Haplogruppe R1b (Y-DNA)
R1b ist eine Haplogruppe des Y-Chromosoms und wird zur Bestimmung von genetischen Verwandtschaftsverhältnissen verschiedener Bevölkerungsgruppen angewandt. Sie ist eine Untergruppe von R. R1b wird definiert durch die Mutation M343 eines Einzelnukleotid-Polymorphismus (SNP), die im Jahr 2004 entdeckt wurde.[3] Von 2001 bis 2005 wurde R1b durch den SNP P25 definiert. In anderen Systemen wird sie auch als Hg1 und Eu18 klassifiziert. Der aktuelle Stand kann über die Seiten der International Society of Genetic Genealogy (ISOGG) abgerufen werden.[4][5]
| Haplogruppe des Y-Chromosoms | |
|---|---|
| Name | R1b |
| Mögliche Ursprungszeit | vor weniger als 18.500 Jahren[1] |
| Möglicher Ursprungsort | Westasien, Zentralasien,[2] Südsibirien,[2] Osteuropäische Ebene[2] |
| Vorgänger | R1 |
| Mutationen | R = M343 |
Aufspaltung
Die Haplogruppe R1b stammt von R1 ab, wie auch R1a. R1b entstand nach derzeitiger Schätzung vor weniger als 18.500 Jahren[6] in Asien. Die Aufspaltung der Haplogruppe R in R1 und R2 ist durch einen Fund in Mal'ta am Baikalsee belegt.[7]
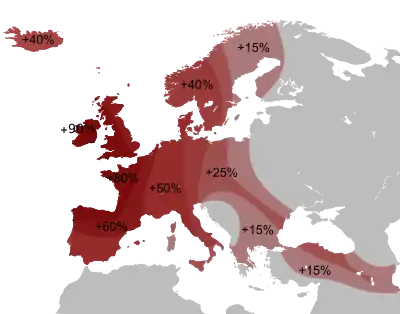
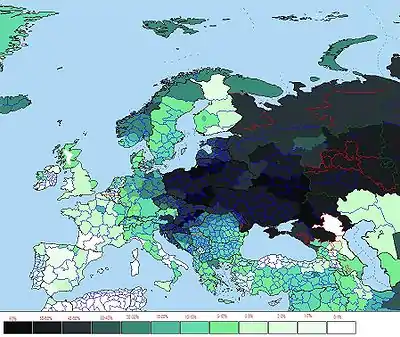
Die männliche Haplogruppe R (Y-DNA) soll auf sibirische Jäger-und-Sammler-Kulturen, die zwischen 18.000 und 25000 Jahren lebten, zurückgehen.[8] Sie ist die Basis der heute häufigsten Haplogruppen in Europa, wobei die R1b-Gruppe in Westeuropa und R1a-Gruppe in Osteuropa dominieren. So hat die R1b-Gruppe in Westeuropa einen mittleren Anteil von 70 %, mit Höchstwerten von über 80 % etwa in Irland, Schottland, Wales und auch Nordfrankreich. Eine Studie von Haak (2015)[9] ergab, dass R1b sowie R1a sich ab 3000 v. Chr. aus der Region der pontisch-kaspischen Steppen in Richtung Westen verbreitete. Vor dieser Zeit wiesen nur 1 von 70 westeuropäischen Individuen eine R1-Gruppe auf. Von den männlichen Proben aus der indoeuropäischen, in den kaukasischen Steppen heimischen Jamnaja-Kultur dagegen verfügen alle über R1b. Hypothetisch gelangten die Haplogruppen R1b und R1a in mehreren Migrationswellen der Jamnaja-Kultur nach Westeuropa. Genetisch wiesen die regionalen Jamnaja-Kulturen eine differente Verteilung der R1-Gruppen auf, die südliche Ethnien dieser Kultur, zeigen vorwiegend R1b, während die nördliche Wald-Steppen-Kultur, vorwiegend R1a-Merkmale aufweisen. Mit der ersten Migrationswelle gelangte R1b um 4200 v. Chr. in die Donau-Region und von dort aus in die Balkangebiete und auch nach Ungarn. Vermutet wird, dass die R1b-Migranten von einer Minderheit von R1a-Männern begleitet wurden. Die Glockenbecher-Kultur löste sich um 2200 v. Chr. auf und es folgte um 2300 v. Chr. die Aunjetitzer Kultur, die u. a. Ausgangspunkt der späteren germanischen, keltischen und italischen Kulturen war.[10]
Die Aufspaltung der R1* Haplogruppe fand wohl eher im zentralen bis westlichen Teil Asiens statt, wo sowohl R1a als auch R1b in relativer Nähe zueinander gefunden wurde.[11] R1b-U106 ist eine Schwestergruppe zur mittelneolithischen R1b-M269 und wird heute vorwiegend im germanischen Sprachraum, den nördlichen Alpen und entlang des Rheins gefunden, während R1b-P312 vorwiegend im romanischen Sprachraum gefunden wird. Die Aufspaltung der Haplogruppe R1b-M269 wird auf das mittlere Neolithikum vor etwa 5000 Jahren angesetzt. Unklar ist, ob diese Aufspaltung im Alpenraum stattfand oder es sich um zwei Ausbreitungswellen handelt, die jeweils in den nördlichen und südlichen Alpenraum expandierten.
Daraus entstanden in der Bronzezeit vor etwa 4176 ± 696 Jahren die Haplogruppen DF27 mit der größten Konzentration in Iberien (rund 44 %), R1b-U152, das vorwiegend im nördlichen Italien und im südlichen Alpengebiet konzentriert ist, und R1b-M529, das sich am stärksten in England und der Bretagne konzentriert. Die Entstehung aller drei Gruppen fällt damit in die Bronzezeit und deutet auf Kontakte mit Einwanderern.
Alle vier Haplogruppen sind Träger der Glockenbecherkulturen und werden im Zuge der Vermischung mit den Schnurkeramikern zu Trägern der Becherkulturen. Weitere Untergruppen wie Z196/Z195, geschätzt auf 3173 ± 502, sind deutlich mit der sogenannten Orientalisierung (d. h. orientalisch-griechischen Importfunden) der beginnenden Eisenzeit entlang der Mittelmeerküsten in Verbindung gebracht. Archäologisch sind Phönizier etwa 200 Jahre später in Portugal nachgewiesen. Diese Ergebnisse widersprechen den Schätzungen von yfull[12] deutlich. Kontrovers wird auch der erhöhte Anteil der baskischen Bevölkerung diskutiert, wobei angrenzende Regionen wie Aragon und Cantabrien ebenfalls erhöhte Werte eisenzeitlicher Schwesterlinien aufweisen. Jüngere Gruppen lassen sich erwartungsgemäß deutlich mit der spanischen Kolonialisierung Amerikas in Verbindung bringen.[13]
| M269 |
| ||||||||||||||||||||||||||||||||||||
Untergruppen
| M343 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Beispiel R1b1a2a1a1a (R-U106)
Diese Untergruppe ist definiert durch den SNP U106, auch bekannt als S21 und M405. Sie repräsentiert über 25 % der gesamten R1b-Population in Europa und findet sich vorwiegend entlang des Rheins, daher wird sie als „germanisch“[14] betrachtet. Die höchste Prävalenz (35%) wurde aus den Niederlanden berichtet.
| U106/S21 |
| |||||||||||||||||||||||||||||||||||||||
Daten nach Myres et al. (2010)[15]
| Population | Sample size | R-M269 | R-U106 | R-U106-1 |
|---|---|---|---|---|
| Austria | 22 | 27 % | 23 % | 0,0 % |
| Central/South America | 33 | 0,0 % | 0,0 % | 0,0 % |
| Czech Republic | 36 | 28 % | 14 % | 0,0 % |
| Denmark | 113 | 34 % | 17 % | 0,9 % |
| Eastern Europe | 44 | 5 % | 0,0 % | 0,0 % |
| England | 138 | 57 % | 20 % | 1,4 % |
| France | 56 | 52 % | 7 % | 0,0 % |
| Germany | 332 | 43 % | 19 % | 1,8 % |
| Ireland | 102 | 80 % | 6 % | 0,0 % |
| Italy | 284 | 37 % | 4 % | 0,0 % |
| Jordan | 76 | 0,0 % | 0,0 % | 0,0 % |
| Middle-East | 43 | 0,0 % | 0,0 % | 0,0 % |
| Netherlands | 94 | 54 % | 35 % | 2,1 % |
| Oceania | 43 | 0,0 % | 0,0 % | 0,0 % |
| Oman | 29 | 0,0 % | 0,0 % | 0,0 % |
| Pakistan | 177 | 3 % | 0,0 % | 0,0 % |
| Palestine | 47 | 0,0 % | 0,0 % | 0,0 % |
| Poland | 110 | 23 % | 8 % | 0,0 % |
| Russia | 56 | 21 % | 5,4 % | 1,8 % |
| Slovenia | 105 | 17 % | 4 % | 0,0 % |
| Switzerland | 90 | 58 % | 13 % | 0,0 % |
| Turkey | 523 | 14 % | 0,4 % | 0,0 % |
| Ukraine | 32 | 25 % | 9 % | 0,0 % |
| United States | 58 | 5 % | 5 % | 0,0 % |
| US (European) | 125 | 46 % | 15 % | 0,8 % |
| US (Afroamerican) | 118 | 14 % | 2,5 % | 0,8 % |
Beispiel R1b1a2a1a2 (R-P312/S116)
Neben R-U106, ist R-P312 die zweite große Gruppe in Europa die beide von R1b1a2 (R-M269) abstammen. Sie ist auch als S116 bekannt. Myres et al. beschreiben sie als sich vom westlichen Einzugsgebiet des Rheins her ausbreitend.[15] Allgemein betrachtet man sie als atlantische Gruppe die mit der keltischen und römischen Expansion in Zusammenhang gebracht wird.
| P312 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Country | Sampling | sample | R-M269 | Source |
|---|---|---|---|---|
| Wales | National | 65 | 92,3 % | Balaresque et al. (2009) |
| Spain | Basques | 116 | 87,1 % | Balaresque et al. (2009) |
| Ireland | National | 796 | 85,4 % | Moore et al. (2006) |
| Spain | Catalonia | 80 | 81,3 % | Balaresque et al. (2009) |
| France | Ille-et-Vilaine | 82 | 80,5 % | Balaresque et al. (2009) |
| France | Haute-Garonne | 57 | 78,9 % | Balaresque et al. (2009) |
| England | Cornwall | 64 | 78,1 % | Balaresque et al. (2009) |
| France | Loire-Atlantique | 48 | 77,1 % | Balaresque et al. (2009) |
| France | Finistère | 75 | 76,0 % | Balaresque et al. (2009) |
| France | Basques | 61 | 75,4 % | Balaresque et al. (2009) |
| Spain | East Andalucia | 95 | 72,0 % | Balaresque et al. (2009) |
| Spain | Castilla La Mancha | 63 | 72,0 % | Balaresque et al. (2009) |
| France | Vendée | 50 | 68,0 % | Balaresque et al. (2009) |
| France | Baie de Somme | 43 | 62,8 % | Balaresque et al. (2009) |
| England | Leicestershire | 43 | 62,0 % | Balaresque et al. (2009) |
| Italy | North-East (Ladin) | 79 | 60,8 % | Balaresque et al. (2009) |
| Spain | Galicia | 88 | 58,0 % | Balaresque et al. (2009) |
| Spain | West Andalucia | 72 | 55,0 % | Balaresque et al. (2009) |
| Portugal | South | 78 | 46,2 % | Balaresque et al. (2009) |
| Italy | North-West | 99 | 45,0 % | Balaresque et al. (2009) |
| Denmark | National | 56 | 42,9 % | Balaresque et al. (2009) |
| Netherlands | National | 84 | 42,0 % | Balaresque et al. (2009) |
| Italy | North East | 67 | 41,8 % | Battaglia et al. (2008) |
| Armenia | Ararat Valley | 41 | 37,3 % | Herrera et al. (2012) |
| Russia | Bashkirs | 471 | 34,40 % | Lobov (2009) |
| Germany | Bavaria | 80 | 32,3 % | Balaresque et al. (2009) |
| Armenia | Lake Van | 33 | 32,0 % | Herrera et al. (2012) |
| Armenia | Gardman | 30 | 31,3 % | Herrera et al. (2012) |
| Italy | West Sicily | 122 | 30,3 % | Di Gaetano et al. (2009) |
| Poland | National | 110 | 22,7 % | Myres et al. (2007) |
| Serbia | National | 100 | 10,0 % | Belaresque et al. (2009) |
| Slovenia | National | 75 | 21,3 % | Battaglia et al. (2008) |
| Slovenia | National | 70 | 20,6 % | Balaresque et al. (2009) |
| Turkey | Central | 152 | 19,1 % | Cinnioğlu et al. (2004) |
| Macedonia | National | 64 | 18,8 % | Battaglia et al. (2008) |
| Italy | East Sicily | 114 | 18,4 % | Di Gaetano et al. (2009) |
| Crete | National | 193 | 17,0 % | King et al. (2008) |
| Italy | Sardinia | 930 | 17,0 % | Contu et al. (2008) |
| Armenia | Sasun | 16 | 15,4 % | Herrera et al. (2012) |
| Iran | North | 33 | 15,2 % | Regueiro et al. (2006) |
| Moldova | 268 | 14,6 % | Varzari (2006) | |
| Greece | National | 171 | 13,5 % | King et al. (2008) |
| Turkey | West | 163 | 13,5 % | Cinnioğlu et al. (2004) |
| Romania | National | 54 | 13,0 % | Varzari (2006) |
| Turkey | East | 208 | 12,0 % | Cinnioğlu et al. (2004) |
| Algeria | Northwest (Oran area) | 102 | 11,8 % | Robino et al. (2008) |
| Russia | Roslavl (Smolensk Oblast) | 107 | 11,2 % | Balanovsky et al. (2008) |
| Iraq | National | 139 | 10,8 % | Al-Zahery et al. (2003) |
| Nepal | Newar | 66 | 10,60 % | Gayden et al. (2007) |
| Bulgaria | National | 808 | 10,5 % | Karachanak et al. (2013) |
| Tunisia | Tunis | 139 | 7,2 % | Adams et al. (2008) |
| Algeria | Algiers, Tizi Ouzou | 46 | 6,5 % | Adams et al. (2008) |
| Bosnia-Herzegovina | Serbs | 81 | 6,2 % | Marjanovic et al. (2005) |
| Iran | South | 117 | 6,0 % | Regueiro et al. (2006) |
| Russia | Repyevka (Voronezh Oblast) | 96 | 5,2 % | Balanovsky et al. (2008) |
| UAE | 164 | 3,7 % | Cadenas et al. (2007) | |
| Bosnia-Herzegovina | Bosniaks | 85 | 3,5 % | Marjanovic et al. (2005) |
| Pakistan | 176 | 2,8 % | Sengupta et al. (2006) | |
| Russia | Belgorod | 143 | 2,8 % | Balanovsky et al. (2008) |
| Russia | Ostrov (Pskov Oblast) | 75 | 2,7 % | Balanovsky et al. (2008) |
| Russia | Pristen (Kursk Oblast) | 45 | 2,2 % | Balanovsky et al. (2008) |
| Bosnia-Herzegovina | Croats | 90 | 2,2 % | Marjanovic et al. (2005) |
| Qatar | 72 | 1,4 % | Cadenas et al. (2007) | |
| China | 128 | 0,8 % | Sengupta et al. (2006) | |
| India | various | 728 | 0,5 % | Sengupta et al. (2006) |
| Croatia | Osijek | 29 | 0,0 % | Battaglia et al. (2008) |
| Yemen | 62 | 0,0 % | Cadenas et al. (2007) | |
| Tibet | 156 | 0,0 % | Gayden et al. (2007) | |
| Nepal | Tamang | 45 | 0,0 % | Gayden et al. (2007) |
| Nepal | Kathmandu | 77 | 0,0 % | Gayden et al. (2007) |
| Japan | 23 | 0,0 % | Sengupta et al. (2006) | |
Verbreitung
R1b ist die häufigste Haplogruppe in Westeuropa und im südlichen Ural. Es tritt auch im südlichen Sibirien, Zentralasien, Mittel-Osteuropa, Nordafrika und Westasien auf. Jüngere Auswanderungen von europäischen Kolonisten verbreitete die Gruppe nach Amerika, Australien, Afrika und Indien. Allein 100 Länder gehörten zeitweilig zur englischen Krone, historisch sind noch weitaus mehr Auswanderungen dokumentiert. So finden sich mittlerweile rund um den Erdball Bevölkerungsteile, die dieses Merkmal tragen und durchaus nicht dem europäischen Phänotyp entsprechen müssen. R1b gehört heute neben einer chinesischen Gruppe und der Schwestergruppe R1a zu den einflussreichsten und am weitesten verbreiteten y-Haplogruppen der Welt, wobei mit der Dominanz auch immer eine Verdrängung anderer Haplogruppen einhergeht.
Europa
In Mitteleuropa ist die R1b bisher erst mit der Glockenbecherkultur nach 3000 v. Chr. nachweisbar. Die europäischen Varianten deuten auf einen Gründereffekt hin.[15]
Cassidy et al. identifizierten die Y-DNA dreier frühbronzezeitlicher irischer Männer als R1b (Rathlin 1 2026-3020 v. Chr. R1b1a2a1a2c1g, Rathlin 2 2024-1741 v. Chr. R1b1a2a1a2c1, Rathlin 3 1736-1534 v. Chr. R1b1a2a1a2c). Maternal gehörten sie den Haplogruppen mtDNA U5a1b1e, U5b2a2 und J2b1a an.[16]
Klar ist allerdings, dass in Europa mehrere Ereignisse einer Genkonversion aufgetreten sind, von denen einige sicher mit der Römerzeit in Verbindung gebracht werden können.[17]
Die größten Konzentrationen von R1b finden sich heute in der ansässigen Bevölkerung Westeuropas: Im Süden von England sind es etwa 70 %, im nördlichen und westlichen England, Wales, Schottland, Irland bis über 90 %, in Spanien 70 %, in Frankreich 60 %.[18] In Portugal sind es über 50 %.[19] Bei den Basken sind es 88,1 %. Bei den West-Deutschen sind es 47 %, bei den Italienern 40 %[20] und bei den Norwegern 25,9 %.[21] In Ostdeutschland ist die Verbreitung erheblich geringer, was auf die einzigartige Stellung der Region Deutschland als Schnittmenge eines Ost-West- und Nord-Süd-Gefälles hinweist. Bei den Völkern Mittel- und Osteuropas tritt sie noch seltener auf, bei den Tschechen und Slowaken 35,6 %, bei den Polen 16,4 %, bei den Letten 15 %, bei den Ungarn 13,3 %, im europäischen Teil Russlands ist es nur noch auf einige Inseln und Metropolen beschränkt. Man kann also von einem West-Ost-Gefälle sprechen, das sich bis zur Wolga hinzieht und im östlichen Ural bis zum Altai wieder ansteigt.
Ural
Außerhalb Europas tritt eine vergleichsweise hohe Konzentration der Haplogruppe R1b bei einigen Populationen der Baschkiren (Burzyan, Gayna und andere) von bis zu 87 % auf, speziell in den Regionen Perm (84,0 %) und Baymaksky (81,0 %) im Südosten von Baschkortostan.[22] Bei den Baschkiren sind auch die Mutationen M269 und M73 häufig. Trofimova et al. (2015) fand eine überraschend hohe Frequenz von R1b-L23 (Z2105/Z2103) bei den Menschen der Wolga-Ural-Region (östlich der Wolga): 21 von 58 (36,2 %) unter den Baschkiren in der südöstlichen Burzyan-Region von Baschkortostan; 11 von 52 (21,2 %) unter den Udmurten sowie unter den Komi (8 %), Mordwinen (6,8 %), Bessermenen (3,8 %) und Tschuwaschen (2,3 %) trugen R1b-L23 (Z2105/2103),[23] derselbe Typ wurde für die Yamna-Kultur im Samara Oblast und im Orenburg Oblast festgestellt. Diese Präsenz blieb für 2000 Jahre relativ stabil.
Asien
In Iran, die Assyrer, R1b-M269 um 55 %.[24]
Die altaische Variante der Haplogruppe R1b-M73 tritt mit 49 % bei Kumandinern auf.[25]
Afrika
Bei Kamerun-Mandara wurden bis zu 65 % gefunden.[26] Auf São Tomé und Príncipe wurden 8,7 % Träger der Haplogruppe R1b ermittelt, in der Bevölkerungsgruppe der Forros bis zu 17,7 %.[27] Neuere Studien vermuten eine Rückwanderung aus Eurasien in vorislamischer Zeit, jedenfalls für die Gruppen in Kamerun.[28][29]
Einzelnachweise
- T. M. Karafet, F. L. Mendez, M. B. Meilerman, P. A. Underhill, S. L. Zegura, M. F. Hammer: New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. In: Genome research. Band 18, Nummer 5, Mai 2008, S. 830–838, ISSN 1088-9051. doi:10.1101/gr.7172008. PMID 18385274. PMC 2336805 (freier Volltext).
- Anatoly A. Klyosov, Giancarlo T. Tomezzoli, "DNA Genealogy and Linguistics. Ancient Europe." Scientific Research. The Academy of DNA Genealogy. Newton, USA 2013. European Patent Office, Munich, Germany. Vol.: 3, No.: 2, Seiten.: 101-111. doi:10.4236/aa.2013.32014.
- C. Cinnioglu, R. King, T. Kivisild, E. Kalfoglu, S. Atasoy, G. L. Cavalleri, A. S. Lillie, C. C. Roseman, A. A. Lin, K. Prince, P. J. Oefner, P. Shen, O. Semino, L. L. Cavalli-Sforza, P. A. Underhill: Excavating Y-chromosome haplotype strata in Anatolia. In: Human genetics. Band 114, Nummer 2, Januar 2004, S. 127–148, ISSN 0340-6717. doi:10.1007/s00439-003-1031-4. PMID 14586639.
- http://isogg.org/tree/ISOGG_HapgrpR.html
- https://docs.google.com/spreadsheets/d/1n9MBaZWKBWUx2DN9aEN0CLCDmtnp64Hts-GrYGGPRRI/edit?pli=1#gid=0
- T. M. Karafet, F. L. Mendez, M. B. Meilerman, P. A. Underhill, S. L. Zegura, M. F. Hammer: New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. In: Genome research. Band 18, Nummer 5, Mai 2008, S. 830–838, ISSN 1088-9051. doi:10.1101/gr.7172008. PMID 18385274. PMC 2336805 (freier Volltext).
- Variations of R1b Ydna in Europe: Distribution and Origins. Архивировано из первоисточника 25 марта 2012.
- Wolfgang Haak, Iosif Lazaridis, et. al: Massive migration from the steppe was a source for Indo-European languages in Europe. Nature volume 522, S. 207–211 (2015) (auf nature.com)
- Wolfgang Haak, et al: Massive migration from the steppe was a source for Indo-European languages in Europe. Nature. 2015 Jun 11; 522(7555): 207–211 ( auf ncbi.nlm.nih.gov)
- Thorwald Ewe: Die geheimnisvollen Dritten. bild der wissenschaft, 12. Ausgabe Dezember 2020 ( auf wissenschaft.de)
- Haak et al. (2015)
- siehe offizielle Webseite
- Characterization of the Iberian Y chromosome haplogroup R-DF27 in Northern Spain, P. Villaescusa at all Dez. 2016, doi:10.1016/j.fsigen.2016.12.013
- Dabei unterschieden sich die einzelnen Ethnien zum Teil deutlich. Die Identität der Ethnien wird in der Forschung traditionell über die Sprache bestimmt wird. Kennzeichen der germanischen Sprachen sind unter anderen bestimmte Lautwandel gegenüber der rekonstruierten indogermanischen Ursprache, die als germanische oder erste Lautverschiebung zusammengefasst werden.
- N. M. Myres, S. Rootsi, A. A. Lin, M. Järve, R. J. King, I. Kutuev, V. M. Cabrera, E. K. Khusnutdinova, A. Pshenichnov, B. Yunusbayev, O. Balanovsky, E. Balanovska, P. Rudan, M. Baldovic, R. J. Herrera, J. Chiaroni, J. Di Cristofaro, R. Villems, T. Kivisild, P. A. Underhill: A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe. In: European journal of human genetics : EJHG. Band 19, Nummer 1, Januar 2011, S. 95–101, ISSN 1476-5438. doi:10.1038/ejhg.2010.146. PMID 20736979. PMC 3039512 (freier Volltext).
- Lara M. Cassidy et al., Neolithic and Bronze Age migration to Ireland and establishment of the insular Atlantic genome, 2015.
- S. M. Adams, T. E. King, E. Bosch, M. A. Jobling: The case of the unreliable SNP: recurrent back-mutation of Y-chromosomal marker P25 through gene conversion. In: Forensic science international. Band 159, Nummer 1, Mai 2006, S. 14–20, ISSN 0379-0738. doi:10.1016/j.forsciint.2005.06.003. PMID 16026953.
- Ornella Semino, A. Silvana Santachiara-Benerecetti, Francesco Falaschi, L. Luca Cavalli-Sforza and Peter A. Underhill, "Ethiopians and Khoisan Share the Deepest Clades of the Human Y-Chromosome Phylogeny, " The American Journal of Human Genetics, Volume 70, Issue 1, 265—268, 1 January 2002.
- R. Gonçalves, A. Freitas, M. Branco, A. Rosa, A. T. Fernandes, L. A. Zhivotovsky, P. A. Underhill, T. Kivisild, A. Brehm: Y-chromosome lineages from Portugal, Madeira and Açores record elements of Sephardim and Berber ancestry. In: Annals of Human Genetics. Band 69, Pt 4Juli 2005, S. 443–454, ISSN 0003-4800. doi:10.1111/j.1529-8817.2005.00161.x. PMID 15996172.
- C. Capelli, F. Brisighelli, F. Scarnicci, B. Arredi, A. Caglia', G. Vetrugno, S. Tofanelli, V. Onofri, A. Tagliabracci, G. Paoli, V. L. Pascali: Y chromosome genetic variation in the Italian peninsula is clinal and supports an admixture model for the Mesolithic-Neolithic encounter. In: Molecular phylogenetics and evolution. Band 44, Nummer 1, Juli 2007, S. 228–239, doi:10.1016/j.ympev.2006.11.030, PMID 17275346.
- Estimating Scandinavian and Gaelic Ancestry in the Male Settlers of Iceland — Agnar Helgason et al., 2000, Am. J. Hum. Genet. 67:697-717, 2000
- Лобов А. С. (2009) «Структура генофонда субпопуляций башкир» (автореферат диссертации) (Memento des Originals vom 16. August 2011 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- Трофимова Натал'я Вадимовна: Изменчивость Митохондриальной ДНК и Y-Хромосомы в Популяциях Волго-Уральского Региона. Zusammenfassung. Dissertation für den Grad des Kandidaten für biologische Wissenschaften. Februar 2015, abgerufen am 1. Juli 2018.
- V. Grugni, V. Battaglia, B. Hooshiar Kashani, S. Parolo, N. Al-Zahery, A. Achilli, A. Olivieri, F. Gandini, M. Houshmand, M. H. Sanati, A. Torroni, O. Semino: Ancient migratory events in the Middle East: new clues from the Y-chromosome variation of modern Iranians. In: PloS one. Band 7, Nummer 7, 2012, S. e41252, ISSN 1932-6203. doi:10.1371/journal.pone.0041252. PMID 22815981. PMC 3399854 (freier Volltext).
- Балаганская О. А. Полиморфизм Y-хромосомы у тюркоязычного населения Алтая, Саян, Тянь-Шаня и Памира в контексте взаимодействия генофондов Западной и Восточной Евразии // Автореферат кандидатской диссертации по биологическим наукам. М., МГНЦ РАМН, 2011, С.10.
- Wood et al. 2005.
- R. Gonçalves, H. Spínola, A. Brehm: Y-chromosome lineages in São Tomé e Príncipe islands: evidence of European influence. In: American journal of human biology : the official journal of the Human Biology Council. Band 19, Nummer 3, 2007 May-Jun, S. 422–428, ISSN 1042-0533. doi:10.1002/ajhb.20604. PMID 17420998.
- ET Wood, Stover, Ehret, Destro-Bisol, Spedini, Mcleod, Louie, Bamshad, Strassmann: Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes Archiviert vom Original am 26. Juni 2008. In: European Journal of Human Genetics. 13, Nr. 7, 2005, S. 867–76. doi:10.1038/sj.ejhg.5201408. PMID 15856073.
- F Cruciani, Santolamazza, Shen, Macaulay, Moral, Olckers, Modiano, Holmes, Destro-Bisol: A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes. In: American Journal of Human Genetics. 70, Nr. 5, 2002, S. 1197–214. doi:10.1086/340257. PMID 11910562. PMC 447595 (freier Volltext)., pp. 13–14
| Evolutionsbaum Haplogruppen Y-chromosomale DNA (Y-DNA) | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adam des Y-Chromosoms | ||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0’1'2’3'4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1’2'3’4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A1 | A2’3'4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2’3 | A4=BCDEF | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2 | A3 | B | CT | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| G | IJK | H | ||||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| G1 | G2 | IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | L | K(xLT) | T | ||||||||||||||||||||||||||||||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||||||||||||||||||||||||||
| I1 | I2 | J1 | J2 | M | NO | P | S | |||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| R1 | R2 | |||||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| R1a | R1b | |||||||||||||||||||||||||||||||||||||||||||||||||