Entwicklungsbiologie
Die Entwicklungsbiologie erforscht die Vorgänge, durch die einzelne Organismen wachsen und sich von der einzelnen Zelle zu einem komplexen vielzelligen Organismus entwickeln (Ontogenese). Die Entwicklungsbiologie hat ihren Ursprung in der Embryologie und behandelt heute genetische und epigenetische Prozesse der Selbstorganisation von Zellen auf der Basis ererbter Genregulationsnetzwerke, der physiko-chemischen Eigenschaften von Zellen und Geweben sowie von Umweltfaktoren.
Weitgehend synonym wird der Begriff Entwicklungsgenetik verwendet, er kann aber auch Aspekte der Verhaltensgenetik bezeichnen, vernachlässigt jedoch epigenetische Aspekte der Entwicklung. Die Reifung des Nervensystems untersucht die Entwicklungsneurobiologie. Mit den Zusammenhängen zwischen Entwicklungsbiologie und Evolution befasst sich die Evolutionäre Entwicklungsbiologie (EvoDevo).
Entwicklung der Tiere
Entwicklung: Definition
Ernst Haeckel definierte Entwicklung als den gesamten Vorgang von der befruchteten Eizelle bis zum erwachsenen Lebewesen.[1] Heute sieht man Entwicklung als genetischen und epigenetischen Prozess der Selbstorganisation von Zellen auf der Basis ererbter genetischer Netzwerke, der physiko-chemischen Eigenschaften von Zellen und Geweben sowie von Umweltfaktoren.
Faktoren, Mechanismen und Hauptstadien der Entwicklung
Die Entwicklung verläuft als komplexes Wechselspiel erstens verschiedener genetisch bestimmter Entwicklungsfaktoren (Genprodukte) wie z. B. Morphogene, Adhäsionsmoleküle, Wachstumsfaktoren, Entwicklungsgene, aber auch epigenetischer Entwicklungsfaktoren wie Chromatinveränderungen und Faktoren der inneren und äußeren Umwelt des Embryos sowie zweitens von Entwicklungsmechanismen wie z. B. Induktion, Zellproliferation, Zelldifferenzierung, Zellmigration, Signaltransduktion, Apoptose, Zellwachstum oder Musterbildung.
Genprodukte
Zu den genetisch bestimmten Entwicklungsfaktoren zählt eine Reihe entwicklungswirksamer Gene bzw. Genprodukte (Proteine). Ein Morphogen ist eine Substanz, die die räumliche Ordnung der Zelldifferenzierung und damit indirekt die Formbildung kontrolliert. Morphogene können damit Musterbildung während der Morphogenese bewirken.[2] Adhäsionsmoleküle sind Proteine, die eine Haftung an einer Zelle oder an einem Substrat bewirken (Bsp. Cadherine, Integrine). Ein Wachstumsfaktor ist ein Protein, das im extrazellulären Raum vorhanden sein muss, damit bestimmte Zelltypen wachsen und sich normal entwickeln.[3] Hierzu zählen etwa Knochenmorphogenetische Proteine. Entwicklungsgene gibt es in Klassen. Man unterscheidet Maternaleffekt-Gene zur Festlegung der Polarität des Embryos, Segmentierungsgene zur segmentalen Gliederung des Embryos, Kompartiment-Gene für Feldunterteilungen, Historegulationsgene für zellulare Differenzierung sowie als eine der wichtigsten Klassen homöotische Gene zur Identitätsbestimmung von Segmenten und Feld-Gene für die Festlegung von Organ-Anlagefelder (z. B. eyeless). Mutationen innerhalb homöotische Gene können zur Umwandlung oder dem Verlust von Organstrukturen oder Körperabschnitten führen. Diese Gene enthalten als charakteristische Sequenz stets eine Homöobox, die für einen Transkriptionsfaktor codiert. Die bekanntesten homöotischen Gene sind die Hox-Gene.
Epigenetische Faktoren
Epigenetische Faktoren sind Einflüsse, die epigenetische Veränderungen von Zellen hervorrufen. Es werden epigenetische Faktoren der inneren und der äußeren Umwelt unterschieden. Zur inneren Umwelt zählt die chemische Zusammensetzung der inneren Umgebung des Embryos, im Besonderen die umgebenden Zellen und Interaktionen zwischen Zellen und Zellpopulationen, aber auch räumlich-geometrische Bedingungen wachsender Gewebe- und Organanlagen. Zur äußeren Umwelt zählen die chemische Zusammensetzung der Umwelt, die Ernährung, sowie physikalische Bedingungen wie Licht, Mechanik, Schwerkraft (Eipolarisierung) und Temperatur (Geschlechtsdetermination bei Schildkröten und Krokodilen).
Mit dem Ausdruck Epigenese werden die graduellen Prozesse der embryonalen Morphogenese von Organen beschrieben. Diese beruhen auf Mechanismen auf der Ebene von Zellen und Zellverbänden, das sind Turing-Mechanismen oder allgemein Musterbildungsprozesse in der Biologie. Beispiele hierfür findet man etwa bei der Erklärung der embryonalen Extremitätenentwicklung der Wirbeltiere.
Entwicklungsmechanismen
Eine Reihe spezifischer Entwicklungsmechanismen, die Entwicklungsphysiologie, bestimmt den spezifischen Verlauf der Morphogenese: Induktion ist die Beeinflussung der Entwicklung einer embryonalen Zielgruppe durch eine andere,[3] die Induktionssubstanzen freisetzt, z. B. die Induktion der Linsenentwicklung beim Wirbeltierauge durch ektodermales Gewebe. Zellproliferation sind wiederholte, schnelle und kontrollierte Zellteilungen, die zu Gewebewachstum führen. Zelldifferenzierung meint einerseits die Differenzierung von Zellen im Vergleich untereinander, andererseits die individuelle Zellentwicklung und damit die Differenzierung einer Zelle im Lauf der Entwicklungszeit. Die Determination, das heißt Programmierung einer Zelle erfolgt auf einem bestimmten Entwicklungsweg. Eine programmierte, aber noch nicht endgültig differenzierte Zelle erhält die Bezeichnung "-blast", die reife, ausdifferenzierte Zelle heißt "-cyt". Es wird zwischen der reversiblen Spezifizierung einer Zelle und ihrer nicht reversiblen Differenzierung bzw. Determination oder Commitment unterschieden.[4] Die Zelldifferenzierung ist zusammen mit der Zellteilung (Mitose) eine notwendige, aber nicht hinreichende Voraussetzung dafür, dass ein entwickelnder, mehrzelliger Embryo seine Form erhalten kann (Morphogenese). Molekularbiologisch äußert sich Zelldifferenzierung darin, dass nicht das gesamte Genom gleichzeitig zur Proteinerzeugung genutzt wird, sondern nur die für den jeweiligen Zelltyp relevanten Gene aktiv sind. Man spricht von differenzieller Genexpression.[3]
Stammzellen sind Zellen, die ihre Teilungsfähigkeit aufrechterhalten haben und erst nach der Vermehrung weiter differenzieren. Nach der asymmetrischen Teilung bleibt die eine Tochterzelle weiterhin Stammzelle, die andere schlägt einen neuen Weg ein.[4] Stammzellen werden nach ihrem Entwicklungspotenzial unterschieden: Totipotente Stammzellen (Eizelle) können den gesamten Organismus hervorbringen. Pluripotente Stammzellen sind embryonale Stammzellen aus dem Blastoderm, die noch auf keinen bestimmten Zelltyp festgelegt sind. Sie bringen Zellen verschiedener Art hervor, die jedem der Keimblätter zugehören können. Multipotente Stammzellen können verschiedene Zellen einer bestimmten Linie hervorbringen, d. h. verschiedenen Derivate eines Keimblatts. Unipotente Stammzellen differenzieren zu einem einzigen Zelltyp, z. B. Stammzellen der Epidermis der Haut. Mit dem Fortschritt der Entwicklung erfolgt eine zunehmende Einschränkung der Entwicklungspotenz von Zellen von totipotent über pluripotent und multipotent zu unipotent. Unter Zellmigration versteht man die aktive Ortsveränderung von Zellen oder Zellverbänden, beispielsweise bei der Entstehung des Nervensystems. Signaltransduktion beschreibt die Ereigniskette zur Überführung einer Botschaft vom Zelläußeren über die Zellmembran hinweg ins Zellinnere. Die Signaltransduktion kommt in Gang, wenn ein extremes Signalmolekül, ein Ligand, an Membranverankerte Rezeptormoleküle bindet.[2] Apoptose ist der kontrollierte, programmierte Zelltod ohne Beeinträchtigung des umgebenden Gewebes und betrifft Gewebe, das nur in einer bestimmten Entwicklungsphase benötigt und anschließend zu Vesikeln abgebaut und durch Makrophagen vernichtet wird, z. B. die mesenchymalen Zwischenräume zwischen Fingern und Zehen bei Wirbeltierextremitäten oder der Verlust des Schwanzes der Kaulquappe.
Selbstorganisation als Entwicklungsprinzip meint das Entstehen von Ordnung aus anfänglicher Unordnung oder Strukturierung aus Nichtstrukturierung von Zellen und Geweben. Hierbei kann eine initiale, unter Umständen geringe Veränderung eines Parameters wie einer Genaktivität oft auf dem Weg über Schwellenwerteffekte eine nicht lineare, nicht chaotische Reaktion des gesamten Systems bewirken. Vorgänge der Musterbildung schließlich lassen wohlgeordnete und reproduzierbare räumliche Muster verschieden differenzierter Zellen entstehen.[2] Dies kann sowohl zweidimensional sein, etwa bei der Bildung von Farbmustern auf Haut (Fische) und Fellen (Zebra, Gepard), Schmetterlingsflügeln oder Vogelgefiedern (Kreise, Rhomben, Streifen), als auch dreidimensional, etwa bei der Wirbeltier-Extremitätenentwicklung oder bei der Ausbildung neuronaler Muster. Theoretische Grundlage für Musterbildung ist das Turingmodell oder Turing-Mechanismus, ein chemisches Reaktions-Diffusionsmodell, in dem Turing 1952 erstmals die chemische Basis der Morphogenese beschrieb.[5] Das Turingmodell wurde zu einem Modell mit lokalem Aktivator und lateraler Inhibition (LALI-Modell) ausgeweitet.[6] LALI-Modelle lösen sich von chemischer Reaktion und erlauben Patterning auf Zellebene durch Zellinteraktion. Musterbildungsprozesse sind daher nicht im Detail genetisch determiniert. Neben genetisch notwendigen Bedingungen liegt die hinreichende Information der eigentlichen Musterbildungsprozesse auf Zellebene und ist damit epigenetisch.
Im Verlauf der Entwicklung kommt es zur Ausbildung differenzierter Zellen für spezifische Gewebetypen (Haut, Muskel, Nerven, Organe etc.). Es gibt Regionen des Embryos, in denen in einer ganz bestimmten Phase der Entwicklung ein oder wenige ganz bestimmte Gene der Zellen exprimiert und bestimmte Signalproteine produziert werden.[7] Die Fähigkeit, unterschiedlich konservierte Kernprozesse an unterschiedlichen Orten zu bestimmten Zeiten im Organismus zu aktivieren und diese Reaktionsräume zu schaffen, wird Kompartimentierung genannt. Ein Insektenembryo bildet in der mittleren Phase der Entwicklung ca. 200 Kompartimente aus. Die Ausprägung dieser Kompartimente ist die eigentliche Aufgabe der Hox-Gene.[7]
Entwicklungsstadien
Die Hauptstadien der Entwicklung sind in zeitlicher Folge Zygote → Furchung → Gastrulation → Organogenese → Fetogenese → Larvalstadium (Larvalentwicklung mit Metamorphose) → Juvenilstadium (Wachstum) → Adultstadium → (Maturität), Altern → Tod. Dabei werden die Phasen Zygote, Furchung und Gastrulation als Frühentwicklung bezeichnet.
Ausgewählte Entwicklungsstadien der embryonalen Mausentwicklung (E = Embryonic day)[8]
| Phase | Ereignis |
|---|---|
| E1.0 | Befruchtung |
| E2.0 | Erste Furchung nach 24 Stunden. Holoblastische Furchung. Zellzyklus 12 Stunden, relativ lange im Vergleich zu Xenopus. 4–16 Zellen |
| E3.5 | Embryo. Blastocyste besteht aus Trophoblast (Ernährungsteil), innerer Zellmasse und Blastocoel |
| E4.0 | Epiblast und primitives Endoderm. Embryoblast und aembryonaler Pol erkennbar. aembryonaler Pol = hauptsächlich Epiblast, embryonaler Pol = Blastocoel |
| E4.5 | Implantation der Blastocyste in Gebärmutterschleimhaut. |
| E5.0 | Erste Signale zur Etablierung der antero-posterioren Körperachse. |
| E5.5 | Invagination (Einstülpung) des prospektiven Endoderms in den inneren, flüssigkeitsgefüllten Hohlraum (Blastocoel) der Blastula. Innenliegender Teil = Endoderm, außenliegender Teil = Ekoderm. Einengung der Blastula. Einstülpung = Urdarm (Archenteron); Öffnung = Urmund (später After) |
| E6.0 | Extraembryonales Ektoderm in der Einstülpung. Proamniumhöhle = Sekundäre Leibeshöhle |
| E6.5 | Zylindrische Struktur des Embryos. Beginn der Gastrulation |
| E7.0 | Amnion |
| E7.5 | Neuralplatte. Beginn der Neurulation. Herzschlauch |
| E8.0 | Erste 8 Somitenpaare. Ohrplakode |
| E9.0 | Urkeimzellen-Wanderung. 16 Somitenpaare. Vordere Extremitätenknospen. Hintere Knospe ca. einen halben Tag später. |
| E10.0 | Linsenvesikel im Auge separiert. |
| E11.0 | Extremitätenentwicklung. Früheste Anzeichen von Zehen. 45–47 Somitenpaare |
| E11.5 | Augenabgrenzungen deutlich sichtbar. Vordere Extremitätenknospe formt Handplatte |
| E12.0 | Zahnknospen |
| E13.0 | 53–55 Somitenpaare. Haarfollikel über Auge und Ohr. Ellbogen. Handgelenksknochen. Gehörknöchelchen. |
| E14.0 | 56–60 Somitenpaare. Gaumen. Kappenstadium der Zähne |
| E15.0 | Innenohr: Hörschnecke (Cochlea). Gonaden. Apoptose der Zehenzwischenräume. Pankreas. Lunge. Glockenstadium der Zähne |
| E15.5 | Olfaktorische Entwicklung (Geruchssinn) |
| E16.0 | Vorhirn, Gehirnumstrukturierungen |
| E17.0 | Hautverdickung und -faltungen, lange Schnurrhaare |
| E18.0 | Augen durch Augenlider schwach sichtbar |
| E19.0 | Geburt |
Wissenschaftsgeschichte der Embryologie
Entwicklung wichtiger Modellorganismen der Tiere
Die Entwicklung wird anhand von Modellorganismen beschrieben. Modellorganismen müssen dafür einer Reihe von Anforderungen genügen, das sind unter anderem umfangreiche Zucht, kurze Generationsdauer, einfache und kostengünstige Haltung, das Vorhandensein natürlicher Mutanten, induzierbare Eiablage, hohe Nachkommenzahl sowie einfache Beobachtung und Untersuchungsmethoden. Als Modellorganismen der Entwicklungsbiologie der Wirbeltiere dienen der Zebrabärbling (Danio rerio), der Glatte Krallenfrosch (Xenopus laevis), das Huhn (Gallus gallus) und die Hausmaus (Mus musculus). Bei den Wirbellosen gilt als Referenzmodell für die Befruchtung und frühe Entwicklung der Seeigelkeim. Ferner werden für Entwicklungsstudien die Taufliege Drosophila melanogaster, der Fadenwurm Caenorhabditis elegans oder Pristionchus pacificus, der Reismehlkäfer Tribolium castaneum sowie der Krebs Partiale hawaiensis als Modellorganismen verwendet.
Gametogenese
Die Keimzellen (Gameten) bilden als Resultat meiotischer Zellteilung die haploiden Zellen, die der geschlechtlichen Fortpflanzung dienen und im jeweiligen elterlichen Organismus als weibliche Eizelle (Oocyte) und männliches Spermium heranwachsen. Die Haploidie der Gameten bildet die Voraussetzung für die genetische Neukombination bei der Befruchtung und damit für die genetische Vielfalt in der Population.
Befruchtung
Die Befruchtung oder Fertilisation ist die Verschmelzung von männlichen und weiblichen Keimzellen. Es kommt zur Vermischung des elterlichen genetischen Materials im Rahmen der geschlechtlichen Fortpflanzung. Das ist der eigentliche komplexe Initiationsvorgang der embryonalen Entwicklung. Spermium und Eizelle ziehen sich mit Hilfe diffusibler Substanzen an, die von der Eizelle freigesetzt werden und auf das Spermium chemoattraktiv wirken. Im Weiteren gelingt es einem Spermium, an die Eizelle anzudocken. Die Membranen beider Keimzellen verschmelzen. Der männliche Vorkern dringt in die Eizelle ein. Es kommt zu einer Änderung des Membranpotenzials der Eizelle, wodurch das Eindringen weiterer Spermien (Polyspermie) verhindert wird. Nach dem Eindringen des Spermiums in den Zellkern der Eizelle wird die Eizelle durch die Verschmelzung der beiden Zellkerne befruchtet und besitzt nun als Zygote Erbmaterial des Vaters und der Mutter.[4]
Furchung
Nach der Befruchtung beginnt die Embryogenese mit der Furchung als erster Phase. Die Furchung ist die rasche Abfolge wiederholter Zellteilungen durch Abschnürungen der Zygote. Der Vorgang verläuft für alle Zellen synchron. Der menschliche Keim besteht nach 7 Tagen und 7 Furchungsrunden aus etwas über 100 Zellen, von denen etwa zehn Embryoblastzellen darstellen.[3]
Der Zweck der Furchung sind schnelle Zellteilungen am Beginn der Entwicklung, ausgehend von der Zygote. Der entstehende Zellverband bleibt dabei gleich groß, denn zusätzliches Zellplasmamaterial wird nicht bereitgestellt. Nach der Geometrie der einzelnen Teilungen unterscheidet man verschiedene Furchungstypen. Die Stadien der Furchung führen zum Entstehen der Morula, einer mit Zellen gefüllten Kugel. Danach beginnt die Entwicklung der Blastula, oder bei den Säugern der Blastocyste, einer flüssigkeitsgefüllten Hohlkugel. Der entstandene Hohlraum in ihrem Innern, das Blastocoel, ist die primäre Leibeshöhle. Die Blastula ist nach sechs Zellteilungszyklen in fünf Territorien gegliedert, die bei dem Frosch Xenopus laevis bereits alle drei Keimblätter enthalten.
Bei den Säugern treten schon in der frühen Phase evolutionär Strukturen auf, die sie von den sich aquatisch entwickelnden Amphibien unterscheiden. So entsteht in der inneren Zellmasse der Blastocyste durch Aussparung die Amnionhöhle, die den Embryo aller Amniontiere schützt und ihm eine von einem Lebensraum in äußerem Wasser unabhängige Entwicklung erlaubt (Fruchtwasser). Bei Fischen, Reptilien und Vögeln wird ein Dottersack gebildet, ein extraembryonales, häutiges Gebilde zur Ernährung des Embryos. Bei den Säugern ist dieser nur in reduzierter Form noch vorübergehend vorhanden.[2] Denn die Blastocyste eines Säugers differenziert sich nach der Furchung in Trophoblast, eine extraembryonale Zellschicht, die für die nährende Verbindung zur Gebärmutter sorgt, und Embryoblast. Aus dem Embryoblasten geht der eigentliche Embryo hervor. Im Unterschied hierzu wird bei den Amphibien die gesamte Blastula zur Larve, sie entwickeln also keine embryonalen Hüllgewebe.
Die embryonalen Zellen des Embryoblasten eines Säugerkeims sind noch pluripotent und zeigen keine erkennbaren Unterschiede untereinander, sie heben sich aber nun von den Trophoblastzellen ab. Totipotente Zellen liegen nur in früheren Stadien vor, beim Menschen womöglich noch im Achtzellstadium.[3]
Gastrulation und Keimblattbildung
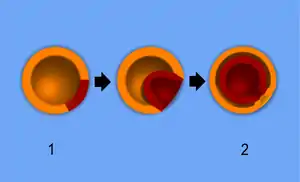
Die Gastrulation führt auf alternativen Wegen zur Einstülpung der Blastula an ihrem vegetativen Pol und zu Zellverschiebungen in das Innere der Blastula. Die Blastula wird im Zuge der Gastrulation reorganisiert. Bei den Neumündern (Deuterostomia), zu denen die Wirbeltiere gehören, ist die Einstülpungsstelle der Urmund, der zum After wird, während am gegenüberliegenden Pol der Mund neu durchbricht (Neumund), womit die Gastrulation beendet ist.[3] Durch die Gestikulation entsteht eine mehrschichtige Organisation, die bei den bilateral gebauten Tieren (Bilateria) drei Keimblätter, bei diploblastischen Tieren wie Nesseltieren und Rippenquallen zwei Keimblätter aufweist. Von dieser Grundorganisation aus nimmt die Organbildung (Organogenese) ihren Anfang. Man unterscheidet zwischen Entoderm, Mesoderm und Ektoderm. Keimblätter sind die ersten embryonalen Zellschichten, aus denen jeweils nicht nur eines, sondern mehrere Gewebe und Organe hervorgehen. Aus dem Entoderm bilden sich Verdauungstrakt, Pankreas, Leber, Atmungstrakt, Schilddrüse und Harnwege. Das Mesoderm führt zur Entwicklung von Unterhaut, Chorda, Knorpel, Knochen, Skelettmuskulatur und Blutzellen. Das Ektoderm ist hauptverantwortlich für das Entstehen von Nieren, Haut und Nervensystem. Die final ausgebildeten Organe besitzen aber in der Regel jeweils Bestandteile aller drei Keimblätter.[4] Das Mesoderm bildet zwei weitere Schichten, wobei die eine Schicht das Innere der Körperhöhle abdichtet und die andere die Außenseite des Darms bildet. Weitere Mesoderm-Kompartimente bilden jeweils Zelllieferanten für neue Organabschnitte.
Festlegung der Körperachsen
Die Entscheidung, wo das vordere und hintere Ende, wo oben und unten, rechts und links im Embryo angelegt werden (Körperachsen), gehört zu den grundlegenden frühen Entwicklungsdeterminationen und wird bei manchen Tieren teilweise bereits maternal in der Eizelle festgelegt. Prinzipiell gibt es unterschiedliche Mechanismen für die Achsenfestlegung: Externe Orientierungshilfen können Schwerkraft oder Licht sein, so etwa beim Hühnchen, bei dem die Kopf-Schwanz-Polarität (cranial-caudale Achse) durch Schwerkraft und durch die Richtung bestimmt wird, mit der das Ei beim Transport durch den Eileiter in Rotation gesetzt wird, während die Rücken-Bauchpolarität (dorso-ventrale Achse) in der Struktur des Eis selbst liegt. Bei Xenopus existiert eine animal-vegetative Eipolarität. Der Eintrittsort des Spermiums ruft in Verbindung mit Schwerkraft eine Symmetriebewegung hervor. Der Schwanzpol entsteht in der Folge diagonal zur Eintrittsstelle des Spermiums. Maternale Determinanten legen bei D. melanogaster bereits vor der Befruchtung weitgehend über cytoplasmatische Determinanten (mRNA) im Ei selbst die antero-posteriore Achse fest. Die Bilateralsymmetrie liegt hier also weitgehend unter der Kontrolle des mütterlichen Genoms. Beim Zebrafisch ist die animal-vegetative Achse wegen der waagerechten Lage des Eis horizontal orientiert. Wenig vorprogrammiert in Bezug auf die Achsenbildung ist das Ei der Maus.[2] Die Polarität der Säuger ist auch nach der Furchung offensichtlich noch nicht festgelegt, wenn auch Untersuchungen dafür sprechen, dass der Eintrittsort des Spermiums auch hier eine Rolle spielt.[3]
Organogenese
Die Organogenese ist der Prozess in mehrzelligen tierischen Organismen, bei dem die Entwicklung der Organanlagen im Laufe der Embryogenese abläuft. Die Organogenese folgt auf die Furchung und Gastrulation; an sie schließt die Fetogenese an.
Plazenta
Neurulation
Entwicklung des Nervensystems
Entwicklung der Neuralleistenzellen, Abkömmlinge
Blutgefäßentwicklung, Herzentwicklung
Muskelentwicklung
...
Somitenentwicklung
Extremitätenentwicklung
Entwicklung des Urogenitalsystems
Nierenentwicklung
Augenentwicklung
Lungenentwicklung
Schilddrüsenentwicklung
Entwicklung von Schädel und Gesicht
Zahnentwicklung
Haarentwicklung
...
Geschlechtsdetermination
Vielzellige Organismen besitzen bisexuelle Potenz. Die Geschlechtsdetermination erfolgt hier genetisch über Selektorgene. Diese können auf dem männlichen Chromosom (Y) liegen, wie beim Menschen (Sex determing region zw. SRY-Gen), oder auf dem weiblichen Chromosom (X bzw. W) wie bei der Taufliege. Das ungleiche geschlechtsbestimmende Chromosom heißt Heterosom im Vergleich zu den gleichartigen Autosomen. Genotypische Geschlechtsdetermination geschieht bei den Tieren nicht nach einem einheitlichen Prinzip. Manche Tiere entwickeln Hermaphroditen, etwa C. elegans. Er entwickelt zusätzlich männliche, aber keine weiblichen Individuen. Bei den Anneliden Ophryotrocha puerilis kommt es zuerst zur Ausprägung eines männlichen Phänotyps und später eines weiblichen. Begegnen sich zwei Weibchen, muss sich in einem Duell der schwächere von beiden in ein Männchen zurückentwickeln. Treffen sich zwei Männchen, entwickelt sich nur eines der beiden zum Weibchen. Krokodile und viele Schildkröten entwickeln das Geschlecht modifikatorisch bzw. phänotypisch oder epigenetisch abhängig von der Umgebungstemperatur im Brutgelege. Erreicht die ansteigende Bruttemperatur einen Schwellenwert, wechselt das Geschlecht von weiblich zu männlich.[2]
Regeneration
Regeneration ist die Fähigkeit eines Individuums, verloren gegangene Körperteile zeitlebens wieder zu erzeugen. Hohes Regenerationsvermögen haben Schwämme, Süßwasserpolypen (Hydra) und Strudelwürmer. Hydra kann Kopf und Fuß vollständig ersetzen, abhängig davon wo man sie durchtrennt. Ferner kann sie auch Nervenzellen ersetzen. Unter den Amphibien können die Schwanzlurche (Urodela) Extremitäten regenerieren, solange noch Häutungen bevorstehen. Wirbeltiere können neben der einmaligen Regeneration des Milchgebisses bei Säugern nur quer gestreifte syncytiale Muskelzellen wieder in einkernige Zellen zerlegen, die dann den Charakter von multipotenten Stammzellen für das Muskelgewebe haben. Die beiden Probleme bei der Regeneration sind das Erkennen von Art und Umfang des fehlenden Materials und die Frage, woher das Material für den Ersatz herkommt. Die erste Frage ist wenig geklärt, bei der zweiten gibt es entweder die Verwendung multipotenter Stammzellen oder die Transdifferenzierung von Zellen, bei der diese reembryonalisiert, also in einen früheren Zustand dedifferenziert werden. Die nur bedingte Regenerationsfähigkeit der meisten Arten wird als Hauptursache für den zwingenden Tod gesehen. Unter den Tieren gilt einzig die Hydra durch ihre hohe Regenerationsfähigkeit als potenziell unsterblich.[2]
Kontrolle des Zellwachstums und Krebs
Die Zellproliferation unterliegt während der gesamten Entwicklung strenger Wachstumskontrolle der Zellenzahl. Hierfür existieren Hemmfaktoren in Zellen, die im zwischenzellulären Diffusionsraum (Interstitium) diffundieren. Differenzierungsfaktoren, etwa in vermehrungsfähigen Zellen (-blasten), können in niedriger Konzentration das Zellenwachstum fördern, in hoher Konzentration dagegen die terminale Differenzierung auslösen. Wachstumshemmende Substanzen kommen als Zelladhäsionsmoleküle und als Bestandteile der extrazellulären Matrix vielfach auch in nicht diffundierender Form vor.[2]
Der gemeinsame Nenner unterschiedlicher Arten von Krebs ist das übermäßige, unkontrollierte Wachstum bestimmter Zelltypen. Krebs setzt die zellimmanente oder soziale Proliferations- bzw. Differenzierungskontrolle außer Kraft. Es kommt entweder zu einer zu schnellen Vermehrung von Vorläuferzellen, ohne dass ausreichend viele differenzierte Zellen zustande kommen, oder es kommt ohne Beschleunigung des Zellzyklus zu gestörtem Wachstum. Im häufigeren zweiten Fall bleibt entweder bei der Zellteilung der Stammzellencharakter beider Tochterzellen erhalten, die Teilungsaktivität wird fortgesetzt, obwohl das Differenzierungsprogramm abgeschlossen ist, oder Zellen werden nicht oder nicht rechtzeitig durch Apoptose eliminiert.[2]
Postnatale Wirbeltierentwicklung
Metamorphose
Metamorphose bedeutet die Aufgabe eines ersten larvalen Phänotyps und die gleichzeitige Entwicklung eines neuen Phänotyps. Dieser besetzt eine neue ökologische Nische und besiedelt einen neuen Lebensraum. Metamorphose bedingt die Umgestaltung auf jedem organismischen Niveau. Das reicht von der äußeren Morphologie über die Physiologie bis zu neuen Enzymausstattungen der Zellen. Alle Erscheinungsformen eines Organismus, der ein oder mehrere Metamorphosen vom Embryonalstadium, über die Larve, Puppe bis zum Imago durchläuft, lassen sich vom selben Genom ableiten. Aus der Reihenfolge der Basenpaare der DNA ist mit dem heutigen Wissenstand nicht ablesbar, in welcher Erscheinungsform die Phänotypen auftreten.[2]
Zur Entwicklung der Taufliege
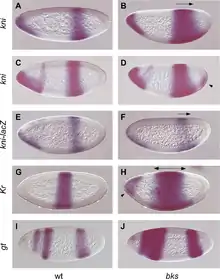
D. melanogaster ist einer der ältesten, am besten untersuchten Modellorganismen in der Biologie. Bereits zu Beginn des 20. Jahrhunderts konnten an D. melanogaster Gene auf bestimmten Chromosomen sowie Genabstände auf diesen lokalisiert werden, ohne dass damals molekularbiologische genetische Kenntnisse vorlagen. An der Taufliege konnten unter anderem grundlegende Kenntnisse zur Achsenbildung und Körpersegmentierung gewonnen werden. Für beide Vorgänge sind Morphogene ausschlaggebend, diffundierende Substanzen, die unterschiedlich starke und gerichtete Konzentrationsgradienten im syncytialen Embryo ausbilden. Diese Gradienten sind bereits maternal in der Oocyte vor der Befruchtung wirksam. Sie beeinflussen die Transkription anderer Gene. Es kommt im Zuge einer hierarchischen Kaskade von Genaktivierungen[3] zu zunehmend komplexer werdenden molekularen Vormustern, periodischen Pattern, die zu synchronen Zonenbildungen und damit zur Segmentierung des Larvenkörpers münden. Die maßgeblichen Genklassen von Segmentierungsgenen hierfür sind Maternalgene, Lückengene, Paarregel-Gene und Segmentpolaritätsgene.[9]
Das Studium der Entwicklung von D. melanogaster führte neben vielen anderen Erkenntnissen vor allem zur Entdeckung des Hox-Gen-Komplexes. Mutationen in der Expressionsabfolge homöotischer Gene, also auch von Hox-Genen, sogenannte homöotische Mutationen, führten bei Drosophila zur Entdeckung der Antennapedia-Mutante, einem Tier, dem Beine anstelle der Antennen aus dem Kopf wachsen. Auch der Hedgehog-Signalweg sowie das Masterkontrollgen Pax-6, das die frühe Augenentwicklung bei Wirbeltieren und Wirbellosen maßgeblich mit initiiert, wurden bei D. melanogaster erstmals beschrieben.
Die Gesamtentwicklung von D. melanogaster ist im Gegensatz zum Fadenwurm in hohem Maß eine regulative Entwicklung, da Programmierung der verschiedenen Entwicklungswege auf einer wechselseitigen Absprache zwischen Zellen, d. h. auf Zellinteraktionen, beruht. Die Entwicklung einzelner Zelltypen ist abhängig von ihrer Nachbarschaft.[2]
Zur Entwicklung des Fadenwurms
Der Fadenwurm Caenorhabditis elegans schuf als Modellorganismus der Wirbellosen Einblicke in grundlegende Entwicklungsvorgänge. Die Erkenntnisse gelten zum Teil auch bei Wirbeltieren. Seine Entwicklung ist im Gegensatz zur Wirbeltier-Entwicklung als streng deterministisch charakterisiert durch Zellkonstanz (Eutelie). Danach hat jedes adulte Hermaphrodit-Individuum exakt dieselbe Anzahl von 959 Zellen. Die Zellteilungen sind im frühen Stadium asymmetrisch; alle Teilungen bis zum adulten Tier sind einzeln analysiert und erlauben es, einen Zellstammbaum für alle adulten Zellen herzustellen und deren jeweiligen Entwicklungsweg exakt zu bestimmen. An C. elegans wurde der Mechanismus der Apoptose entdeckt, der dazu führt, dass exakt 131 während der Entwicklung erforderliche Zellen wieder gezielt abgebaut werden, ohne dass es dabei zu Schädigung von Nachbargeweben kommt. Asymmetrische Zellteilung, die zu einer größeren und einer kleineren Tochterzelle führt, ist verantwortlich für die frühe Ausbildung der antero-posterioren Körperachse. Direkte Zell-Zellkontakte sind dagegen maßgebend für die dorso-ventrale Achsenbildung. Wichtiger Signalweg ist bei letzterer der Notch-Signalweg. Im weiteren frühen Entwicklungsverlauf spielt auch der Wnt-Signalweg eine wichtige Rolle.[4] Demgegenüber spielen Signalwege, die Diffusion erzeugen und als Morphogen einen Gradienten ausbilden, bei diesem Tier keine Rolle. Die Gesamtentwicklung des Fadenwurms ist eine Mosaikentwicklung, da das Mosaik der Determinanten im Ei zu einer frühen Aufgabenzuweisung führt und den Zellen erlaubt, unabhängig von anderen ihren Entwicklungsweg zu gehen.[2]
Entwicklung der Pflanzen

Entwicklung wichtiger Modellorganismen der Pflanzen
Bei Pflanzen sind das Kleine Blasenmützenmoos (Physcomitrella patens), die Ackerschmalwand (Arabidopsis thaliana), der Mais (Zea mays subsp. mays), das Große Löwenmaul (Antirrhinum majus) und die Garten-Petunie (Petunia hybrida) wichtige Modellorganismen.
Wachstum und Entwicklung der Pflanzen
Methoden der Entwicklungsbiologie
Die Entwicklungsbiologie bedient sich verschiedener Methoden, die meist auch aus der Genetik bekannt sind. Die wichtigsten sind:
Studium der Entwicklungsbiologie an deutschsprachigen Universitäten
Die meisten Universitäten unterhalten einen Lehrstuhl für molekulare Entwicklungsbiologie, einige haben dedizierte Institute für Embryologie, so etwa die Universitäten Freiburg,[10] Göttingen,[11] oder die LMU-München,[12] oder auch für Pflanzenembryologie, etwa an der Universität Mainz.[13] Dazu heißt es an der LMU München: "Embryologie befasst sich mit allen Entwicklungsvorgängen von der Gametogenese über die Embryogenese und Fetogenese bis zur Geburt, sowie den morphologischen Aspekten des Geschlechtszyklus und der Trächtigkeit. Die Embryologie vermittelt damit wesentliche Grundlagen für das wichtige Gebiet der Reproduktionsbiologie."[14] Die Universität Wien bietet etwa die Vorlesungen Einführung in die Entwicklung und Vergleichende Entwicklungsbiologie der Wirbeltiere im Biologie-Bachelor-Studiengang. Einen Masterstudiengang Genetik und Entwicklungsbiologie bieten neben anderen Hochschulen etwa die Universitäten Wien[15] oder Freiburg.[16]
Siehe auch
Literatur
- Lewis Wolpert: Principles of Development: Das Original mit Übersetzungshilfen. (= Easy Reading Information Series). 3. Auflage. Spektrum Akademischer Verlag, 2007, ISBN 978-3-8274-1856-2.
- Lewis Wolpert: Principles of Development. 4. Auflage. Oxford Univ. Press, 2011, ISBN 978-0-19-955428-7. (englisch)
- Werner A. Müller, Monika Hassel: Entwicklungsbiologie und Reproduktionsbiologie von Mensch und Tieren. 5., vollständig überarbeitete Auflage. Springer-Verlag, Berlin/ Heidelberg/ New York 2012, ISBN 978-3-642-28382-6.
- Christiane Nüsslein-Volhard: Das Werden des Lebens. Wie Gene die Entwicklung steuern. dtv, München 2006, ISBN 3-423-34320-6.
- Scott F. Gilbert: Developmental Biology. 10. Auflage. Sinauer Associates, Sunderland 2013, ISBN 978-0-87893-978-7.
- Michael Kühl, Susanne Gessert: Entwicklungsbiologie. 1. Auflage. UTB basics Taschenbuch, 2010, ISBN 978-3-8252-3331-0.
- Shirley J. Wright: A Photographic Atlas of Developmental Biology. (Lose Blattsammlung). 2005, ISBN 0-89582-629-1.
Weblinks
- Max-Planck-Institut für Entwicklungsbiologie
- Gesellschaft für Entwicklungsbiologie (GfE)
- Entwicklungsbiologie und Bioinformatik (englisch)
- Zusammenstellung einiger online-Ressourcen zur Entwicklungsbiologie
- Entwicklungsbiologie übersichtlich für Schüler und Studenten
- UNSW Embryology
Videos:
- Morphogenesis and Embryogenesis
- Hühnchenentwicklung (Video)
- Froschentwicklung (Video)
- Zebrafisch Somitenentwicklung (Video)
- Eric Wieschaus: Patterning Development in the Embryo (Video)
- Viele Videos und Animationen in YouTube zu Furchung (cleavage) und Gastrulation (gastrulation).
Einzelnachweise
- Ernst Haeckel: Generelle Morphologie der Organismen. Berlin 1866.
- Werner A. Müller, Monika Hassel: Entwicklungsbiologie und Reproduktionsbiologie von Mensch und Tieren. 3., vollst. überarb. Auflage. Springer, 2003.
- Neil A. Campbell, Jane B. Reece: Biologie. 6., überarb. Auflage. Pearson Studium, 2006.
- Michael Kühl, Susanne Gessert: Entwicklungsbiologie. UTB basics, 2010.
- Alan Turing: The chemical basis of morphogenesis. (PDF; 1,2 MB). In: Phil. Trans. R. Soc. London B 237, 1952, S. 37–72. (Originalartikel)
- H. Meinhard, A. Gierer: Application of a theory of biological Pattern Formation based on lateral inhibition. In: J. Cell Sci. 15, 1974, S. 321–346.
- Mark C. Kirschner, John C. Gerhart: Die Lösung von Darwins Dilemma – Wie Evolution komplexes Leben schafft. Rowohlt, 2007, ISBN 978-3-499-62237-3. (Orig.: The Plausibility of Life, 2005)
- UNSW Embryology: Mouse Timeline Detailed
- Christiane Nüsslein-Volhard: Das Werden des Lebens. Wie Gene die Entwicklung steuern. München 2006.
- Universität Freiburg: Molekulare Embryologie (Memento des Originals vom 3. November 2014 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- Universität Göttingen: Embryologie
- Lehrstuhl für Anatomie, Histologie und Embryologie (Memento vom 26. Juni 2015 im Internet Archive)
- Universität Mainz: Pflanzenembryrologie
- Embryologie LMU München
- Universität Wien: Masterstudium: Genetik und Entwicklungsbiologie (Memento vom 3. November 2014 im Internet Archive)
- Uni Freiburg: Masterstudium: Genetik und Entwicklungsbiologie