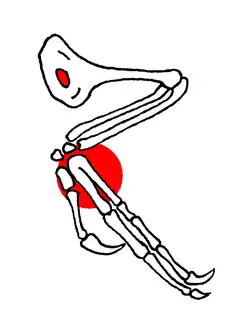
Der Brustkorb von Confuciusornis wurde verstärkt durch Hakenfortsätze an den Rippen und durch so genannte Sternalrippen, das heißt mit dem Brustbein verbundene Rippenelemente. Das Gabelbein hingegen hatte wie bei Archaeopteryx und verschiedenen (Nichtvogel-)Theropoden keine gabelförmige, sondern eine bumerangähnliche Form.
In zeichnerischen Rekonstruktionen (siehe auch das Bild in der obigen Box) wirkt die Hand von Confuciusornis eher urtümlich. Tatsächlich waren die Mittelhandknochen (Metacarpalia) II und III nahe der Handwurzel zusammen mit dem halbmondförmigen Handwurzelknochen zu einem Carpometacarpus verschmolzen. Bei modernen Vögeln sind die Verschmelzungen der Hand so weitreichend, dass nur noch ein Fingerglied des ersten Fingers frei beweglich ist: Der „Daumen“ trägt den so genannten Eck- oder Daumenfittich (Alula), dessen Abspreizen bei bestimmten Flugmanövern zur Manipulation des Luftstroms wichtig ist. Einen solchen Daumenfittich besaßen Confuciusornis und Archaeopteryx nach bisheriger Erkenntnis noch nicht. Dieses Merkmal ist bis auf eine Ausnahme (Microraptor) erst bei Vögeln der höher entwickelten Gruppe Ornithothoraces wie beispielsweise Eoalulavis („früher Alula-Vogel“) ausgebildet.
Der Oberarm von Confuciusornis endet in einem ausgedehnten Pectoralkamm, der eine ovale Öffnung aufweist. Diese Öffnung, deren Funktion ungeklärt ist, hielt man für eine Autapomorphie, also für das Exklusivmerkmal dieser Gattung. Im Jahr 2002 wurde der Vogel Sapeornis beschrieben, der ebenfalls aus den Gesteinen der Jehol-Gruppe stammt und ein ähnliches Oberarmfenster besaß. Demnach könnte dieses Merkmal unter frühen Vögeln weiter verbreitet sein als zuerst angenommen.
Wichtige Neuerungen, die Confuciusornis (und Sapeornis) von noch urtümlicheren Vögeln wie Archaeopteryx, Rahonavis und Jeholornis unterschieden, waren die Verkürzung des knöchernen Schwanzes und die Ausbildung eines Pygostyls durch Verschmelzen der letzten Schwanzwirbel. Ein derartiger Umbau des Schwanzskeletts war für die Flugnavigation von Vorteil.
Systematik
Arten der Gattung Confuciusornis
Von den sechs bisher beschriebenen Arten der Gattung Confuciusornis sind nach Auffassung der Forschergruppe um Luis M. Chiappe zwei ungültig:[2] Confuciusornis suniae und Confuciusornis chuonzhous beruhen auf der Fehlinterpretation innerartlicher Unterschiede von Confuciusornis sanctus. Alle Exemplare, die den beiden Arten zugerechnet wurden, gehörten demnach wie die Mehrheit aller Funde zu C. sanctus.
Confuciusornis dui, die vierte 1999 beschriebene Art, unterscheidet sich von C. sanctus unter anderem in den Handproportionen, in der Form und der Ausprägung mehrerer Schädelelemente sowie in der Form des Brustbeins und der am Brustbein ansetzenden Rippen. 2009 und 2010 wurden zwei weitere Arten beschrieben, Confuciusornis feducciai benannt nach dem US-amerikanischen Paläornithologen Alan Feduccia[3] und Confuciusornis jianchangensis[4].
Gattungen der Familie Confuciusornithidae
Zur Familie Confuciusornithidae zählt neben Confuciusornis die Gattung Changchengornis, die sich von der ersteren unter anderem durch einen kürzeren Schnabel, das Fehlen eines Oberarmfensters, die Form einiger Schultergürtel-Elemente und die Länge der rückwärtsgerichteten ersten Zehe (Hallux) unterscheidet. Der längere Hallux ist ein Hinweis dafür, dass Changchengornis Äste besser umgreifen konnte als Confuciusornis.
Gemeinsame Merkmale aller Mitglieder der Familie Confuciusornithidae sind das Fehlen von Zähnen, die nahezu viereckige Ausprägung des Deltopectoralkamms am Oberarm, der V-förmige hintere Rand des Brustbeins und die Größenverhältnisse der Handkrallen.
Die im Jahr 2001 als Jinzhouornis beschriebenen Vogelfossilien stammen ebenfalls aus den Gesteinen der Jehol-Gruppe und sind der Familie Confuciusornithidae zuzurechnen. Bisher liegt noch keine detaillierte Beschreibung dieser Gattung in englischer Sprache vor.
Verwandtschaftsverhältnisse mesozoischer Vögel
| Aves (Avialae, Vögel) |
| |||||||||||||||||||||||||||||||||||||||
(Kladogramm in Anlehnung an Zhou 2004.)
Die Mitglieder der Familie Confuciusornithidae gehören innerhalb der Gruppe Pygostylia zu den urtümlichsten Formen. Bei den Ornithothoraces, den höher entwickelten Vertretern dieser Gruppe, ist die Zahl der Rückenwirbel (Thoracalia) reduziert und der erste Finger bildet einen Daumenfittich (Alula) aus (siehe auch Punkt 2). Alle Vertreter moderner Vogelordnungen sind Nachkommen dieser Gruppe.
Flugfähigkeit und Lebensweise
Die Flugfähigkeit von Confuciusornis wird von den verschiedenen paläornithologischen Arbeitsgruppen unterschiedlich eingeschätzt. Confuciusornis besaß große gebogene Krallen an Händen und Füßen. Auf dieser Beobachtung beruht die Idee, dass der Flugapparat von Confuciusornis für das Abheben vom Boden nicht leistungsfähig genug war. Anstatt dessen kletterte Confuciusornis mit Hilfe seiner Krallen auf Bäume, um aus dem freien Fall den Schlagflug zu beginnen.
Dieser Ansicht widerspricht, dass der Hinterfuß unspezialisiert und im Vergleich zu rezenten auf Bäumen lebenden Vögeln schlecht an das Umgreifen von Ästen angepasst war. Außerdem dürften die langen Schwungfedern beim Klettern hinderlich gewesen sein. Gegner der Theorie vom kletternden Confuciusornis halten die verschiedenen Anpassungen des Skeletts an den Vogelflug für ausreichend, um beispielsweise aus dem Lauf vom Boden zu starten. Laut Peters und Qiang sei die Idee, Confuciusornis sei nur eingeschränkt flugfähig gewesen, angesichts der großen Flügel abwegig.[5] Letztere sprechen auch dafür, dass die Vertreter der Gattung gute Segelflieger waren.
Die Arbeitsgruppe um Lianhai Hou hält für wahrscheinlich, dass Confuciusornis ein Pflanzenfresser war.[6] Dalsätt und andere hingegen beschrieben ein 2006 Confuciusornis-sanctus-Exemplar aus der Jiufotang-Formation, das einen direkten Beleg für die Art der Ernährung lieferte:[7] Mehrere Wirbel und Rippen des Teleostiers Jinanichthys im unteren Hals- und vorderen Brustbereich des Vogelskeletts zeigen an, dass Confuciusornis unter anderem Fische fraß. Die Fischknochen waren vermutlich Teil eines Speiballens, den der Vogel kurz vor seinem Tod hochwürgte. Dalsätt und andere interpretieren die Schnabelmorphologie von Confuciusornis als die eines Allesfressers ähnlich den rezenten Krähen – Fische waren ihrer Ansicht nach neben Samen und/oder anderen Pflanzenteilen ein üblicher Teil der Nahrung.
Massenvorkommen von Confuciusornis-Skeletten in den Seeablagerungen der Fundstätte Sihetun weisen darauf hin, dass die Vögel möglicherweise in Schwärmen die Ufer von Seen aufsuchten. Wenn in manchen der Schichten mehr als 40 Exemplare je 100 Quadratmeter vorkommen, könnte die Ursache dafür der gemeinsame Tod infolge einer Naturkatastrophe gewesen sein: Bei einem der häufigen vulkanischen Aschefälle starben die Individuen zu gleicher Zeit und ihre Kadaver wurden vom Seeufer aus in den See eingeschwemmt. Aus dieser Theorie folgt, dass Individuen der Gattung Confuciusornis ähnlich wie Vertreter vieler rezenter Vogelarten in Gruppen zusammenlebten oder zeitweilig zusammentrafen.
Sexualdimorphismus und Paarungsverhalten
Die Exemplare von Confuciusornis unterscheiden sich hinsichtlich ihrer Befiederung: Etwa fünf bis zehn Prozent der Individuen weisen Abdrücke eines Paares verlängerter Schwanzfedern auf. Eine Erklärung für diesen Unterschied könnte laut Ansicht verschiedener Bearbeiter die Geschlechter-Zugehörigkeit sein. Falls diese Theorie stimmt, läge das früheste Indiz für Sexualdimorphismus bei Vögeln vor.
Die verhaltensbiologischen Konsequenzen eines derartigen Geschlechterunterschieds wären vielfältig: Es könnte eine Form von Partnerwahl gegeben haben, bei der Individuen des einen Geschlechts vor Individuen des anderen ihr Gefieder zur Schau stellten. Falls die männlichen Tiere diejenigen mit den langen Schwanzfedern waren, weist das ungleiche Verhältnis der Individuenzahlen darauf hin, dass eine Form von Polygynie („Vielweiberei“) bestand. Die aufwändige Wahl des Geschlechtspartners durch das Weibchen ist unter anderem dann zweckmäßig und ökonomisch, wenn sich das Männchen an der Brutpflege beteiligt, zum Beispiel die Gelege der von ihm begatteten Weibchen beschützt.
Der Unterschied in der Befiederung macht deutlich, dass es bereits bei den Vögeln der unteren Kreidezeit ein komplexes Sozialverhalten gegeben haben dürfte.
Wachstum und Entwicklung
Die Evolution der Wachstumsmuster bei Vögeln wurde anhand der Gliedmaßenknochen von Confuciusornis sanctus und anderer früher Vögel untersucht. Durch das stetige Wachstum eines Knochens von innen nach außen (Akkretion) lassen sich auf ähnliche Weise wie bei Baumringen (siehe Dendrochronologie) Veränderungen im Wachstum während der Entwicklung eines Individuums nachvollziehen. Die Unterscheidung von Wachstumsphasen erfolgt bei der Analyse eines Knochenquerschnitts unter dem Mikroskop.
Die Knochenrinde (Kortikalis) von Confuciusornis besteht aus schnell wachsendem, gut durchblutetem inneren Rindengewebe, das dem Fibrolamellargewebe heutiger Vögel gleicht, und einem langsam wachsenden äußeren Rindengewebe. Letzteres war intern stärker zoniert sowie von weniger Blutgefäßen durchsetzt, es repräsentiert das spätere Wachstum des Vogels. De Ricqlès und andere zeigen 2003 anhand knochenhistologischer Untersuchungen, dass Confuciusornis in 20 oder etwas weniger Wochen ausgewachsen war.[8] Diese Zeitspanne ist lang verglichen mit den meisten rezenten Vogelarten, aber etwas kürzer als bei den oft größeren Coelurosauriern. Offenbar erreichten frühe Vögel eine Verringerung der Erwachsenenkörpergröße (Verzwergung), in dem sie gegenüber ihren Dinosaurier-Vorfahren besonders die frühe Phase schnellen Wachstums, die durch das innere Rindengewebe repräsentiert wird, verkürzten.
Der Nachweis von Fibrolamellargewebe bei Confuciusornis belegt dennoch, dass, anders als zuvor angenommen,[9] bereits urtümliche Vögel (gleich ihren Vorfahren und Nachfahren) Phasen schnellen Wachstums zeigten; sie also bereits einen hinreichend hohen Metabolismus besaßen, um hohe Wachstumsraten aufrechtzuerhalten. Das langsam wachsende Knochengewebe der Enantiornithiformes, das zunächst als beispielhaft für urtümliche Vögel galt, repräsentiert offenbar eine nachträgliche nochmalige Wachstumsverlangsamung, die bei den ersten Vögeln und bei Confuciusornis derart noch nicht vorlag. Sehr hohe Wachstumsraten, wie sie moderne Vögel zeigen, bildeten sich dann sekundär im Verlauf der Kreidezeit heraus.
Als mögliche Ursache für langsame Wachstumsraten bei frühen Vögeln wird auch eine besondere Entwicklungsstrategie diskutiert: Fossile Embryonen des Kreidevogels Gobipteryx zeigen ein gut ausgebildetes verknöchertes Flügelskelett, das die Schlussfolgerung zulässt, Vertreter dieser Gattung und vielleicht urtümliche Vögel im Allgemeinen seien extrem frühreife Nestflüchter gewesen, die bereits kurz nach dem Schlüpfen fliegen konnten. Gemäß dieser Theorie würde ein Großteil der Energie des Jungvogels für das Fliegen aufgewandt und stünde daher nicht für das Größenwachstum zur Verfügung. Dieser Zustand würde erst bei Vogelarten überwunden, die eine intensive Brut- und Nachwuchspflege betrieben. Dadurch müssten die Jungvögel nicht direkt nach der Geburt flügge und ohne Hilfe der Eltern überlebensfähig sein, sondern könnten einen Großteil der durch Nahrung aufgenommenen Energie in das Wachstum investieren.
Die Bedeutung von Confuciusornis als Element der Jehol-Avifaunen

In der folgenden stratigraphischen Tabelle sind vom Älteren (unten) zum Jüngeren (oben) die Fundschichten der Jehol-Gruppe sowie die jeweils beschriebenen Vogelarten aufgelistet.
| Tabelle 2: Zusammensetzung der Jehol-Avifaunen in stratigraphischer Abfolge | ||||
| Formation | Schichtglied | urtümliche Vögel | Ornithothoraces | |
| Enantiornithes | Ornithurae | |||
| Jiufotang- Formation |
Confuciusornis sp., Sapeornis chaoyangensis, Jeholornis prima |
Sinornis santanensis, Longchengornis sanyanensis, Cuspirostrisornis houi, Largirostrornis sexdentorius, Boluochia zhengi, Longipteryx chaoyangensis |
Chaoyangia beishanensis, Yanornis martini, Yixianornis grabaui, Songlingornis linghensis | |
| Yixian- Formation |
Jingangshan- Schichten |
Confuciusornis sp. | --- | --- |
| Dawangzhangzi- Schichten |
Confuciusornis sp. | Liaoxiornis delicatus, Liaoxiornis sp., Jibeinia luanhera |
--- | |
| Jianshangou- Schichten |
Confuciusornis sanctus, Confuciusornis dui, Changchengornis hengdaoziensis |
Protopteryx fengningensis, Eoenantiornis buhleri |
Liaoningornis longiditris | |
Im Gegensatz zu den anderen Vogelgattungen kam Confuciusornis über mehr als 15 Millionen Jahre, also nahezu während des gesamten Bildungszeitraums, in den Sedimenten der Jehol-Gruppe vor. Diese Beobachtung zeigt, dass sich Confuciusornis trotz seiner vergleichsweise schlecht entwickelten Flugeigenschaften gegen konkurrierende Vögel mit fortschrittlicheren Bauplänen durchsetzen konnte.
In der Abfolge besteht dennoch ein Trend hin zu höher entwickelten Vögeln. Spätestens während der Bildung der Jiufotang-Formation dominierten die Enantiornithes, eine Gruppe, die bis zu ihrem Aussterben am Ende der Kreidezeit in vielen Ökosystemen vorherrschte. Interessanterweise erscheint die Vogelart Jeholornis prima, die dem Urvogel Archaeopteryx noch ähnlicher war als Confuciusornis, erst zum Ende des Bildungszeitraums der Jehol-Gruppe.
Siehe auch: Paläornithologie
Literatur
Allgemeines, Geschichte
- Ursula B. Göhlich, Gerald Mayr: Zu Besuch bei Confuciusornis & Co. in Nordost-China. In: Natur und Museum. 131, Nr. 11, 2001, S. 401–409.
- Erik Stokstad: Exquisite Chinese Fossils Add New Pages to Book of Life. In: Science. 291, Nr. 5502, 2001, S. 232–236, doi:10.1126/science.291.5502.234.
- Carl C. Swisher III, Yuan-Qing Wang, Xiao-Lin Wang, Xing Xu, Yuan Wang: Cretaceous age for feathered dinosaurs of Liaoning, China. In: Nature. 400, Nr. 6739, 1999, S. 58–61, doi:10.1038/21872.
- Zhong-He Zhou, Lianhai Hou: The Discovery and Study of Mesozoic Fossil Birds in China. In: Luis M. Chiappe, Lawrence M. Witmer (Hrsg.): Mesozoic Birds. Above the Heads of the Dinosaurs. University of California Press, Berkeley 2002, ISBN 0-520-20094-2, S. 160–182.
Anatomie, Systematik, Phylogenese
- Luis M. Chiappe, Shu'an Ji, Qiang Ji, Mark A. Norell: Anatomy and Systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of Northeastern China. (PDF; 9,7 MB) In: Bulletin of the American Museum of Natural History. Nr. 242, 1999, S. 98.
- Dieter Stefan Peters: Ein nahezu vollständiges Skelett eines urtümlichen Vogels aus China. In: Natur und Museum. 126, Nr. 9, 1996, S. 198–302.
- Dieter Stefan Peters, Ji Qiang: The diapsid temporal construction of the Chinese fossil bird Confuciusornis. In: Senckenbergiana lethaea. 78, Nr. 1–2, 1998, S. 153–155, doi:10.1007/BF03042766.
- Andrea Goernemann: Osteologie eines Exemplars von Confuciusornis aus der unteren Kreide von West-Liaoning, China. In: Archaeopteryx. 17, 1999, S. 41–54.
- Andrzej Elzanowski, Albrecht Manegold, Dieter Stefan Peters: Redescription of a skull of Confuciusornis sanctus. In: Archaeopteryx. 23, 2005, S. 51–55.
- Lianhai Hou, Zhong-He Zhou, Larry D. Martin, Alan Feduccia: A beaked bird from the Jurassic of China. In: Nature. 377, Nr. 6550, 1995, S. 616–618, doi:10.1038/377616a0.
- Qiang Ji, Luis M. Chiappe, Shu'an Ji: A new Late Mesozoic confuciusornithid bird from China. In: Journal of Vertebrate Paleontology. 19, Nr. 1, 1999, S. 1–7.
- Larry D. Martin, Zhong-He Zhou, Lian-Hai Hou, Alan Feduccia: Confuciusornis sanctus compared to Archaeopteryx lithographica. In: Naturwissenschaften. 85, Nr. 6, 1998, S. 286–289, doi:10.1007/s001140050501.
- Qicheng Wu: Fossil Treasures from Liaoning. Geological Publishing House, Peking 2002, ISBN 0-520-20094-2.
- Zhong-He Zhou: The origin and early evolution of birds: discoveries, disputes and perspectives from fossil evidence. In: Naturwissenschaften. 91, Nr. 10, 2004, S. 455–471, doi:10.1007/s00114-004-0570-4.
Flugfähigkeit, Lebensweise, Sozialverhalten, Reproduktion
- Luis M. Chiappe, Shu'an Ji, Qiang Ji, Mark A. Norell: siehe oben
- J. Dalsätt, Z. Zhou, F. Zhang, P. G. P. Ericson: Food remains in ‘Confuciusornis sanctus’ suggest a fish diet. In: Naturwissenschaften. 93, Nr. 9, 2006, doi:10.1007/s00114-006-0125-y.
- J. David Ligon: The Evolution of Avian Breeding Systems. Oxford University Press, Oxford 1999, ISBN 0-19-854913-X.
- Dieter Stefan Peters, Qiang Ji: Mußte Confuciusornis klettern? In: Journal für Ornithologie. 140, Nr. 1, 1999, S. 41–50.
- David J. Varricchio, Frankie D. Jackson: Origins of avian reproduction: answers and questions from dinosaurs. In: Palaeovertebrata. 32, Nr. 2–4, 2003, S. 149–169.
Wachstum, Individualentwicklung
- Anusuya Chinsamy, Andrzey Elzanowski: Evolution of growth patterns in birds. In: Nature. 412, Nr. 6845, 2001, S. 402–403 doi:10.1038/35086650.
- Kevin Padian, Armand J. de Ricqlès, John R. Horner: Dinosaurian growth rates and bird origins. In: Nature. 412, 2001, S. 405–408, doi:10.1038/35086500.
- A. J. de Ricqlès, K. Padian, J. R. Horner, E.-T. Lamm, N. Myhrvold: Osteohistology of Confuciusornis sanctus (Theropoda: Aves). In: Journal of Vertebrate Paleontology 23, Nr. 2, S. 373–386.
Biostratigraphie der Jehol-Gruppe
- Xiao-lin Wang u. a.: Vertebrate Biostratigraphy of the Lower Cretaceous Yixian Formation in Lingyuan, Western Liaoning and its neighboring southern Nei Mongol (Inner Mongolia), China. In: Vertebrata PalAsiatica. 38, Nr. 2, 2000, S. 81–99.
- Zhonghe Zhou, Paul M. Barett, Jason Hilton: An exceptionally preserved Lower Cretaceous ecosystem. In: Nature. 421, Nr. 6925, 2003, S. 807–814, doi:10.1038/nature01420.
Einzelnachweise
- L. Hou, Z. Zhou, Y. Gu, H. Zhang: Confuciusornis sanctus, a new Late Jurassic sauriurine bird from China. In: Chinese Science Bulletin. 40, Nr. 18, 1995, S. 1545–1551.
- L. M. Chiappe, J. Shu'an, J. Qiang, M. Norell: Anatomy and Systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of Northeastern China. (PDF; 9,7 MB) In: Bulletin of the American Museum of Natural History. Nr. 242, 1999, S. 98.
- Z. Zhang, C. Gao, Q. Meng, J. Liu, L. Hou and G. Zheng. 2009. Diversification in an Early Cretaceous avian genus: evidence from a new species of Confuciusornis from China. Journal of Ornithology 150(4):783-790
- L. Li, J.-q. Wang, and S.-l. Hou. 2010. A new species of Confuciusornis from Lower Cretaceous of Jianchung, Liaoning, China. Global Geology 29(2):183-187
- Dieter Stefan Peters, Ji Qiang: The diapsid temporal construction of the Chinese fossil bird Confuciusornis. In: Senckenbergiana lethaea. 78, Nr. 1–2, 1998, S. 153–155, doi:10.1007/BF03042766.
- L. Hou, L. D. Martin, Z. Zhou, A. Feduccia, F. Zhang: A diapsid skull in a new species of the primitive bird ‘Confuciusornis’. In: Nature. 399, 1999, S. 680, doi:10.1038/21411.
- J. Dalsätt, Z. Zhou, F. Zhang, P. G. P. Ericson: Food remains in 'Confuciusornis sanctus' suggest a fish diet. In: Naturwissenschaften. 93, Nr. 9, 2006, doi:10.1007/s00114-006-0125-y.
- A. J. de Ricqlès u. a.: Osteohistology of Confuciusornis sanctus (Theropoda: Aves). In: Journal of Vertebrate Paleontology. 23, Nr. 2, 2003, S. 373–386.
- Anusuya Chinsamy, Andrzey Elzanowski: Evolution of growth patterns in birds. In: Nature. 412, Nr. 6845, 2001, S. 402–403 doi:10.1038/35086650.