Pantoffeltierchen
Die Pantoffeltierchen (Paramecium) sind eine Gattung von rundum bewimperten Protisten. Das Merkmal „Zilien“ („Wimpern“) haben die Pantoffeltierchen mit einigen anderen Einzellern gemeinsam, die man traditionell zu den Wimpertierchen (Ciliata, Ciliophora) zählt. Als erste Protisten ernannte die Deutsche Gesellschaft für Protozoologie die Vertreter der Gattung Paramecium im Jahr 2007 zu den Einzellern des Jahres.
| Pantoffeltierchen | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
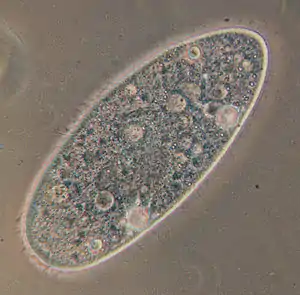
Paramecium aurelia | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Paramecium | ||||||||||||
| O.F. Müller, 1773 |
Vorkommen
Pantoffeltierchen leben vorwiegend im Süßwasser, einige Arten wie Paramecium woodruffi sind aber euryhalin und kommen auch im Brackwasser von Flussmündungen, selten sogar im Meer, vor.[1] Die meisten Vertreter trifft man in Gewässern wie z. B. Tümpeln, Teichen, Seen, Flüssen, aber auch in Wasserpfützen an. Somit sind sie ein wichtiger Bestandteil des Ökosystems Süßwasser. Pantoffeltierchen sind häufig vorkommende Ciliaten. Die Gattung Paramecium bildet im Gegensatz zu vielen anderen Einzellern keine Cysten als Überdauerungsstadien aus.[2]
Aussehen
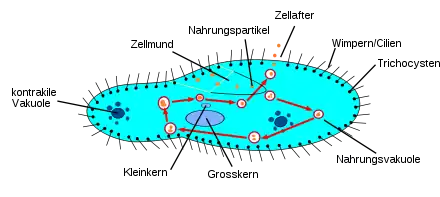
Pantoffeltierchen sind frei lebende, sich aktiv bewegende Einzeller mit einer Länge zwischen 50 und 300 Mikrometern. In Aufsicht ist ihre Körperform langgestreckt oval („zigarrenförmig“) bis rundlich, wobei das Vorderende mehr oder weniger asymmetrisch schief abgeschnitten (also sohlen- oder „pantoffelförmig“) wirkt. Im Wasser vermögen sie sich frei schwimmend schnell zu bewegen, wobei sie sich fast immer nach links spiralförmig um ihre Achse drehen. Von oben nach unten (dorsoventral) ist der Zellleib abgeplattet. Auf der Unterseite (Ventralseite) sitzt ein grubenförmiges Mundfeld. Diese grubenförmige Einsenkung zieht sich – in etwa halber Körperlänge – asymmetrisch vom linken Vorderende bis zur Körpermitte. Am Ende geht die Einsenkung in den Mundtrichter (Vestibulum) über, an dessen spitzem Ende eine Mundöffnung (der Zellmund) liegt. Vom Zellmund aus zieht sich eine röhrenartige Buccalhöhle, die in Aufsicht meist s-förmig erscheint und mit spitzem Ende blind endet, nach innen. Die Anordnung und Morphologie der Cilien in der Buccalhöhle ist wichtig für die genaue Bestimmung der Arten. Die Zelloberfläche wird durch eine steife, aber dabei elastische Hülle, Pellicula genannt, verstärkt. Diese ist auf der Oberseite in langgestreckt sechseckige (hexagonale) Felder gegliedert. Aus jedem der Felder entspringt eine, selten auch zwei Zilien („Wimpern“), die reihig auf der gesamten Oberfläche sitzen. Bei einigen Arten sind sie am Hinterende büschelartig verlängert. Unter der Pellikula liegt ein dünner, Rinde (Cortex) genannter Bereich des Zellplasmas, in dem die Trichocysten sitzen (vgl. unten im Kap. Verteidigung). Das darunter liegende Endoplasma wirkt dünnflüssiger und enthält zahlreiche Organellen, oft auch Kristalle, die sich mit der Plasmaströmung im Zellkörper mitbewegen. Normalerweise sind zwei kontraktile Vakuolen vorhanden. Diese pulsieren alternierend (abwechselnd) und geben ihr Inneres durch eine Pore nach außen ab. Im typischen Fall sind zwei Arten von Zellkernen vorhanden: ein einzelner, großer Makronukleus von elliptischer bis nierenförmiger Gestalt, ein wenig seitlich in der Körpermitte gelegen, und eine je nach Art unterschiedliche Anzahl viel kleinerer Mikronuklei, die meist in unmittelbarer Nachbarschaft liegen.[3]
Die größten Arten zählt man zu den „Riesen“ unter den Einzellern, da sie schon mit bloßem Auge als kleine, weiße Pünktchen in einem Wassertropfen zu erkennen sind.
Fortbewegung mit Hilfe von Wimpern
Pantoffeltierchen fallen unter dem Mikroskop durch schnelles Umherschwimmen auf. Sie sind außen von vielen (etwa 10.000) Wimpern (Zilien) umgeben, die der Fortbewegung dienen. Durch das rhythmische Krümmen und Wiederaufrichten der Wimpern ziehen sich Schlagwellen über den Körper des Pantoffeltierchens. Durch die spiralförmige Anordnung dieser Wimpern wird das Pantoffeltierchen um seine Längsachse gedreht, wodurch die körperbedingte seitliche Bahnabweichung nicht zur Kreisbewegung, sondern zur Schraubenbahn wird. Es hat für einen Einzeller eine recht große Geschwindigkeit von 1 bis 1,4 mm/s. Hindernisse oder Engpässe passiert das Pantoffeltierchen mit Leichtigkeit, da es sich aufgrund einer elastischen Zellmembran (genauer: Pellicula) mühelos an ihnen vorbei- oder hindurchbewegen kann. Drehungen in jede Richtung sind möglich, bei Schreckreaktionen sogar durch Umkehr des Cilienschlages ein plötzliches Rückwärtsschwimmen.
Nahrungsaufnahme, Verdauung und Ausscheidung
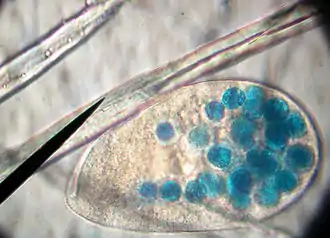
Das Pantoffeltierchen nimmt seine Beute und Nahrung durch seinen „chemischen“ Sinn (Chemorezeptoren) und durch Tastreize (andere molekulare Rezeptoren) wahr. Es ernährt sich vorwiegend von Bakterien, die durch Wimpernschläge zum Mundfeld befördert werden. Die Wimpern wirken also auch bei der Nahrungsaufnahme mit, indem sie Nahrungspartikel heranstrudeln. Mit Hilfe der Mundfeldbewimperung gelangen die Bakterien über das Mundfeld zum Zellmund, wo sie anschließend im Zellschlund in eine Nahrungsvakuole importiert werden. Dieser Vorgang heißt Endocytose.
Zu dieser Nahrungsvakuole führen zahlreiche bandartige Strukturen, mithilfe derer viele Membranbläschen herantransportiert werden, die die Nahrungsvakuole vergrößern. Hat sie eine bestimmte Größe erreicht, schnürt sie sich ins Zellinnere ab. Überschüssiges Wasser wird der Nahrungsvakuole entzogen.
Zunächst gelangen Acidosomen in die Nahrungsvakuole und senken den pH-Wert auf 1,2 ab. Über Lysosomen gelangen Verdauungsenzyme ins Bläschen, das von nun an Verdauungsvakuole heißt. Während die Verdauung abläuft und die Bakterien zersetzt werden, wird die Verdauungsvakuole auf einer ovalförmigen Bahn durch die Zelle transportiert. Dies nennt man Cyclose.
Die verwertbaren Nahrungsstoffe werden resorbiert und die unverdaulichen Substanzen über die Cytopyge (den so genannten Zellafter) ausgeschieden. Damit dies geschehen kann, muss das Verdauungsbläschen als Kontaktvakuole am Zellafter Kontakt mit der Zellmembran aufnehmen. Dieser Vorgang heißt Exozytose. Da es nahezu ständig Nahrung in sich hineinstrudelt, kann ein Pantoffeltierchen innerhalb weniger Stunden seine Körpermasse verdoppeln.
In die Zelle eingedrungenes überschüssiges Wasser wird mit Hilfe von sternförmig angeordneten Zuführungskanälen in die Sammelblasen von zwei pulsierenden Bläschen (kontraktilen Vakuolen) geleitet und aus diesen über einen Exkretionsporus aktiv ausgeschieden.
Vermehrung
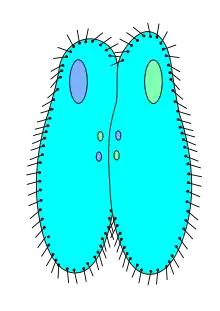
Pantoffeltierchen vermehren sich normalerweise ungeschlechtlich durch Querteilung in zwei Tochterzellen. Das Pantoffeltierchen zieht sich in die Länge und das Mundfeld teilt sich. Es bildet sich jeweils eine weitere pulsierende Vakuole aus. Der Mikronukleus und der Makronukleus verdoppeln sich. Das Pantoffeltierchen schnürt nun noch den Zellleib so durch, dass jedes der beiden auf diese Weise neu entstehenden Individuen einen Kleinkern, einen Großkern, zwei pulsierende Vakuolen und ein Mundfeld enthält. Das Pantoffeltierchen teilt sich unter günstigen Bedingungen bis zu siebenmal pro Tag.[4]
Gelegentlich kommt es auch zu geschlechtlichen Vorgängen, die man Konjugation nennt und bei denen die Paramecien mit anderen Individuen der gleichen Art Erbinformationen austauschen. Zwei Pantoffeltierchen legen sich dazu an den Mundfeldern aneinander. Die Zellmembranen verschmelzen in diesem Bereich, die Wimpern verschwinden. Die Großkerne lösen sich allmählich auf. Die Kleinkerne teilen sich durch Reduktionsteilung in vier haploide Tochterkerne. Von diesen vier sterben drei ab, und einer teilt sich zu zwei haploiden Kernen. Jeweils ein Tochterkern wandert in das andere Pantoffeltierchen, um mit dem dort verbliebenen Tochterkern zu einem neuen diploiden Kern zu verschmelzen. In jedem Individuum teilt sich der durch Verschmelzung entstandene neue Kern in zwei Tochterkerne. Der eine ist der neue Kleinkern, der andere entwickelt sich (unter mehrfacher Verdoppelung und vollständiger Neuorganisation der Chromosomen) zu einem Großkern. Nun trennen sich die Konjugationspartner wieder, Wimpern und Mundfelder werden im fehlenden Abschnitt ergänzt.
Die Konjugation wird unter anderem durch jahreszeitliche Änderungen oder sich verschlechternde Umweltbedingungen stimuliert.
Verhalten
Das Pantoffeltierchen reagiert sehr stark auf Reize der Umgebung (Berührung, Temperatur, chemische Reize, Belichtung). In geschlossenen Glasröhren schwimmen sie immer in Richtung Wasseroberfläche, obwohl sie schwerer als Wasser sind. Sie reagieren auf Schwerkraft (Gravitaxis). Pantoffeltierchen nehmen chemische und thermische Reize nur mit dem vorderen Teil des Körpers wahr. Trifft ein Pantoffeltierchen auf ein Hindernis, schwimmt es durch Umkehrung des Wimpernschlages ein Stück zurück und vollführt eine leichte Drehung. Dann schwimmt es wieder nach vorn. Trifft es wieder auf das Hindernis, so probiert es das Pantoffeltierchen solange mit dieser Methode, bis es an dem Hindernis vorbeikommt. Das Pantoffeltierchen arbeitet koordiniert, da nicht nur die betroffene Stelle, sondern das ganze Lebewesen die Reaktion ausführt.
Feinde
Zu den Feinden der Pantoffeltierchen gehören Amöben und Einzeller der Gattung Didinium. Letztere gehören ebenfalls zu den Ciliaten. Didinium ergreift mit Hilfe einer nasenartigen Vorstülpung das Pantoffeltierchen und verleibt es sich anschließend ein. Amöben und Sonnentierchen umhüllen das Pantoffeltierchen mit Scheinfüßchen und verdauen es anschließend in einer Nahrungsvakuole.
Gegen Angreifer versucht sich das Pantoffeltierchen mit Hilfe der Trichocysten zu wehren. Es handelt sich dabei um stäbchenförmige Gebilde enthaltende Haarbläschen, deren Spitzen kalzifiziert sind, die direkt unterhalb der Zellmembran liegen und bei Gefahr lange, klebrige Proteinfäden (Eiweißfäden) ausschleudern. Sobald ein Angreifer ein Pantoffeltierchen berührt, verlängern sich diese Gebilde explosionsartig und schießen die Proteinfäden ins Wasser. Ein ganzes Büschel der Proteinfäden kann dem Pantoffeltierchen helfen, sich Feinde vom Leib zu halten. In den abgeschossenen Proteinfäden können sich manche Fressfeinde verfangen und letztendlich auch absterben. Ausgestoßene Trichocysten werden durch neue ersetzt, die in Vesikeln im Cytoplasma entstehen.
Nutzung durch den Menschen
Pantoffeltierchen werden als Indikator für die Gewässerverschmutzung genutzt.[5] Außerdem werden sie auch als Fischfutter verwendet.[6]
Kerndimorphismus und Genom
Wie viele Ciliaten sind Pantoffeltierchen kerndimorph, d. h. der Zellkern kommt in zwei verschiedenen Formen vor. Ein großer Zellkern, Makronukleus genannt, ist somatisch aktiv, in ihm werden Gene abgelesen und transkribiert. Ein oder mehrere kleine Zellkerne oder Mikronuklei sind in dieser Zeit in Ruhestadien. Ihre Gene werden nicht transkribiert. Bei der normalen, ungeschlechtlichen Zellteilung werden Makro- und Mikronukleus wie üblich separat, über eine Mitose, verdoppelt und auf die Tochterzellen verteilt. Bei der geschlechtlichen Fortpflanzung (einer Meiose), entweder über ein Konjugation zwischen zwei Zellen oder über Autogamie, wird der Makronukleus nicht verdoppelt, sondern zerstört. Anschließend wird aus einem Mikronukleus ein neuer Makronukleus gebildet.
Der genetische Gehalt in Makro- und Mikronukleus ist identisch, d. h. in ihnen sind im Prinzip dieselben Gene vorhanden. Allerdings wird das Genom bei der Bildung des Makronukleus vollkommen umgebaut. Alle Chromosomen werden mehr als achthundert Mal verdoppelt. Anschließend werden aus zahlreichen kurzen Sequenzstücken daraus funktionale Gene zusammengesetzt. Repetitive DNA und Transposonen werden ausgeschnitten. Durch den vollständigen Umbau ist der Makronukleus anschließend homozygot.
Da alle Kopien des Makronukleus bei einer geschlechtlichen Fortpflanzung verloren gehen, werden letztlich nur Mutationen im Mikronukleus dauerhaft vererbt. Dadurch besitzen Pantoffeltierchen eine äußerst geringe Mutationsrate.[7] Vielleicht erleichtert durch den eigenartigen Fortpflanzungsmodus wurden aber im Genom von Paramecium (untersucht wurde die Art Paramecium tetraurelia aus dem aurelia-Artkomplex) mit fast 40.000 eine ungewöhnlich hohe Anzahl von Genen gefunden. Dieses Pantoffeltierchen besitzt damit, überschlägig, etwa die doppelte Anzahl an Genen wie der Mensch, Homo sapiens. Die ungewöhnlich hohe Genanzahl wird auf mehrere Runden (mindestens drei) von Verdoppelung des gesamten Genoms zurückgeführt, bei der zahlreiche paraloge Gene entstehen.[8]
Arten
Die Gattung Paramecium gehört zu den Peniculia[9], einer Gruppe der Wimpertierchen, die je nach Autoren im Rang einer Unterordnung, einer Ordnung oder einer Unterklasse eingeordnet wird. Innerhalb der Gruppe steht sie nach genetischen Daten relativ isoliert mit hoher Distanz zu anderen, sie wird deshalb formal als einzige Gattung in die damit monotypische Familie Parameciidae gestellt, die meisten Bearbeiter verzichten aber ganz auf die Angabe einer Familie. Mögliche Schwestergruppe könnte die überwiegend marine Gattung Frontonia sein.[10][11]
Die Gattung wird nach morphologischen und genetischen Daten in vier[12], nach neueren Ergebnissen sogar fünf[13] Untergattungen geteilt. Die Abgrenzung und Anzahl der Arten anzugeben ist schwierig. Es werden etwas weniger als 20 Morphospezies unterschieden, die sich nach anatomischen Merkmalen unter dem Mikroskop unterscheiden lassen. Die Arten können teilweise anhand der generellen Körpergestalt[12], von langgestreckt „zigarrenförmig“ bis „pantoffelförmig“ mit in Aufsicht rundlicher Gestalt mit abgerundet schief abgeschnitten wirkendem Vorderende, gruppenweise unterschieden werden. Zwei Arten sind durch endosymbiontische Algen grün gefärbt. Eine genaue Bestimmung[14] beruht auf der Gestalt und Anordnung der Cilien, der Zahl der Mikronuklei und verschiedener Vakuloen. Innerhalb der Morphospezies werden aber zusätzlich Kryptospezies unterschieden, das sind Arten, die morphologisch gleich (oder fast gleich) aussehen, sich aber anhand von genetischen, biologischen und ökologischen Daten unterscheiden lassen. Bekannt ist insbesondere der „Paramecium aurelia“-Artenkomplex.[14] Was sich morphologisch als eine Morphospezies darstellt, konnte nach dem Paarungsverhalten in fünfzehn „Stämme“ oder „Linien“ geschieden werden, bei denen jeweils zwei Paarungstypen (o, nach englisch odd, ungleich und e, nach englisch even, gleich) existieren. Jeder dieser Stämme akzeptiert bei der Konjugation (im Wesentlichen) nur Individuen desselben Stamms, wobei sich ausschließlich o und e miteinander paaren – dadurch ist es im Labor möglich, durch entsprechende Auslese im Kulturmedium die Konjugation ganz zu unterdrücken. Die Stämme sind später von ihrem Entdecker als Arten beschrieben worden, wobei er sie im Namen einfach (lateinisch) durchnummerierte: Paramecium primaurelia, Paramecium biaurelia, Paramecium triaurelia usw. Da diese in der Reproduktion gegeneinander isoliert sind, ist ein wesentliches Kriterium für die Abgrenzung einer Art, trotz der fehlenden morphologischen Merkmale, erfüllt. Genetisch[15] konnte die Eigenständigkeit der kryptischen Arten des aurelia-Artenkomplexes im Wesentlichen bestätigt werden. Unerwarteterweise existierten aber einige Stämme, die nach der genetischen Verwandtschaft zu einer bestimmten Linie gehörten, sich aber mit Angehörigen eines „falschen“ Stammes paarten; hier wird ein späterer Wechsel durch eine nachträgliche Mutation angenommen. Auch andere Morphospezies wie Paramecium multimicronucleatum erwiesen sich als aus kryptischen Arten zusammengesetzt,[16] sind aber schlechter erforscht.
In der folgenden Aufstellung[17] sind nur die Morphospezies enthalten.
Untergattung Paramecium.
- Paramecium caudatum Ehrenberg, 1833. Typusart der Gattung.
- Paramecium aurelia – Komplex (mit 15 kryptischen Arten)
- Paramecium multimicronucleatum Powers & Mitchel, 1910
- Paramecium jenningsi Diller & Earl, 1958
- Paramecium wichtermani Mohammed & Nashed, 1968
- Paramecium africanum Dragesco, 1970
- Paramecium jankowskii Dragesco, 1972
- Paramecium ugandae Dragesco, 1972
- Paramecium schewiakoffi Fokin et al., 2001
- Paramecium buetschlii Krenek, Berendonk & Fokin, 2015
Untergattung Chloroparamecium
- Paramecium bursaria Focke, 1836
Untergattung Helianter
- Paramecium putrinum Claparéde & Lachman, 1858
- Paramecium duboscqui Chatton and Brachon, 1933
Untergattung Cypriostomum
- Paramecium calkinsi Woodruff, 1921
- Paramecium polycaryum Woodruff & Spencer, 1923
- Paramecium nephridiatum Gelei, 1925
- Paramecium woodruffi Wenrich, 1928
- Paramecium pseudotrichium Dragesco, 1970
Untergattung Viridoparamecium
- Paramecium chlorelligerum Kahl, 1935
Siehe auch
Literatur
- Heinz Streble, Dieter Krauter: Das Leben im Wassertropfen. Mikroflora und Mikrofauna des Süßwassers. Ein Bestimmungsbuch. 10. Auflage. Kosmos, Stuttgart 2006, ISBN 3-440-10807-4.
- H.-D. Görtz (Hrsg.): Paramecium. Springer Verlag, Berlin 1988.
- R. Wichterman: The Biology of Paramecium. 2. Aufl., Plenum Press, New York 1986.
Einzelnachweise
- Alexei O. Smurov & Sergei I. Fokin (1999): Resistance of Paramecium Species (Ciliophora, Peniculia) to Salinity of Environment. Protistology 1: 43-53.
- R. Wichterman: The Biology of Paramecium. Springer, 2012, ISBN 978-1-4757-0372-6, S. 345–355.
- R. Wichterman: The Biology of Paramecium. Springer, 2012, ISBN 978-1-4757-0372-6, S. 6 bis 8.
- H. Linder: Herman Linder Biologie.
- Pantoffeltierchen Einzeller des Jahres – Deutsche Gesellschaft für Protozoologie kürt Paramecium auf scinexx.de
- Futter für Aquarienfische - Haltung und Zucht der Pantoffeltierchen auf aqua-spider.de
- Way Sung, Abraham E. Tucker, Thomas G. Doak, Eunjin Choi, W. Kelley Thomas, Michael Lynch (2012): Extraordinary genome stability in the ciliate Paramecium tetraurelia. PNAS Proceedings of the National Academy of Sciences USA 109 (47): 19339–19344, doi:10.1073/pnas.1210663109.
- Jean-Marc Aury et al. (2006): Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature 444: 171–178, doi:10.1038/nature05230.
- Sina M. Adl et al. (2012): The Revised Classification of Eukaryotes. Journal of Eukaryotic Microbiology 59(5): 429–493. doi:10.1111/j.1550-7408.2012.00644.x.
- Michaela C. Strüder-Kypke, André-Denis G. Wright, Sergei I. Fokin, Denis H. Lynn (2000): Phylogenetic Relationships of the Subclass Peniculia (Oligohymenophorea, Ciliophora) Inferred from Small Subunit rRNA Gene Sequences. Journal of Eukaryotic Microbiology 47(4): 419-429. doi:10.1111/j.1550-7408.2000.tb00069.x.
- Yan Zhao, Zhenzhen Yi, Eleni Gentekaki, Aibin Zhan, Saleh A. Al-Farraj, Weibo Song (2015): Utility of combining morphological characters, nuclear and 4 mitochondrial genes: An attempt to resolve the conflicts of species 5 identification for ciliated protists. Molecular Phylogenetics and Evolution 94:718–729. doi:10.1016/j.ympev.2015.10.017.
- Sergei I. Fokin (2011): Paramecium genus: biodiversity, some morphological features and the key to the main morphospecies discrimination. Protistology 6(4): 227–235.
- Martin Kreutz, Thorsten Stoeck, Wilhelm Foissner (2012): Morphological and Molecular Characterization of Paramecium (Viridoparamecium nov. subgen.) chlorelligerum Kahl, 1935 (Ciliophora). Journal of Eukaryotic Microbiology 59(6): 548–563 doi:10.1111/j.1550-7408.2012.00638.x.
- R. Wichterman: The Biology of Paramecium. Springer, 2012, ISBN 978-1-4757-0372-6.
- Francesco Catania, François Wurmser, Alexey A. Potekhin, Ewa Przyboś, Michael Lynch (2009): Genetic Diversity in the Paramecium aurelia Species Complex. Molecular Biology and Evolution 26(2): 421–431, doi:10.1093/molbev/msn266.
- Sebastian Tarcz, Alexey Potekhin, Maria Rautian, Ewa Przyboś (2012): Variation in ribosomal and mitochondrial DNA sequences demonstrates the existence of intraspecific groups in Paramecium multimicronucleatum (Ciliophora, Oligohymenophorea). Molecular Phylogenetics and Evolution 63(2):500-509, doi:10.1016/j.ympev.2012.01.024.
- basierend auf S.I.Fokin: Paramecium – die Gattung. Deutsche Gesellschaft für Protozoologie: Einzeller des Jahres 2007, mit Ergänzungen.
Weblinks
- Umfangreiche Informationen auf der Website der Deutschen Gesellschaft für Protozoologie zum „Einzeller des Jahres“ 2007
- Video: Paramecium unter dem Mikroskop
- Videos zu Paramecium herausgegeben vom Institut für den Wissenschaftlichen Film.
Pantoffeltierchen (2007) | Tetrahymena (2009) | Augentierchen (2010) | Dictyostelium (2011) | Acanthamoeba (2012) | Actinophrys (2013) | Trompetentierchen (2014) | Vampiramöben (2015) | Trichomonas vaginalis (2016) | Diaphanoeca grandis (2017) | Dictyocysta mitra (2018) | Nuclearia (2019) | Dinophysis acuta (2020) | Physarum polycephalum (2021) | Blastocystis (2022)