Energiebereitstellung
Unter Energiebereitstellung (auch Energiestoffwechsel oder Metabolismus)[1] versteht man in der Physiologie die Mobilisierung, den Transport sowie den Abbau energiereicher Substrate unter ATP-Gewinnung (Resynthese) in den Muskelzellen der Tiere. Sie dient damit der Ausführung von Muskelarbeit. Es werden verschiedene Arten der Energiebereitstellung unterschieden, differenziert nach Energieträger (Kreatinphosphat, Kohlenhydrate, Fette oder Eiweiße) und Stoffwechselweg (aerob (oxidativ), anaerob lactacid (mit Lactat) oder anaerob alactacid (ohne Lactat)). Als aerob werden die in Verbindung mit Sauerstoff ablaufenden Stoffwechselprozesse bezeichnet, die in den Mitochondrien ablaufen. Der anaerobe Stoffwechsel läuft hingegen außerhalb der Mitochondrien im Zytoplasma ab. Wenn dieser mit einer vermehrten Lactatproduktion erfolgt, spricht man von einer lactaciden Energiebereitstellung, ansonsten von alactacidem Stoffwechsel.[2]
Einführung
Damit ein Muskel Kontraktionsarbeit verrichten kann, braucht er Energie, welche aus exothermen chemischen Reaktionen stammt. Dabei wird chemische in mechanische Energie umgewandelt. Der Energiebedarf des Organismus steigt somit bei körperlicher Arbeit an. Die für die Muskelkontraktion benötigte Energie wird zum größten Teil durch Hydrolyse (Wasseranlagerung) von Adenosintriphosphat (ATP) in Adenosindiphosphat (ADP) und Phosphat (Pi) zur Verfügung gestellt. Das ATP ist somit der direkte Energielieferant der Muskulatur. Da jedoch sein Vorrat sehr begrenzt ist, muss die Muskulatur zum Beispiel während sportlicher Aktivität ATP (wieder)herstellen, um die Aktivität aufrechterhalten zu können. Die für den Wiederaufbau (Resynthese) von ATP benötigte Energie wird wiederum durch schrittweise Oxidation der Nährstoffe Zucker (Kohlenhydrate), Fette beziehungsweise Fettsäuren und Eiweiße (Aminosäuren) gewonnen.[2] Ihr stehen dabei drei grundsätzlich unterschiedliche Mechanismen zur Verfügung. Es gibt die anaerob-alactacide Energiebereitstellung, welche anaerob (ohne Zuhilfenahme von Sauerstoff) und alactacid, also ohne (nennenswerte) Milchsäureproduktion (Lactatproduktion), verläuft. Der zweite Resyntheseweg ist der anaerob-lactacide, welcher zwar auch anaerob verläuft, aber mit einer Lactatproduktion verbunden ist. Demgegenüber wird bei der aeroben Energiebereitstellung die Energie unter Sauerstoffverbrauch freigesetzt.
Geschichte
Schon 1841 beobachteten Berzelius und 1877 Du Bois-Reymond, dass ein enger Zusammenhang zwischen Muskelkontraktion und Stoffwechsel herrscht, und zeigten, dass mit der Kontraktionsarbeit der Muskelzelle eine Lactatbildung verbunden ist. Diese Beobachtungen brachten 1914 Parnas und Wagner dazu, die dem Abbau des Glykogendepots des Muskels zugrundeliegende Lactatbildung als unmittelbare Energiequelle der Kontraktionsarbeit zu sehen. Gestützt wurde diese Aussage durch die Tatsache, dass die Bildung des Lactats aus Glykogen mit einer Energiefreisetzung verbunden ist. Jedoch wurden durch die Versuche von Hoett und Marks 1926 die Abhängigkeit des Kontraktionsvorganges von der Lactatbildung in Frage gestellt und von Clark und Eggleton 1932 bewiesen, dass eine deutlich über die Lactatverwertung hinausgehende Lactatbildung erst bei einer längeren Muskelarbeit auftritt. (Da die Lactatverwertung lange Zeit nicht sonderlich beachtet wurde, ging man damals allerdings davon aus, dass erst nach längerer Muskelarbeit überhaupt Lactat gebildet wird).
Nachdem die Substanz Kreatinphosphat im Muskel entdeckt wurde, wurde ein Zusammenhang zwischen dem Kreatinphosphatstoffwechsel und den Kontraktionsvorgängen in der Muskelzelle geschlossen, da während des Kontraktionsvorganges ein Abfall und in der Erholungsphase ein Wiederanstieg des Kreatinphosphats zu beobachten war. Während diese und weitere Erkenntnisse schon Zweifel an der Abhängigkeit des Kontraktionsvorganges und der Lactatbildung aufkommen und eine wichtige Stellung des Kreatinphosphats erkennen ließen, wurde durch Lundgaard 1931 die Lactattheorie der Muskelkontraktion eindeutig widerlegt. Durch einen geeigneten Versuch unter Zuhilfenahme eines von ihm vergifteten Muskels mit einem besonderen Substrat, welches eine Lactatbildung verhindert, dieser aber dennoch kontraktionsfähig und die geleistete Arbeit proportional zum Abbau des Kreatinphosphats war, stellte er die Lactattheorie unwiderruflich ein. In der Lactatbildung wurde so also ein anaerober Resyntheseweg für das Kreatinphosphat gesehen. Des Weiteren wurde festgestellt, dass unter aeroben Bedingungen eine Lactatbildung ausbleibt und die Resynthese des Kreatinphosphats durch oxidative Reaktionen stattfindet.
Schließlich entdeckte Lohmann 1931 dann das Adenosintriphosphat (ATP). Durch diese Entdeckung wurde jetzt auch das Kreatinphosphat als unmittelbare Energiequelle der Muskelarbeit in Frage gestellt, da in der Folgezeit dem ATP eine sehr wichtige Bedeutung als Koenzym, Regulationsfaktor im Zellstoffwechsel, Energieüberträger und unmittelbare Energiequelle zugeschrieben wurde. Als herausgefunden wurde, welch hohen Energiegehalt das ATP tatsächlich besitzt, wurde es als unmittelbare Energiequelle der Muskelkontraktion erkannt und durch Lohmann die ihm zugrundeliegenden quantitativen Reaktionen formuliert:[3]
- Kreatinphosphat + ADP ↔ Kreatin + ATP (Kreatinkinase),
Energiespeicher
Während die energiereichen Phosphate ATP und Kreatinphosphat (KrP) innerhalb der Muskelzelle nutzbar sind, können Glykogen, Fette und Eiweiße auch aus anderen Depots genutzt werden. Die verschiedenen Energiespeicher unterscheiden sich deutlich in der verfügbaren Menge und in der maximal möglichen Energieflussrate.[2]
| Substrat | ungefähre Menge in Phosphatresten (-P) in mmol/kg Muskel | maximale Einsatzdauer (im Wettkampf) |
|---|---|---|
| ATP Adenosintriphosphat | 6 | 2 – 3 s (theoretisch, da sofortige Resynthese) |
| KrP Kreatinphosphat | 20 – 25 | 7 – 10 s (theoretisch, da sofortige Resynthese) |
| Phosphatspeicher insgesamt (Phosphagen) | 30 | 7 – 10 s (20 s) |
| Glucose | 270 | 45 – 90 s (anaerober Abbau) |
| Glykogen | 3000 | 45 – 90 min (aerober Abbau) |
| Fette (Triglyzeride) | 50.000 | mehrere Stunden |
| Eiweiße (Aminosäuren) | ? | nur bei extremer Ausdauerbelastung |
Adenosintriphosphat (ATP)
Der direkt verfügbare ATP-Speicher reicht unter starken muskulären Beanspruchungen nur aus, um für ungefähr eine bis zwei Sekunden, also ein bis drei Muskelkontraktionen,[2] Energie bereitzustellen. Selbst unter der Voraussetzung, dass ATP bis zu AMP gespalten wird, herrscht im ruhenden Muskel nur ein ATP-Vorrat von ca. 6 µMol/g = 6 mMol/kg.[4] Wenn man nun die Tatsache bedenkt, dass der Mensch täglich so viel ATP verbraucht, wie es seinem Körpergewicht entspricht, ist es also umso erstaunlicher, dass ATP, das doch als so wichtig für die Muskelkontraktion gilt und die einzige unmittelbare Energiequelle darstellt, nur so beschränkt in der Muskelzelle vorhanden ist.
Kreatinphosphat (KrP)
Da der im Muskel vorhandene ATP-Vorrat nur für eine bis drei Muskelkontraktionen ausreicht (ungefähr zwei Sekunden Belastungsdauer),[5] muss der Körper ständig um eine Resynthese des ATPs als lebensnotwendige Substanz bemüht sein. Hier kommt das Kreatinphosphat ins Spiel, welches eine energiereiche chemische Verbindung aus Kreatin (Kr) und einem Phosphatrest ist. Die vorliegende Bindung zwischen dem Phosphat und dem Kreatin hat ein dem ATP entsprechendes Energiepotential. Durch die schnell ablaufende Reaktion:
- ADP + Kreatinphosphat ↔ ATP + Kreatin
wird durch die Abspaltung des Phosphatrestes und dessen Übertragung auf ADP das ATP resynthetisiert.[6] Zudem ist Kreatinphosphat in etwa drei- bis viermal so großer Menge (20–30 µMol/g) gegenüber dem ATP in der Muskelzelle vorrätig.[7] Der Kreatinphosphatspeicher ist also von großer Bedeutung für die Leistungsfähigkeit der Skelettmuskulatur, da er bei starker Konzentrationsarbeit ungefähr zehn Sekunden (Untrainierte ca. 6 s, Hochtrainierte ca. 12–20 s)[8] lang in der Lage ist, die dafür notwendige Energie bereitzustellen. Außerdem ist es die Energiequelle, welche das ATP sofort resynthetisieren kann, bis dann zu einem späteren Zeitpunkt andere Reaktionswege aktiviert sind.
Im Kreatinphosphat wird des Weiteren eine entscheidende Rolle als Energiegefälle gesehen, wodurch hohe Substratdurchsätze ermöglicht werden. Es steht auch fest, dass der Kreatinphosphatgehalt von der Höhe und Dauer der geleisteten Arbeit abhängt. Kommt es zu extrem starken Belastungen, kann der Kreatinphosphatspeicher fast vollständig ausgeschöpft werden und nach Ende der Belastung schnell wieder aufgefüllt werden. Sollte es jedoch dazu kommen, dass die Nachlieferung der energiereichen Phosphate unterbleibt, kommt es zu einem Erlöschen der Kontraktionsfähigkeit des Muskels.[9]
Glucose
Beim gesunden Menschen enthält das Blut einen bestimmten Glucoseanteil innerhalb einer Bandbreite von Konzentrationen (siehe auch Blutzucker). Wird diese Energie umgesetzt, so kommt es zu einem fortwährenden Ersatz aus den beiden nächstgenannten Energieträgern.
Glykogen
Dies ist eine Form der Glucose, sozusagen die „lagerfähige Form“. Glykogen kann als Muskelglykogen in der Skelettmuskulatur (ca. 1,5 g Glykogen/100 g Muskelfeuchtgewebe) und in der Leber gespeichert werden. Leberglykogen (75–90 g) dient zur Konstanthaltung des Blutzuckerspiegels (80–100 mg%) und trägt somit zur Aufrechterhaltung der Funktionsfähigkeit des Zentralnervensystems (ZNS) bei. Da das ZNS auf eine ständige Glucosezufuhr aus dem Blut angewiesen ist und selbst über geringe Glykogenvorräte verfügt, sichern bis zu 60 % der von der Leber an das Blut abgegebenen Glucose den Gehirnstoffwechsel. Bei lange andauernden submaximalen Belastungen (Langzeitausdauer) spielt die Glucoseaufnahme des Muskels aus dem durchströmenden Blut und somit dem Leberglykogen eine bedeutende Rolle. Studien von Coggan (1990) zeigen, dass nach einer 90-minütigen Belastung mit ca. 60 % der VO2max die Oxidation der Plasmaglucose etwa ein Drittel der gesamten Kohlenhydratoxidation beträgt.[10]
Bei starker Entleerung der Glykogendepots in der Leber tritt ein Abfall des Blutzuckerspiegels auf und kann bei weniger als 70 mg% bereits koordinative Störungen bewirken. Im Normalfall wird einer zu intensiven Glucoseverstoffwechselung zuungunsten des Gehirnstoffwechsels jedoch durch Schutzmechanismen vorgebeugt. So sinkt die Plasmainsulinkonzentration, die die Durchlässigkeit der Glucose durch die Zellmembran regelt, bei abnehmenden Glykogenvorräten durch lang andauernde muskuläre Arbeit auf bis zu 50 % des Ruheausgangswertes ab. Zusätzlich kann die Leber bei lange andauernden Belastungen teilweise Glucose aus Substraten wie Alanin und Glycerin neu herstellen (Gluconeogenese).[10]
Bei intensiver Dauerleistung (Wettkampf) reichen die Glykogenreserven des Körpers etwa 60 min bis 90 min zur Aufrechterhaltung des Glucose-Nachschubs.
Fette
Körperfett liegt im Unterhautfettgewebe (Hautdepot) und in der Muskelzelle in Form von Triglyceriden vor. Triglyceride bestehen aus drei an Glycerin gebundenen Fettsäuren. Die freien Fettsäuren (FFS) können in fast allen Organen oxidiert werden. In der Muskelzelle wird es in sogenannte „C2-Körper“ Acetyl-CoA umgewandelt und in den Citratzyklus eingeschleust. Allerdings ist die chemische Reaktion sehr langsam, so dass diese Form der Energiebereitstellung mit wachsender Belastung einen abnehmenden relativen Anteil der bereitgestellten Energie liefert. Bei weiter wachsender Intensität nimmt auch ihr absoluter Anteil ab. Der intramuskuläre Triglyceridgehalt beträgt 0,3 – 0,8 Vol.-%. Die freien Fettsäuren werden unter Wasseraufnahme (Hydrolyse) aus den Triglyceriden freigesetzt. Die Lipolyse (Triglyceridspaltung) wird durch die belastungsbedingte Freisetzung der Katecholamine Adrenalin und Noradrenalin und bei länger andauernder Belastung vorrangig durch das Wachstumshormon Somatropin stimuliert. Gehemmt wird sie durch die Blutlactatkonzentration. So führen Blutlactatwerte von 5 – 8 mmol/l zu einer signifikanten Senkung des Plasmaspiegels der Fettsäuren.[11]
Die Nutzung der Fettoxidation ist von verschiedenen Faktoren wie Belastungsdauer, Belastungsintensität und intramuskulärem Glykogenangebot abhängig. Die Fettdepots des Unterhautfettgewebes werden hauptsächlich bei lange andauernden Belastungen geringer und mittlerer Intensität und bereits reduzierten Glykogenreserven genutzt, die Mobilisation setzt erst nach einer 15 bis 30-minütigen Belastungszeit ein. Der Ausdauertrainingszustand spielt hier eine große Rolle, da mit zunehmendem Leistungsniveau der Prozentsatz der Fettsäureverbrennung an der Energiebereitstellung ansteigt und somit Kohlenhydratdepots geschont werden.[11]
Blutfette sind eine Zwischenform als Energieträger. Neben der Verstoffwechselung von Zucker sind die Muskelzellen auch in der Lage, Energie direkt aus Fett zu mobilisieren.
Eiweiße
→ Siehe auch Aminosäure-Stoffwechsel
Da Eiweiße (Proteine) im Normalfall im Baustoffwechsel zu Aminosäuren abgebaut werden, werden sie nur bei lange andauernden Belastungen (ab 90 Minuten) und reduziertem intramuskulären Glykogenspeicher verstoffwechselt und können dann 5 – 15 % am Gesamtenergiestoffwechsel abdecken. Eine Oxidation von Aminosäuren lässt sich durch die Zunahme von Harnstoff im Blutserum, zum Beispiel nach einer Langzeitausdauerbelastung nachweisen. Dabei sinkt auch die Konzentration von Leucin, Isoleucin und Valin im Blut, was auf eine Beteiligung am oxidativen Stoffwechsel in der Skelettmuskulatur hinweist. In Extremsituationen wie zum Beispiel bei einem Aufenthalt in großen Höhen ist der Organismus auch in der Lage, indirekt Muskelprotein abzubauen, sich also quasi selbst zu verstoffwechseln.[12]
Energiestoffwechsel
→ Siehe auch Energiestoffwechsel
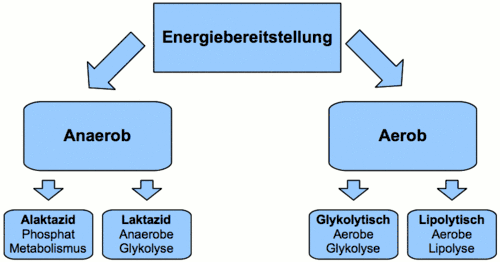
Die für die Resynthese von ATP benötigte Energie kann auf unterschiedliche Weise mobilisiert werden. Es werden vier Arten der Energiebereitstellung unterschieden, differenziert nach Energieträger und Stoffwechselweg. Auf der Grundlage der Energiebereitstellungsformen, die im Wettkampf in einer bestimmten anteilsmäßigen und zeitlichen Struktur auftreten, erfolgt in der sportwissenschaftlich fundierten Trainingslehre die leistungsstrukturelle Ableitung der Trainingsbereiche.
Abbau von ATP setzt Energie frei
Die Grundlage für jede Muskelkontraktion ist der Abbau von Adenosintriphosphat zu Adenosindiphosphat (ADP) und Phosphat (P).[13] Das ATP ist eine hochenergetische Verbindung, bestehend aus Adenin mit Ribose und drei Phosphaten. Es stellt die einzige Energiequelle dar, welche die Zelle direkt benutzen kann. Das so wichtige ATP ermöglicht nicht nur mechanische Arbeit, sondern auch sehr wichtige energieerfordernde Transformationen, die Aktivierung der freien Fettsäuren und die Erhaltung der labilen Proteinstrukturen.[3] Die für die Muskelkontraktion wichtige Reaktion des ATP an der Myosin-ATPase lautet:
- ATP → ADP + P (+ Energie).
In einer anderen (nicht typischerweise Myosin-ATPase assoziierten) chemischen Reaktion kann ATP bis zu AMP (Adenosinmonophosphat) abgebaut werden:
- ATP → AMP + PP (+ Energie).
Letztere Reaktion stellt aber eine untergeordnete Rolle in der Energiegewinnung dar. Die Spannungsentwicklung des Muskels ist stark abhängig vom vorliegenden ATP-Gehalt. Erniedrigungen dieses Gehaltes führen (ab einem kritischen Schwellenwert) zunächst zu einer Einschränkung der Spannungsentwicklung und schließlich bis zur Kontraktionsunfähigkeit bei Erregungsreizen. Somit gehen die Veränderungen des ATP-Gehalts mit Veränderungen der potenziellen Arbeitsleistung einer Muskelzelle einher. Sich verkürzende Muskeln, welche jedoch keine Arbeit leisten, zeigen keinen, beziehungsweise nur einen unwesentlichen ATP-Abfall. Arbeit verrichtende Muskeln, welche auch unter Belastung stehen, zeigen einen in Abhängigkeit dieser Arbeitsleistung erniedrigten ATP-Spiegel und eine dementsprechende Wärmebildung. Die Wärmeentwicklung bei der Muskelarbeit geht also mit einer Veränderung des ATP-Spiegels einher und lässt sich als Folge der Entropie erklären.[14]
ADP
Durch den Abbau des durch die Myosin-ATPase entstandenen ADP kann durch eine geeignete Reaktion, die Myokinase (2 mol ADP → 1 mol ATP + 1 mol AMP), unter extremen Notfallbedingungen ATP gewonnen werden (diese Reaktion spielt für die Energiebereitstellung der Muskulatur im Regelfall aber keine relevante Rolle).[15] Dadurch wäre das wichtige ATP aber unmittelbar für weitere Kontraktionsarbeit bereitstellbar. Folgernd durch die aus der Myosin-ATPase hohen ADP-Konzentrationen und der Sensitivität der Myokinase auf hohe ADP-Spiegel – d. h. ihre Aktivität wird durch einen hohen ADP-Spiegel gesteigert – gilt die ADP-Konzentration als Regelgröße für die Bereitstellung von ATP aus ADP. Somit ist das scheinbare Endprodukt ADP keine zu vernachlässigende Größe, denn seine energiereiche Phosphatbindung kann zumindest theoretisch noch genutzt werden.
Anaerob alactacid (Phosphatmetabolismus)
Bei der anaerob-alactaciden Energiebereitstellung wird kein Sauerstoff benötigt und es entsteht keine Milchsäure. Sie spielt in den ersten Sekunden einer sportlichen Belastung die entscheidende Rolle und reicht nur für einige Sekunden bzw. wenige maximale Muskelkontraktionen (z. B. Kurzsprints, Antritte, manche Formen des Krafttrainings), weil das als Energieträger dienende Kreatinphosphat nur in geringer Menge in den Muskelzellen vorhanden ist. Die ATP-Bildungsrate (genauer: die Resynthese von ATP aus ADP und dem Energieträger pro Zeiteinheit) ist beim anaerob-alactaciden Stoffwechsel am höchsten. Nachdem der vorab vorhandene ATP-Vorrat der Muskulatur bereits nach wenigen Sekunden aufgebraucht ist, erfolgt die weitere ATP-Resynthese in den folgenden zehn bis 30 Sekunden mittels des ebenfalls schnell verfügbaren Kreatinphosphates.
Die energieliefernden anaerob-alactaciden Reaktionen:
- ATP + H2O → ADP + P + Energie
- ADP + Kreatinphosphat ↔ ATP + Kreatin
Anaerob lactacid
→ Siehe auch Milchsäuregärung
Der anaerob-lactacide Stoffwechsel benötigt keinen Sauerstoff, führt aber zur Bildung von Milchsäure (Lactat). Er läuft sehr schnell ab (etwa die halbe ATP-Bildungsrate des anaerob-alactaziden Stoffwechsels) und reicht bei annähernder Höchstbelastung (95 %) etwa 20 – 40 Sekunden. Energieträger ist die im Zytosol vorhandene Glucose, aus der über die Glykolyse ATP gewonnen wird. Nachteil: Bei Steigerung der Leistung bzw. Aufrechterhaltung einer hohen Leistungsabforderung können die aeroben Mechanismen des Abbaus und der Verwertung des gebildeten Lactats einen starken Anstieg der Lactatkonzentration nicht verhindern. Es kommt schließlich zu einem sprunghaften Lactatanstieg, die Leistungserbringung muss abgebrochen oder die Intensität stark reduziert werden.
Beim anaerob lactaciden Energiestoffwechsel wird durch den Abbau von Traubenzucker (Glucose) oder Glykogen (eine Speicherform der Glucose) über chemische Reaktionen Lactat und ATP hergestellt:
- Glykogen ↔ 2ATP + Milchsäure.
Diese Reaktion wird also als Glykolyse mit anschließender Milchsäuregärung bezeichnet, welche im Sarkoplasma stattfindet (vgl. Weineck 2006, S. 101). Als Energielieferant wird Glucose (insbesondere auch aus Glykogen) herangezogen. Das intrazelluläre Glykogen ist energetisch vorteilhafter, da es nicht erst über die Blutwege herbeigeführt werden muss. Der Abbau von 1 Mol Glucose zu Lactat bringt 2 Mol ATP. Wird Glykogen verwertet, bringt das rein rechnerisch 3 Mol ATP.[16] Das Zwischenprodukt Brenztraubensäure (Pyruvat) wird während der Milchsäuregärung anaerob zu Lactat umgewandelt.
Das bei der Milchsäuregärung entstandene Lactat hat jedoch auf den gesamten Stoffwechsel, sowohl lokal als auch allgemein, Auswirkungen, da es über den Lactat-Shuttle-Mechanismus in andere Bereiche des Körpers transportiert wird. Nach maximalen Belastungen sind im Muskel Lactatwerte von bis zu 25 mMol/kg, im Blut bis zu 20 mMol/kg[17] zu finden. Damit einher geht in der Regel eine extreme Übersäuerung im lokalen Gewebe sowie im arteriellen Blut, was mit einer Azidose (stark herabgesetzter pH-Wert) verbunden ist. Durch die Azidose kommt es zu einer Enzymhemmung, welche ein Erliegen der glykolytischen Stoffwechselprozesse herbeiführt. Dieser Abbruch der Maximalbelastung stellt eine wichtige Schutzfunktion für den Organismus dar. Sie verhindert eine zu starke Übersäuerung des Muskels, welche eine Zerstörung intrazellulären Eiweißstrukturen zur Folge hätte.[17]
Aerob glykolytisch
Der aerob glykolytische Stoffwechsel nutzt unter Einsatz von Sauerstoff Kohlenhydrate. Er spielt bei den Energiebereitstellungen für alle Belastungen, deren Dauer über eine Minute hinausgeht, eine Rolle.[18] Die Energie wird nach der vereinfachten Formel Traubenzucker + Sauerstoff → Wasser + Kohlendioxid + Energie gewonnen. Dieser Weg hat folgende Merkmale: Er ist schneller als Fettstoffwechsel (ATP-Bildungsrate etwa ein Viertel des anaerob-alactaciden Stoffwechsels), das Glykogen (die spezifische Form des Traubenzuckers) ist im Muskel gespeichert, muss nicht erst antransportiert werden und Glucose kann durch kohlenhydrathaltige Getränke nachgeführt werden. Er nutzt die bei der Weiterverstoffwechselung der energiereichen Zwischenprodukte freiwerdende Energie. Hierbei handelt es sich in erster Linie um das beim anaerob-lactaciden Stoffwechsel angefallene Lactat und das Pyruvat. Die Teilprozesse oxidative Decarboxylierung, Citratzyklus und Atmungskette finden im Gegensatz zu den anaeroben Stoffwechselwegen nicht im Cytoplasma statt, sondern in den Mitochondrien. Der aerobe Kohlenhydratstoffwechsel hat bei mittlerer und submaximaler Intensität den größten Anteil an der Muskelarbeit. Nachteil: Die körpereigenen Glykogen-Reserven sind auf etwa 60 bis 90 min Dauerbelastung begrenzt, bei stundenlanger Muskelarbeit begrenzt die Aufnahmefähigkeit des Darms für Kohlenhydrate die Intensität der Leistung.
Die aktivierte Essigsäure (Acetyl-CoA), welche durch oxidative Decarboxylierung entstanden ist, durchläuft für den weiteren Abbau den Citronensäurezyklus und die Atmungskette. Bei dieser Art von Energiebereitstellung werden aus 1 Mol Glucose etwa 32 Mol ATP gewonnen. Wird das intrazelluläre Glykogen zum Abbau verwendet, entstehen sogar 34 Mol ATP:
- 1 Glucose + 6 O2 + 32 ADP + 32 P → 6 CO2 + 6 H2O + 32 ATP
Beim aeroben Abbau von Glucose kann etwa 15 mal so viel ATP gewonnen werden wie bei der Milchsäuregärung.[19] Diese hohe Energieausbeute hat jedoch auch einen entscheidenden Nachteil. Mit Hilfe der oxidativen Verbrennung werden zwar viele Mol ATP bereitgestellt, doch erfolgt diese Energiebereitstellung über lange Reaktionsketten, weshalb es längere Zeit dauert, bis diese Energie zur Verfügung steht.
Aerob lipolytisch (Aerobe Lipolyse)
→ Siehe auch Fettverbrennung
Der aerob lipolytische Stoffwechsel nutzt unter Einsatz von Sauerstoff Fettsäuren. Die Energie wird nach der vereinfachten Formel Fett + Sauerstoff → Wasser + Kohlendioxid + Energie gewonnen (Lipolyse – 1 Mol FFS ergibt ca. 130 Mol ATP). Der Prozess findet ebenfalls zum wesentlichen Teil (ATP-Gewinnung) in den Mitochondrien statt (Betaoxidation) und hat bei niedriger Intensität den größten Anteil an der Muskelarbeit. Vorteil: sehr große körpereigene Reserven (ausreichend für tagelange Dauerbelastungen), Nachteil: sehr langsam (ATP-Bildungsrate nur etwa ein Zehntel des anaerob-alactaciden Stoffwechsels).
Diese Reaktion setzt bei Ausdauerbelastungen nach etwa 20 Minuten ein. Zudem kann in Notfällen noch Protein als Energielieferant dienen, wobei diese beiden Arten der Energiegewinnung (FFS und Eiweiß) insbesondere bei Ausdauerbelastungen (niedrige Belastungsintensität) relevant sind.
Kombination der Formen unter realen Belastungen
Aufgrund der Tatsache, dass die Kontraktionsgeschwindigkeit des Muskels bei den energiereichen Phosphaten am schnellsten und bei der oxidativen Energiebereitstellung am langsamsten ist, bedingt durch verschiedene Flussraten, ist bei verschiedenen Belastungsintensitäten mit verschiedener Belastungsdauer häufig eine Mischform der energieliefernden Systeme zu beobachten. Somit verändert sich die Intensität der Muskelarbeit, also die Kontraktionsgeschwindigkeit der Muskelfaser, in Abhängigkeit von der energetisch möglichen Flussrate.
Es zeigt sich demnach die große Nützlichkeit der unterschiedlichen Flussraten. Sollen zum Beispiel hohe Intensitäten erzielt werden (hohe Energieumsätze), dies ist vor allem bei Schnelligkeitsbelastungen der Fall, müssen höhere Flussraten erzielt werden. Folglich muss auf die anaerob-alactacide (ATP, PKr) als auch auf die lactacide Energiegewinnung zurückgegriffen werden. Sollen geringere Arbeitsintensitäten abgedeckt werden, wie zum Beispiel bei Langstreckenläufen, überwiegen zwangsläufig die aeroben Energiebereitstellungsprozesse.
Sauerstoffdefizit und EPOC
→ Siehe EPOC und Sauerstoffdefizit
Siehe auch
Weblinks
- Die muskuläre Energiebereitstellung im Sport – Dr. Kurt A. Moosburger, Sportarzt und Ernährungsmediziner aus Österreich (PDF; 189 kB)
- Energiebereitstellung – Dr. Peter Wastl, Institut für Sportwissenschaft, Heinrich-Heine-Universität Düsseldorf (PDF; 71 kB)
Literatur
- Bartl, Meinhard, Moisl: Abitur-Training Biologie. Biologie 1. Freising 1987.
- D. Cunningham, J. Faulkner: The effect of training on aerobic and anaerobic metabol-ism during short exhaustive run. In: Med. And Sci. in Sports. 2 (1969), S. 65–70.
- H. Heck, Trainerakademie Köln e.V. (Hrsg.): Energiestoffwechsel und medizinische Leistungsdiagnostik. Schorndorf 1990, ISBN 3-7780-8081-4.
- U. Helmich: Citratzyklus. u-helmich.de, 24. Januar 2009.
- H. Senger, R. Donath: Zur Regulation der oxydativen Substratverwertung im Muskel bei erhöhtem ATP-Umsatz. In: Medizin und Sport. 12 (1977), S. 391–400.
Einzelnachweise
- Fritz Zintl: Ausdauertraining. blv, München 2009, ISBN 978-3-8354-0555-4, S. 46.
- Fritz Zintl: Ausdauertraining. blv, München 2009, ISBN 978-3-8354-0555-4, S. 46–47.
- T. Hettinger, W. Hollmann: Sportmedizin, Grundlagen für Arbeit, Training und Präventivmedizin. 4. Auflage. Stuttgart 2000, ISBN 3-7945-1672-9, S. 62.
- J. Keul, E. Doll, D. Keppler: Muskelstoffwechsel. Die Energiebereitstellung im Skelettmuskel als Grundlage seiner Funktion. München 1969, DNB 457199261, S. 20.
- Andreas Hohmann, Martin Lames, Manfred Letzelter: Einführung in die Trainingswissenschaft. Limpert, Wiebelsheim 2007, ISBN 978-3-7853-1725-9, S. 52.
- J. Weineck: Sportbiologie. 4. Auflage. Balingen 1994, ISBN 3-929587-43-2, S. 101.
- J. Keul, E. Doll, D. Keppler: Muskelstoffwechsel. Die Energiebereitstellung im Skelettmuskel als Grundlage seiner Funktion. München 1969, DNB 457199261, S. 22.
- Andreas Hohmann, Martin Lames, Manfred Letzelter: Einführung in die Trainingswissenschaft. Limpert, Wiebelsheim 2007, ISBN 978-3-7853-1725-9, S. 53.
- J. Keul, E. Doll, D. Keppler: Muskelstoffwechsel. Die Energiebereitstellung im Skelettmuskel als Grundlage seiner Funktion. München 1969, DNB 457199261, S. 25.
- Fritz Zintl: Ausdauertraining. blv, München 2009, ISBN 978-3-8354-0555-4, S. 48.
- Fritz Zintl: Ausdauertraining. blv, München 2009, ISBN 978-3-8354-0555-4, S. 48–49.
- Fritz Zintl: Ausdauertraining. blv, München 2009, ISBN 978-3-8354-0555-4, S. 49.
- J. Weineck: Sportbiologie. 4. Auflage. Balingen 1994, ISBN 3-929587-43-2, S. 38.
- J. Keul, E. Doll, D. Keppler: Muskelstoffwechsel. Die Energiebereitstellung im Skelettmuskel als Grundlage seiner Funktion. München 1969, DNB 457199261, S. 19.
- J. Koolman u. a.: Taschenatlas der Biochemie. Georg Thieme Verlag, 2002, ISBN 3-13-759403-0, S. 336 (books.google.de).
- J. Keul, E. Doll, D. Keppler: Muskelstoffwechsel. Die Energiebereitstellung im Skelettmuskel als Grundlage seiner Funktion. München 1969, DNB 457199261, S. 32.
- J. Weineck: Sportbiologie. 4. Auflage. Balingen 1994, ISBN 3-929587-43-2, S. 39.
- J. Keul, E. Doll, D. Keppler: Muskelstoffwechsel. Die Energiebereitstellung im Skelettmuskel als Grundlage seiner Funktion. München 1969, DNB 457199261, S. 38.
- J. Weineck, A. Weineck: Leistungskurs Sport. 3. Auflage. Band 1. Forchheim 2004, ISBN 3-00-013707-6, S. 104.