Vault (Organell)
Vault oder zytoplasmatisches (bzw. zytoplasmisches) Ribonukleoprotein Vault ist die Bezeichnung für bestimmte eukaryotische Organellen, deren Funktion noch nicht vollständig geklärt ist. Bei den Vaults[2] handelt es sich um im Zytoplasma von Eukaryoten befindliche Organellen. Sie wurden 1986 entdeckt, als die Zellbiologin Nancy Kedersha und der Biochemiker Leonard Rome von der University of California, Los Angeles (UCLA) sie aus der Leber von Ratten isolierten. Die Vaults erscheinen bei Negativfärbung (englisch negative staining) unter dem Elektronenmikroskop als tonnen- oder fassförmige Partikel mit einer 39-fachen diskreten Dreh- und Inversionssymmetrie (D39d in Schoenflies-Notation). Ihre Feinstruktur ähnelt den Gewölbebögen einer gotischen Kathedrale (mit zwei aufeinandergesetzten Kuppelhälften), wovon sich ihre Bezeichnung ableitet (englisch vault ‚Gewölbe‘).[1] Sie kommen in vielen Arten eukaryotischer Zellen vor und scheinen in dieser Domäne stark konserviert zu sein.[3] Nicht bei allen Organismen konnte im Innern dort gefundener Vaults tatsächlich ein Nukleinsäure-Anteil (Vault-RNA, vRNA) nachgewiesen werden.



Gestalt und Aufbau
Morphologie
Vaults sind vergleichsweise große Partikel aus Ribonukleoprotein; etwa dreimal so groß wie die ebenfalls aus Ribonukleoprotein bestehenden (eukaryotischen) Ribosomen und haben eine Masse von etwa 13 MDa (Mega-Dalton). Sie kommen in den meisten eukaryotischen Zellen, insbesondere bei allen höheren Eukaryonten (wie Tieren, Pflanzen und Pilzen) vor. Die Färbung mit herkömmlichen Techniken ist erschwert durch den Umstand, dass die Vaults in erster Linie aus Proteinen bestehen. Während ihre Größe aufgrund von konventioneller Elektronenmikroskopie (en. conventional transmission electron microscopy, CTEM) mit Negativfärbung ursprünglich auf 35 × 35 × 65 nm geschätzt wurde[4] und sie im Rastertransmissionselektronenmikroskopie (STEM) 35 × 35 × 59 nm groß erscheinen, wurden diese Angaben mit Hilfe der Kryoelektronenmikroskopie (Cryo-EM) von ursprünglich 35 × 35 × 59 nm auf 41 × 41 × 72,5 nm verfeinert. Die Vaults sind damit die größten bekannten Nukleoproteinstrukturen mit nicht-ikosaedrischer Symmetrie (übertroffen nur von Nukleoproteinen bei Viren mit ikosaedrischer Symmetrie). Die Vault-Proteine bilden im Innern einen Hohlraum; die äußere Hülle oder Schale (en. outer shell) hat dabei eine Wandstärke von 1,5 bis 2,4 nm. Ihre beiden symmetrischen Hälften („Kuppelhälften“) bestehen je aus drei Teilen, die einen Körper (en. body), eine Schulterregion (Plexus) und eine Kappe (en. cap) bilden. Die Kappenhöhe beträgt ∼15,5 nm, der Außendurchmesser erreicht ∼13 nm.[5][6]
Proteinstruktur (MVP)
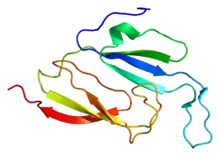
Die Proteinstruktur des „Körpers“ besteht aus einer äußeren Hülle aus insgesamt 78 (je Hälfte 39) Kopien des 95,8 kDa (Kilodalton) großen Vault-Hauptproteins (en. major vault protein, MVP), das etwa 70 % des gesamten Proteins der Partikel ausmacht. Die MVP-Untereinheiten sind Kopf-an-Kopf zusammengesetzt, wobei die N-Termini jeder Kuppelhälfte einander gegenüberliegen. Entsprechend der Form der Vault-Hälfte faltet sich das MVP vom N-Terminus zum C-Terminus in folgende Untereinheiten: neun sich wiederholende Strukturdomänen (R1-R9) inklusive der Schulterdomäne (R1) im sich verjüngenden Abschnitt; dazu im Bereich der Kappe eine „Cap-Helix-Domäne“ und eine „Cap-Ring-Domäne“ (in 39 Kopien in jeder der beiden Kappen).[5]
Die Strukturdomänen R8 und R9 bestehen aus fünf antiparallelen β-Faltblättern, die mit S1 bis S5 bezeichnet werden. Die anderen sieben Strukturdomänen (R1 bis R7) haben zwei zusätzliche β-Faltblätter (S2a und S2b), die zwischen S2 und S3 eingefügt sind.[5]
Die Schulterregion faltet sich zu einer einzigen globulären α/β-Domäne mit 4 antiparallelen β-Faltblättern auf einer Seite und vier α-Helices auf der anderen Seite. Offenbar befinden sich im Schulterbereich die Elemente, die für die Interaktion des Vaults mit Lipid-Rafts verantwortlich sind.[5]
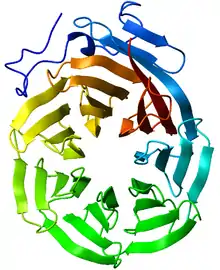
Die Cap-Helix-Domäne faltet sich zu einer α-Helix mit 42 Turns, die in eine Superspirale passt. Die Cap-Ring-Domäne befindet sich am Ende der Kappe und bildet eine U-förmige Struktur mit spiralförmigen Elementen an beiden Enden.[5]
Proteinstruktur (andere)
Im Inneren der Partikel befinden sich zwei assoziierte Vault-Proteine, TEP1 (Telomerase-assoziiertes Protein 1) und VPARP alias Poly-(ADP-Ribose)-Polymerase 4 (PARP4).[7] TEP1 hat eine Größe von 290 KDa und VPARP von 193 KDa.[8]
Das TEP1-Protein scheint sich an der Spitze des flachen Teils der Kappe zu befinden, wo seine WD40-Domäne eine ringförmige 7-blättrige β-Propeller-Faltung (en. 7-bladed beta-propeller fold) bildet. Dieser TEP1-Ring und bindet an die Cap-Domäne, wobei ein bestimmter Typ der vRNA die Cap-Domäne füllt.[5] Der N-terminale Teil von TEP1 enthält 4 sich wiederholende Domänen mit unklarer Funktion, eine RNA- Bindungsdomäne und eine ATP/GTP-Bindungsdomäne. Man konnte zeigen, dass TEP1 mit Telomerase-RNA und verschiedenen humanen vRNAs interagiert.[5]
VPARPs befinden sich hauptsächlich in der Vault-Kappe.[5]
vRNAs
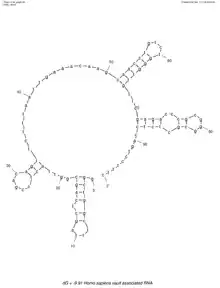
Vaults aus höheren Eukaryonten enthalten auch eine oder mehrere kleine Vault-RNAs (vRNAs, auch vtRNAs) mit 86–141 nt (Nukleinbasen).[12] Die vRNAs befinden sich in den Kappen an den Enden der Vaults.[5] Die Gesamt-Masse der RNA im Vault wird auf 460 kDa geschätzt.[5] Die nicht-kodierende RNA der multiresistenten Vaults kodiert für mehrere regulatorische kleine RNAs (en. regulatory small RNAs).[13]
Funktion
Die Funktion der Vaults noch nicht vollständig aufgeklärt, sie werden aber mit den Kernporenkomplexen Kernporen (en. nuclear pore complex, NPC) in Verbindung gebracht. Insbesondere ihre achteckige Form scheint dies zu unterstützen.[14][15] Darüber hinaus werden Vaults mit einem breiten Spektrum von Zellfunktionen in Verbindung gebracht, darunter der Transport zwischen Zellkern und Zytoplasma (en. nuclear-cytoplasmic transport), die mRNA-Lokalisierung, der Resistenz gegenüber Medikamenten, der Signalübertragung (en. cell signaling), dem Zusammenbau (Assemblierung) der Kernporen und angeborener Immunität.[16] Die drei Vault-Proteine (MVP, VPARP und TEP1) wurden jeweils einzeln und in Kombination (etwa VPARP und TEP1) bei Mäusen ausgeschaltet (siehe „Knockout-Maus“).[17][18][19] Alle Knockout-Mäuse sind lebensfähig, und es wurden keine größeren phänotypischen Veränderungen beobachtet. Dictyostelium kodiert für drei verschiedene MVPs, von denen zwei einzeln und in Kombination ausgeschaltet wurden.[20] Die einzige Abweichung im Phänotyp, die bei dem Doppel-Knockout von Dictyostelium beobachtet wurde, war eine Wachstumsverzögerung unter Ernährungsstress.[21] Die Schlussfolgerung war, dass wenn Vaults tatsächlich an essenziellen zellulären Funktionen beteiligt sein sollten, dann existieren wahrscheinlich redundante Systeme, die ihren Verlust ausgleichen können.
Zusammenhang mit Krebs
In den späten 1990er Jahren fand man heraus, dass bei Krebspatienten, bei denen eine Multiresistenz (d. h. eine Resistenz gegen viele Chemotherapien) diagnostiziert wurde, die Vaults (insbesondere das MVP) übermäßig stark ausgeprägt waren.[22] Auch wenn dies nicht beweist, dass eine erhöhte Anzahl von Vaults zu einer Arzneimittelresistenz führt, deutet dies auf eine gewisse Beteiligung hin. Möglicherweise gibt es hier ein Potenzial, die Mechanismen hinter der Arzneimittelresistenz in Tumorzellen zu erforschen und in der Folge Krebsmedikamente zu verbessern.[20] Einen Hinweis darauf gibt die Entdeckung, dass die Phosphatase PTEN (en. Phosphatase and Tensin homolog) in HeLa-Zellen mit den Vault-Partikeln assoziiert ist.[23] Eine neuere Studie zum Zusammenhang von Vaults und Krebs von Lara et al. erschien 2011.[24]
Entwicklungsgeschichte und Konservierung
Vaults wurden bei Säugetieren, Amphibien, Vögeln und dem Schleimpilz Dictyostelium discoideum identifiziert.[3] Das von der Pfam-Datenbank verwendete Vault-Modell identifiziert Homologe in Paramecium tetraurelia, Kinetoplastida, vielen Wirbeltieren, einem Nesseltier (Seeanemone), Mollusken, Trichoplax adhaerens, Plattwürmern (Echinococcus granulosus) und Choanoflagellaten.[25]
Vaults wurden bei vielen eukaryotischen Arten beobachtet, allerdings scheinen einige Arten kein Ribonukleoprotein zu besitzen. Dazu gehören:[26]
- Arabidopsis thaliana (Ackerschmalwand, eine Pflanze aus der Familie der Brassicaceae, zu der Kohl, Rüben, Raps, Senf, Meerrettich, Kresse, … gehören)
- Caenorhabditis elegans (ein Fadenwurm)
- Drosophila melanogaster (Taufliege)
- Saccharomyces cerevisiae (Backhefe)
Diese vier Arten sind Modellorganismen für Pflanzen, Fadenwürmer, Tiergenetik bzw. Pilze. Trotz dieser Ausnahmen lässt der hohe Grad an Ähnlichkeit der Vaults bei den Organismen, bei denen sie gefunden wurden, auf eine gewisse evolutionäre Bedeutung schließen.[3]
Bei Sequenzvergleichen in den Gendatenbanken (in silico) wurden Homologe des wichtigsten Vault-Proteins (MVP) auch in Bakterien gefunden; Cyanbakterielle Sequenzen scheinen darunter am ähnlichsten zu sein. Auch mit Pfam lassen sich derartige Homologe identifizieren.[27] – Siehe auch den PrePrint von Tymofii Sokolskyi (2019).
Anwendungen
Das Rome-Labor an der UCLA (University of California, Los Angeles) hat mit einer Reihe von Gruppen zusammengearbeitet, um das Baculovirus-System zur Herstellung großer Mengen von Vaults zu nutzen. Wenn das Haupt-Vault-Protein (MVP) in Insektenzellen exprimiert wird, werden die Vault-Partikel auf Polyribosomen im Zytoplasma zusammengebaut.[28] Durch den Einsatz molekulargenetischer Techniken zur Veränderung des Gens, das für das MCP kodiert, wurden Vault-Partikel hergestellt, an deren Sequenz chemisch aktive Peptide gebunden sind. Diese modifizierten Proteine werden in das Innere der Vault-Partikels eingebaut, ohne deren Grundstruktur zu verändern. Proteine und Peptide können alternativ auch in Vaults verpackt werden, indem eine vom VPARP-Protein abgeleitete „Verpackungsdomäne“ (en. packaging domain) angehängt wird. Es wurde so eine Reihe modifizierter Vault-Partikel hergestellt. Damit wurde getestet, ob Vaults biotechnologisch so verändert werden können, dass sie in einer Vielzahl biologischer Anwendungen eingesetzt werden können, z. B. zur Verabreichung von Medikamenten, als biologische Sensoren, zur Verabreichung von Enzymen, bis hin zur kontrollierten Freisetzung (en. controlled release; verzögerte, langsame Freisetzung eines Medikaments etc. nach der eigentlichen Verabreichung) und der Umweltsanierung. Beispielsweise wurde ein Vault mit einem Chemokin verpackt, um das Immunsystem zur Bekämpfung von Lungenkrebs zu aktivieren; dieser Ansatz wurde in Phase I getestet.[29][30]
Weblinks
- The structure of rat liver vault at 3.5 angstrom resolution, auf: RSCB Protein Data Bank (PDB)
- Vaults: Novel Cell Particles — welcome to the vault website! — What are Vaults? (UCLA, Memento im WebArchiv vom 28. Februar 2009)
- Vault Pharma Inc.
- Vault+Ribonucleoprotein+Particles (Vault+Ribonukleoprotein+Partikel) in der US National Library of Medicine — Medical Subject Headings (MeSH)
- Family: Vault (PF01505), EML-EBI Pfam-Eintrag
- Major vault protein, N-terminal IPR002499, InterPro-Eintrag
- MVP (vault) repeat profile, Expasy Prosite Dokumentation PDOC51224
- Vault RNA, Seite in der Rfam-Datenbank
- Major vault protein auf Proteopedia
- Tymofii Sokolskyi: Bacterial Major Vault Protein homologs shed new light on origins of the enigmatic organelle, PrePrint auf: bioRxiv vom 12. Dezember 2019, doi:10.1101/872010.
- Leonard H. Rome, Valerie A Kickhoefer: Development of the vault particle as a plat#x200B;form technology. In: ACS Nano, Band 7, Nr. 2, 26. Februar 2013, S. 889–902, doi:10.1021/nn3052082, PMID 23267674, Epub 31. Dezember 2012.
- Eneida V. Reis, Roberta V. Pereira, Matheus Gomes, Liana K. Jannotti-Passos, Elio H. Baba, Paulo Marcos Zech Coelho, Ana C. A. Mattos, Flávia F. B. Couto, William Castro-Borges, Renata Guerra-Sá: Characterisation of major vault protein during the life cycle of the human parasite Schistosoma mansoni. In: Parasitology international! Band 63, Nr. 1, Februar 2014, S. 120–126, doi:10.1016/j.parint.2013.10.005, PMID 24148287, Epub 19. Oktober 2013.
- Arnau Casañas, Jordi Querol-Audí, Pablo Guerra, Joan Pous, Hideaki Tanaka, Tomitake Tsukihara, Nuria Verdaguer, Ignasi Fita: New features of vault architecture and dynamics revealed by novel refinement using the deformable elastic network approach. In: Acta crystallographica. Section D, Biological crystallography, Band 69, Pt 6, Juni 2013, S. 1054#x200B;—1061, doi:10.1107/S0907444913004472, PMID 23695250, Epub 14. Mai 2013.
Einzelnachweise
- Hideaki Tanaka, Koji Kato, Eiki Yamashita, Tomoyuki Sumizawa, Yong Zhou, Min Yao, Kenji Iwasaki, Masato Yoshimura, Tomitake Tsukihara: The structure of rat liver vault at 3.5 angstrom resolution. In: Science. 323, Nr. 5912, Januar 2009, S. 384–388. bibcode:2009Sci...323..384T. doi:10.1126/science.1164975. PMID 19150846.
- Nancy L. Kedersha, Leonard H. Rome: Isolation and characterization of a novel ribonucleoprotein particle: large structures contain a single species of small RNA. In: The Journal of Cell Biology. 103, Nr. 3, September 1986, S. 699–709. doi:10.1083/jcb.103.3.699. PMID 2943744. PMC 2114306 (freier Volltext).
- Nancy L. Kedersha, Marie-Christine Miquel, Dennis Bittner, Leonard H. Rome: Vaults. II. Ribonucleoprotein structures are highly conserved among higher and lower eukaryotes. In: The Journal of Cell Biology. 110, Nr. 4, April 1990, S. 895–901. doi:10.1083/jcb.110.4.895. PMID 1691193. PMC 2116106 (freier Volltext).
- Nancy L. Kedersha, John E. Heuser, Diane C. Chugani, Leonard H. Rome: Vaults. III. Vault ribonucleoprotein particles open into flower-like structures with octagonal symmetry. In: The Journal of Cell Biology. 112, Nr. 2, Januar 1991, S. 225–235. doi:10.1083/jcb.112.2.225. PMID 1988458. PMC 2288824 (freier Volltext).
- Hideaki Tanaka, Tomitake Tsukihara: Structural studies of large nucleoprotein particles, vaults. In: Proceedings of the Japan Academy, Series B. 88, Nr. 8, 2012, S. 416–433. bibcode:2012PJAB...88..416T. doi:10.2183/pjab.88.416. PMID 23060231. PMC 3491081 (freier Volltext).
- Aida Llauró, Pablo Guerra, Nerea Irigoyen, José F. Rodríguez, Núria Verdaguer, Pedro J. de Pablo: Mechanical stability and reversible fracture of vault particle. In: Biophys J., Band 106, Nr. 3, 4. Februar 2014, S. 687–695, doi:10.1016/j.bpj.2013.12.035, PMID 24507609, PMC 3945774 (freier Volltext). Siehe insbes. Fig. 1
- Valerie A. Kickhoefer, Andrew G. Stephen, Lea Harrington, Murray O. Robinson, Leonard H. Rome: Vaults and telomerase share a common subunit, TEP1. In: The Journal of Biological Chemistry. 274, Nr. 46, November 1999, S. 32712–32717. doi:10.1074/jbc.274.46.32712. PMID 10551828.
- Valerie A. Kickhoefer, Amara C. Siva, Nancy L. Kedersha, Elisabeth M. Inman, Cristina Ruland, Michel Streuli, Leonard H. Rome: The 193-kD vault protein, VPARP, is a novel poly(ADP-ribose) polymerase. In: The Journal of Cell Biology. 146, Nr. 5, September 1999, S. 917–928. doi:10.1083/jcb.146.5.917. PMID 10477748. PMC 2169495 (freier Volltext).
- GenBank.
- The mfold web server. In: mfold.rna.albany.edu.
- Rfam predict (Rfam Vorhersage)
- A. van Zon, M. H. Mossink, R.. J. Scheper, P. Sonneveld, E.. A.. C. Wiemer: The vault complex. In: Cellular and Molecular Life Sciences. 60, Nr. 9, September 2003, S. 1828–1837. doi:10.1007/s00018-003-3030-y. PMID 14523546.
- Helena Persson, Anders Kvist, Johan Vallon-Christersson, Patrik Medstrand, Ake Borg, Carlos Rovira: The non-coding RNA of the multidrug resistance-linked vault particle encodes multiple regulatory small RNAs. In: Nat Cell Biol, Band 11, Nr. 10, Oktober 2009, S. 1268-1271, doi:10.1038/ncb1972, PMID 19749744, Epub 13. September 2009.
- Diane C. Chugani, Leonard H. Rome, Nancy L. Kedersha: Evidence that vault ribonucleoprotein particles localize to the nuclear pore complex. In: Journal of Cell Science. 106 ( Pt 1), September 1993, S. 23–29. doi:10.1242/jcs.106.1.23. PMID 8270627.
- P. Nigel T. Unwin, Ronald A. Milligan: A large particle associated with the perimeter of the nuclear pore complex. In: The Journal of Cell Biology. 93, Nr. 1, April 1982, S. 63–75. doi:10.1083/jcb.93.1.63. PMID 7068761. PMC 2112107 (freier Volltext).
- Walter Berger, E. Steiner, Michael Grusch, Leonilla Elbling, Michael Micksche: Vaults and the major vault protein: novel roles in signal pathway regulation and immunity. In: Cellular and Molecular Life Sciences. 66, Nr. 1, 2008/2009, S. 43–61. doi:10.1007/s00018-008-8364-z. PMID 18759128.
- Valerie A. Kickhoefer, Yie Liu, Lawrence B. Kong, Bryan E. Snow, Phoebe L. Stewart, Lea Harrington, Leonard H. Rome: The Telomerase/vault-associated protein TEP1 is required for vault RNA stability and its association with the vault particle. In: The Journal of Cell Biology. 152, Nr. 1, Januar 2001, S. 157–164. doi:10.1083/jcb.152.1.157. PMID 11149928. PMC 2193651 (freier Volltext).
- Yie Liu, Bryan E. Snow, M. Prakash Hande, Gabriela Baerlocher, Valerie A. Kickhoefer, David Yeung, Andrew Wakeham, Annick Itie, David P. Siderovski, Peter M. Lansdorp, Murray O. Robinson, Lea Harrington: Telomerase-associated protein TEP1 is not essential for telomerase activity or telomere length maintenance in vivo. In: Molecular and Cellular Biology. 20, Nr. 21, November 2000, S. 8178–8184. doi:10.1128/mcb.20.21.8178-8184.2000. PMID 11027287. PMC 86427 (freier Volltext).
- Marieke H. Mossink, Arend van Zon, Rik J. Scheper, Pieter Sonneveld, Erik A. C. Wiemer: Disruption of the murine major vault protein (MVP/LRP) gene does not induce hypersensitivity to cytostatics. In: Cancer Research. 62, Nr. 24, Dezember 2002, S. 7298–7304. PMID 12499273.
- Valerie A. Kickhoefer, Sanjay K. Vasu, Leonard H. Rome: Vaults are the answer, what is the question?. In: Trends in Cell Biology. 6, Nr. 5, Mai 1996, S. 174–178. doi:10.1016/0962-8924(96)10014-3. PMID 15157468.
- Leonard H. Rome, Valerie A. Kickhoefer: Development of the vault particle as a plat form technology. In: ACS Nano. 7, Nr. 2, Februar 2013, S. 889–902. doi:10.1021/nn3052082. PMID 23267674.
- Marieke H. Mossink, Arend van Zon, Rik J. Scheper, Pieter Sonneveld, Erik A. C. Wiemer: Vaults: a ribonucleoprotein particle involved in drug resistance?. In: Oncogene. 22, Nr. 47, Oktober 2003, S. 7458–7467. doi:10.1038/sj.onc.1206947. PMID 14576851.
- Z. Yu, N. Fotouhi-Ardakani, L. Wu, M. Maoui, S. Wang, D. Banville, Shen SH: PTEN associates with the vault particles in HeLa cells. In: The Journal of Biological Chemistry. 277, Nr. 43, Oktober 2002, S. 40247–54022. doi:10.1074/jbc.M207608200. PMID 12177006.
- Pedro C. Lara, Martin Pruschy, Martina Zimmermann, Luis Alberto Henríquez-Hernández: MVP and vaults: a role in the radiation response. In: Radiat Oncol, Band 6, Nr. 148, Oktober 2011, doi:10.1186/1748-717X-6-148, PMID 22040803, PMC 3216873 (freier Volltext).
- Family: Vault (PF01505), EMBL-EBI: Major Vault Protein repeat Pfam family
- Leonard H. Rome, Nancy L. Kedersha, Diane C. Chugani: Unlocking vaults: organelles in search of a function. In: Trends in Cell Biology. 1, Nr. 2–3, August 1991, S. 47–50. doi:10.1016/0962-8924(91)90088-Q. PMID 14731565.
- Toni K. Daly, Andrew J. Sutherland-Smith, David Penny: In silico resurrection of the major vault protein suggests it is ancestral in modern eukaryotes.. In: Genome Biology and Evolution. 5, Nr. 8, 2013, S. 1567#x200B;–1583. doi:10.1093/gbe/evt113. PMID 23887922. PMC 3762200 (freier Volltext).
- Jan Mrazek, Daniel Toso, Sergey Ryazantsev, Xing Zhang, Z. Hong Zhou, Beatriz Campo Fernandez, Valerie A. Kickhoefer, Leonard H. Rome: Polyribosomes are molecular 3D nanoprinters that orchestrate the assembly of vault particles. In: ACS Nano. 8, Nr. 11, November 2014, S. 11552–11559. doi:10.1021/nn504778h. PMID 25354757. PMC 4245718 (freier Volltext).
- Sherven Sharma, Li Zhu, Minu K. Srivastava, Marni Harris-White, Min Huang, Jay M. Lee, Fran Rosen, Gina Lee, Gerald Wang, Valerie A. Kickhoefer, Leonard H. Rome, Felicita Baratelli, Maie St John, Karen Reckamp, Seok Chul-Yang, Sven Hillinger, Robert Strieter, Steven M. Dubinett: CCL21 Chemokine Therapy for Lung Cancer. In: International Trends in Immunity. 1, Nr. 1, Januar 2013, S. 10–15. PMID 25264541. PMC 4175527 (freier Volltext).
- Upendra K. Kar, Minu K. Srivastava, Åsa Andersson, Felicita Baratelli, Min Huang, Valerie A. Kickhoefer, Steven M. Dubinett, Leonard H. Rome, Sherven Sharma: Novel CCL21-vault nanocapsule intratumoral delivery inhibits lung cancer growth. In: PLOS ONE. 6, Nr. 5, 3. Mai 2011, S. e18758. bibcode:2011PLoSO...618758K. doi:10.1371/journal.pone.0018758. PMID 21559281. PMC 3086906 (freier Volltext).
{kind=link}