Schalenweichtiere
Schalenweichtiere (Conchifera) sind alle Weichtiere mit einem primären, flächig ein- oder zweiklappigen oder gehäuseartigen Außenskelett auf der (ursprünglichen) Rückenseite des Körpers. Neben diesem Merkmal wird die Gruppe vor allem begründet durch den Besitz von Kiefern und paarigen Statocysten sowie das Fehlen einer dünnen, überwiegend chitinösen Schutzschicht (Cuticula) auf der Rückenseite des Körpers. Die Conchiferen stellen den weit überwiegenden Teil aller rezenten und fossil bekannten Weichtiere und weisen eine große Formenvielfalt auf.
| Schalenweichtiere | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Weinbergschnecke (Helix pomatia) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Conchifera | ||||||||||||
| Gegenbaur, 1878 | ||||||||||||
| Klassen | ||||||||||||
|
Aufbau, Wachstum und Funktion der Schale
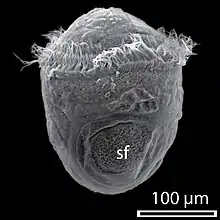
Beim Aufbau des geschlossenen Außenskeletts der Conchifera, auch Schale genannt, werden zwei Schichten unterschieden: Das Periostracum, die äußere, sehr dünne Schicht, auch Schalenhäutchen genannt, bestehend aus organischer Substanz (Conchiolin), und die dicke innere Schicht, bestehend aus Calciumcarbonatkristallen, die in eine Conchiolin-Matrix eingebettet sind. Die innere Schicht gliedert sich bei zahlreichen Weichtieren (Perlbooten, einige Schnecken, einige Muscheln) weiter in das Ostracum (Prismenschicht), die äußere, unmittelbar unterhalb des Periostracums befindliche Calciumcarbonat-Schicht, mit relativ großen, senkrecht zur Schalenoberfläche orientierten Prismen aus Aragonit oder Calcit, und das Hypostracum (Perlmuttschicht), die innere Calciumcarbonat-Schicht, mit sehr kleinen, tafeligen Aragonit-Prismen – das entsprechende, irisierende mineralisch-organische Verbundmaterial wird Perlmutt genannt. Bei anderen Vertretern ist die Calciumcarbonat-Schicht weitgehend homogen aufgebaut, wobei prismatische Strukturen und, bei reinen Aragonitschalern, Kreuzlamelle (winklig zueinander angeordnete, mit einer dünnen „Conchiolin“-Schicht umhüllte Aragonitnadeln) sowie Perlmutt unterschieden werden. Neben Calcit und Aragonit können Molluskenschalen auch das Calciumcarbonatmineral Vaterit in geringen Mengen enthalten.[1][2]
Die Schalen der Conchiferen haben in erster Linie eine Schutzfunktion und weisen eine enorme Formenvielfalt auf. Sie können einfache, mützen- oder napfartige „Panzer“ bilden oder aber röhrenförmige, nicht selten spiralig gewundene Gehäuse, in die sich das Tier bei Gefahr vollständig zurückziehen kann (siehe z. B. → Schneckenhaus). Viele gehäusetragende Conchiferen bilden deckelartige Strukturen aus, mit denen sie die Öffnung des Gehäuses (Apertur) verschließen können, wodurch sich die Schutzwirkung erhöht (beispielsweise die Kopfkappe der Perlboote oder die Opercula bestimmter Schnecken). Muscheln haben zwar keine „echten“ Gehäuse, aber die aus zwei Klappen bestehende Muschelschale erfüllt die gleiche Funktion. Die Ausbildung der Schale ist eng mit der Lebensweise ihrer Besitzer verknüpft. So brauchen Napfschnecken keine „echten“ Gehäuse, da sie auf felsigem Untergrund leben und zum Schutz die flachkegelige Schale an den Fels pressen. Die allgemein freischwimmenden Perlboote sowie ihre engeren und weniger engen fossilen Verwandten (vgl. → Nautiloideen, → Ammoniten) haben ein gekammertes Gehäuse, dass größtenteils mit einem luftähnlichen Gasgemisch gefüllt ist und als Auftriebskörper fungiert (vgl. → Phragmokon).
Die Schale wird bereits früh in der Individualentwicklung angelegt. Typischerweise verdicken sich zunächst die Zellen einer dem Urmund (Blastoporus) gegenüberliegenden Region des Ektoderms, des sogenannten Schalenfeldes, und anschließend stülpt sich der zentrale Teil des Schalenfeldes ins Blastocoel ein und bildet die Schalendrüse. Oft stülpt sich ein Teil der Schalendrüse anschließend knospenartig wieder aus. Schalendrüse bzw. Schalenfeld scheiden entweder erst nach der Ausstülpung oder bereits vorher die Embryonalschale, das uhrglasförmige Protoconch I, ab. Durch Wachstum des Embryos bzw. des auf den Schlupf folgenden Larvenstadiums vergrößert sich das Schalenfeld zum larvalen Mantelepithel und das Protoconch I wird durch randliche Anlagerung von Schalenmaterial zur Larvalschale, dem Protoconch II, erweitert. Ein ausgeprägtes Protoconch II weisen vor allem Schalen von Conchiferen mit relativ langlebiger, planktotropher Veliger-Larve auf. Bei Muscheln wird das Protoconch zweiklappig angelegt und daher auch als Prodissoconch bezeichnet. Auch im Adultstadium erfolgt das Flächenwachstum der Schale ausschließlich randlich. Der während des Adultstadiums gebildete Teil der Schale wird Teleoconch genannt. Beim randlichen Schalenwachstum scheidet die rinnenartig in den Mantelrand eingesenkte Periostracaldrüse das Periostracum ab und Zellen des randlichen Mantelepithels sezernieren die äußere Calciumcarbonatschicht. Da das Flächenwachstum nicht ganz kontinuierlich, sondern in Schüben erfolgt, zeichnen sich Molluskenschalen durch typische Anwachsstreifen an ihren Außenflächen auf. Zellen des Mantelepithels, die weiter entfernt vom Mantelrand liegen, scheiden das Calciumcarbonat der inneren Schalenschichten ab und sorgen somit für weiteres Dickenwachstum der Schale. Daher zeigen die Innenflächen keine Anwachsstreifen. Tintenfische haben ihre Schale ins Innere des Körpers verlegt oder stark reduziert. Bei ihnen wird die embryonale Schalendrüse von aufgefaltetem, umliegendem Ektoderm überwachsen und schließlich abgeschnürt.[3]
Systematik
Zu den Schalenweichtieren gehören die Einschaler (Monoplacophora), die Schnecken (Gastropoda), die Kopffüßer (Cephalopoda) sowie die Kahnfüßer (Scaphopoda) und die Muscheln (Bivalvia). Außerdem werden die ausgestorbenen Schnabelschaler (Rostroconchia) und die Helcionelliden (Helcionellida) zu den Schalenweichtieren gerechnet.
Die traditionelle Einteilung der Schalenweichtiere in Gekrümmtschaler (Cyrtosoma) und Gestrecktschaler (Diasoma), unter anderem anhand unterschiedlicher Tendenzen bei der Schalenausbildung, ist heute nicht mehr aktuell.
Als Schwestergruppe werden den Schalenweichtieren die Stachelweichtiere (Aculifera) gegenübergestellt. Das untenstehende Kladogramm stützt sich auf die im Jahr 2011 in der Zeitschrift Nature veröffentlichten Ergebnisse zweier molekulargenetischer Verwandtschaftsanalysen der Mollusken.[4][5]
| Weichtiere |
| |||||||||||||||||||||||||||||||||||||||||||||
Literatur
- Gerhard Haszprunar, Klaus-Jürgen Götting: Mollusca, Weichtiere. S. 305–362 in: Westheide, Rieger (Hrsg.): Spezielle Zoologie Teil 1: Einzeller und Wirbellose Tiere. 2. Auflage. Spektrum Akademischer Verlag (Elsevier), München 2007, ISBN 978-3-8274-1575-2
Einzelnachweise
- Nicole Spann, Elizabeth M. Harper, David C. Aldridge: The unusual mineral vaterite in shells of the freshwater bivalve Corbicula fluminea from the UK. Naturwissenschaften. Bd. 97, Nr. 8, 2010, S. 743–751, doi:10.1007/s00114-010-0692-9 (alternativer Volltextzugriff: carbonateresearch.com).
- Gernot Nehrke, Harald Poigner, Dorothee Wilhelms-Dick, Thomas Brey, Doris Abele: Coexistence of three calcium carbonate polymorphs in the shell of the Antarctic clam Laternula elliptica. Geochemistry, Geophysics, Geosystems. Bd. 13, Nr. 5, 2012, doi:10.1029/2011GC003996 (Open Access).
- Absatz i.e.L. nach Andreas Wanninger, Tim Wollesen: Mollusca. S. 103–154 in: Andreas Wanninger (Hrsg.): Evolutionary Developmental Biology of Invertebrates 2: Lophotrochozoa (Spiralia). Springer, 2015, ISBN 978-3-7091-1870-2, S. 131 ff.
- Kevin M. Kocot, Johanna T. Cannon, Christiane Todt, Mathew R. Citarella, Andrea B. Kohn, Achim Meyer, Scott R. Santos, Christoffer Schander, Leonid L. Moroz, Bernhard Lieb, Kenneth M. Halanych: Phylogenomics reveals deep molluscan relationships. Nature. Bd. 477, 2011 S. 452–456, doi:10.1038/nature10382
- Stephen A. Smith, Nerida G. Wilson, Freya E. Goetz, Caitlin Feehery, Sónia C. S. Andrade, Greg W. Rouse, Gonzalo Giribet, Casey W. Dunn: Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature. Bd. 480, 2011, S. 364–367, doi:10.1038/nature10526