Pazifischer Maulwurf
Der Pazifische Maulwurf (Scapanus orarius) ist eine Säugetierart aus der Gattung der Westamerikanischen Maulwürfe innerhalb der Maulwürfe (Talpidae). Er kommt entlang der pazifischen Küste Nordamerikas vom Südwesten von British Columbia bis zum Nordwesten von Kalifornien vor. Teilweise tritt er hier gemeinsam mit dem Townsend-Maulwurf auf, nutzt abweichend von diesem aber häufiger waldreiche Habitate. Allerdings ist er auch in offeneren Gebieten anzutreffen. Es handelt sich um einen mittelgroßen Vertreter der Maulwürfe, der wie alle grabenden Angehörigen der Gruppe typische Anpassungen dafür besitzt. Dazu gehören ein walzenförmiger Körper, ein kurzer Hals und grabschaufelartige Vordergliedmaßen. Das Fell ist dunkelgrau gefärbt.
| Pazifischer Maulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpeg.webp)
Pazifischer Maulwurf (Scapanus orarius) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Scapanus orarius | ||||||||||||
| True, 1896 |
Die Lebensweise ist in Teilen gut erforscht. Die Tiere legen unterirdische Tunnel und Gänge an, die weit verzweigt sind und in mehreren Ebenen verlaufen. Äußerlich sichtbar sind die Gangsysteme durch Maulwurfshügel, deren Häufigkeit mit der Dichte der Beutetiere korreliert. Als Hauptnahrung dienen Regenwürmer, daneben frisst der Pazifische Maulwurf auch eine Vielzahl von Insekten und teilweise auch Pflanzen. Der Nachwuchs kommt einmal jährlich zur Welt und wächst in einem gepolsterten Nest auf. Die Art wurde im Jahr 1896 wissenschaftlich eingeführt. Es sind zwei Unterarten anerkannt. Der Bestand wird als nicht bedroht eingestuft. Das häufigere Auftreten in waldreicheren Landschaften führt zu weniger Konfliktpotential mit dem Menschen. Fossilfunde des Pazifischen Maulwurfs sind nicht bekannt.
Merkmale
Habitus
Der Pazifische Maulwurf ist ein mittelgroßer Vertreter der Westamerikanischen Maulwürfe und besitzt eine Kopf-Rumpf-Länge von 10,6 bis 16,8 cm. Der Schwanz wird 2,1 bis 4,6 cm lang und macht damit weniger als ein Viertel der Gesamtlänge aus. Das Gewicht variiert von 61 bis 91 g. Männchen sind in der Regel größer als Weibchen, womit ein auffallender Geschlechtsdimorphismus vorliegt. Zudem sind die Individuen im östlichen Verbreitungsgebiet größer als die im westlichen. Äußerlich ähnelt der Pazifische Maulwurf den anderen Westamerikanischen Maulwürfen und zeigt wie diese eine deutliche Anpassung an eine grabende Lebensweise. Der Körper ist dadurch walzenförmig, der Hals kurz und die Vordergliedmaßen sind zu kräftigen Grabwerkzeugen umgestaltet. Das samtige Fell weist einen dunkelgrauen Farbton auf. Im Winter tendiert es eher zu schwarzen, im Sommer zu braunen Farbtönen. Eine Unterwolle fehlt, die Haare sind etwa gleich lang. Der Schwanz zeigt sich nur spärlich behaart, er ist fleischig, an der Basis nur wenig eingezogen und spitzt sich zum Ende hin zu. Vom ähnlichen Kalifornischen Maulwurf (Scapanus latimanus) unterscheidet sich der Pazifische Maulwurf dadurch, dass der nackte Bereich der spitzen Schnauze bis zum vorderen Rand der Nase reicht. Die Nasenlöcher öffnen sich sichelartig geschlitzt und zeigen nach oben. Generell sind die Augen klein und äußere Ohrmuscheln nicht erkennbar. Die Hinterfußlänge beträgt 1,5 bis 2,4 cm. Weibchen besitzen vier Paare an Zitzen, zwei in der Brust- und je eines in der Bauch- und Leistengegend. Die Zitzen sind stärker nach außen positioniert als bei anderen Vertretern der Gattung.[1][2][3][4]
Schädel- und Gebissmerkmale
Die Schädellänge reicht von 32,1 bis 34,7 mm, die Breite an den Warzenfortsätzen misst 15,4 bis 16,8 mm. Im Bereich der Orbita ist er 7,4 bis 8,4 mm weit, die Höhe des Schädels beträgt 8,8 bis 10,1 mm. Er zeichnet sich durch einen breiten Hirnschädel und ein schmales sowie lang ausgezogenes Rostrum aus. Dessen Breite auf Höhe der Molaren variiert von 8,0 bis 9,0 mm, auf Höhe der Eckzähne von 3,5 bis 4,2 mm. An der Stirnlinie verläuft er flach. Der Mittelkieferknochen überragt nach vorn das Nasenbein. Die Nasenöffnungen richten sich nach vorn. Die Paukenblasen auf der Schädelunterseite sind vollständig und abgeflacht. Am Unterkiefer hebt sich der vergleichsweise große horizontale Knochenkörper hervor. Das Gebiss ist vollständig und setzt sich aus 44 Zähnen mit folgender Zahnformel zusammen: . Wie bei allen Neuweltmaulwürfen ist der obere vordere Schneidezahn vergrößert, aber nicht so stark wie beim Kalifornischen Maulwurf. Die nachfolgenden Zähne bis zum dritten Prämolar ähneln sich weitgehend und sind einspitzig (unicuspid), auch wenn am letzteren ein zusätzlich kleines Höckerchen auftreten kann. Die beiden vorderen Molaren der oberen und unteren Zahnreihe heben sich durch ihre auffallende Größe hervor, der letzte wiederum ist deutlich kleiner. Insgesamt stehen die Zähne gleichmäßig verteilt und sind nicht so dicht gedrängt wie beim Kalifornischen Maulwurf. Die obere Zahnreihe ist 9,5 bis 11,2 mm lang, die untere Prämolaren-Molaren-Reihe erstreckt sich über 10,3 bis 10,9 mm.[1][2][3][4]

Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 34.[4]
Verbreitung und Lebensraum
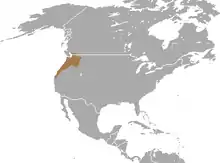
Der Pazifische Maulwurf kommt im westlichen Nordamerika vor. Sein Verbreitungsgebiet erstreckt sich vom Südwesten der kanadischen Provinz British Columbia südwärts über einen Großteil des US-Bundesstaats Washington sowie den westlichen und nördlichen Teil von Oregon bis hin zu den nördlichen Küstenregionen von Kalifornien. Nach Osten hin schließt es die westlichsten Gebiete von Idaho mit ein. Die Nordgrenze wird etwa auf Höhe des Fraser River zwischen Vancouver und Chilliwack erreicht,[5] die Südgrenze liegt bei Mendocino. Eine Population kommt auf Vashon Island im Puget Sound von Washington vor. In Oregon ist ein Großteil des Willamette Valleys sowie der südliche Landesteil nicht besiedelt, in letzterem tritt der Kalifornische Maulwurf auf.[2][3][4]
Die Tiere sind an eine Vielzahl von Landschaften angepasst. In British Columbia wurden sie im Zusammenhang mit verschiedenen Bodentypen dokumentiert, so mit eiszeitlichem Geschiebemergel mit angeschwemmten Tonen, mit Sanden und Kiesen sowie mit Flussablagerungen.[5] Des Weiteren ist die Art auf Ackerflächen und Wiesen beziehungsweise in Graslandschaften mit Artemisia-Bestand ebenso wie in Nadel- und Laubwäldern oder in Dünengebieten anzutreffen. Laubwälder bestehen häufig aus Erlen und Hartriegeln, Nadelwälder aus Gelb-Kiefern, Douglasien, Küstenmammutbäumen, Hemlocktannen und Fichten. Die direkten Küstenlandschaften, küstennahe Seen oder Flussmündungen unter stärkerem Gezeiteneinfluss werden aber gemieden.[6] Das Vorkommen des Pazifischen Maulwurfs ist positiv gekoppelt mit der Dichte der Regenwürmer in einer Region. In zuträglichen Lebensräumen können bis zu 10 Individuen je Hektar auftreten, die Populationsdichte geht in Bereichen mit einer geringen Beutedichte auf bis zu einem Individuum auf 14 ha zurück.[5] Die Höhenverbreitung in British Columbia reicht nur selten über 350 m hinaus. In weiten Teilen seines Verbreitungsgebietes tritt der Pazifische Maulwurf sympatrisch mit dem Townsend-Maulwurf (Scapanus townsendii) auf. In diesen Fällen bevorzugt letztere Art häufiger offenere Landschaften mit schweren Böden, erstere hingegen Flusstäler und Wälder. Allerdings nutzen beide Vertreter teils syntopisch die gleichen Wiesen und Ackerflächen, während der Townsend-Maulwurf nicht in Wälder vordringt, wo der Pazifische Maulwurf anwesend ist.[6][2][3][4]
Lebensweise
Territorialverhalten
Die Lebensweise des Pazifischen Maulwurfs ist nur punktuell gut untersucht. Er ist zumeist unterirdisch aktiv und gräbt ausgedehnte Tunnel- und Gangsysteme, die insgesamt bis zu 450 m lang sein können. Sie bestehen aus engen, nur 5 cm breiten Röhren, die sich gelegentlich in größere Kammern mit einem Durchmesser von 10 cm öffnen. Deren Funktion ist nicht genau bekannt. Prinzipiell bestehen drei verschiedene Gangebenen. Oberflächennahe Gänge werden in der Regel nur einmalig genutzt und dienen hauptsächlich der Nahrungssuche, Wanderungen oder werden von Männchen auf der Suche nach Weibchen in der Paarungszeit angelegt. Sie sind nur 2,5 bis 5 cm tief. Bei diesen Tunneln gräbt ein Tier nicht direkt, sondern drückt das Erdmaterial nach oben, so dass auf der Erdoberfläche kleine Rippeln entstehen. Auf diese Weise kann es bis zu 275 m in einer einzigen Nacht bewältigen. Weitere Gänge zur Nahrungssuche verlaufen in 15 bis 20 cm, teilweise auch bis in 90 cm Tiefe und werden täglich aufgesucht. Sie fungieren gleichzeitig als Fallen für bodenlebende Wirbellose. Hier befinden sich auch häufig die Nester. Zusätzlich werden die Nester durch Baumwurzeln oder ähnliches geschützt. Sie sind mit Pflanzen ausgekleidet. Von der dafür notwendigen Kammer führen radial mehrere Gänge fort. Ein dritter Gangtyp reicht 100 bis 200 cm in den Untergrund und entsteht zumeist in trockenen Jahresabschnitten.[5][6][2][3][4] In den Tunneln und Gängen kommt es wegen der fehlenden Frischluftzufuhr teilweise zu einer Verringerung des Sauerstoffgehaltes auf bis zu 14,3 % (normal 20,9 %) und gleichzeitig zu einem Anstieg des Kohlenstoffdioxid auf bis zu 5,5 % (normal 0,04 %), der durch das Atmen angereichert wird. Eine Beeinträchtigung der Aktivitäten der Tiere ist dadurch nicht zu verzeichnen.[7] Als eine Anpassung daran ist beim Pazifischen Maulwurf wie bei einigen anderen grabenden Maulwurfsarten das Hämoglobin modifiziert, wodurch eine erhöhte Sauerstoff-Bindungsaktivität ermöglicht wird. Diese wirkt auch unbeeinflusst von den relativ niedrigen Temperaturen in den Tunneln.[8][9]
Der Pazifische Maulwurf gräbt zwar ganzjährig, die intensivste Periode fällt aber in den Herbst und Winter, wenn die Böden gut durchfeuchtet und locker sind. Bei den Grabungsaktivitäten schafft das Tier den Aushub an die Erdoberfläche, wodurch die charakteristischen Auswurfhügel (Maulwurfshügel) entstehen. Ein einzelner Hügel hat einen Durchmesser von etwa 30 cm, ist 15 cm hoch und besteht aus 2,6 l Erdmaterial. In seiner aktiven Phase zum Jahresende und -wechsel kann der Pazifische Maulwurf 200 bis 400 Hügel anlegen. Die Anzahl der Hügel in einer Region korreliert mit der Dichte an Beutetieren.[5][10] Für die Errichtung eines Hügels benötigt er rund 33 Minuten.[11][6][2][3][4]
Wie andere grabende Maulwürfe auch ist der Pazifische Maulwurf territorial und einzelgängerisch. Nur während der Paarungszeit kommen männliche und weibliche Tiere zusammen. Jedes Individuum unterhält ein einzelnes Revier, das rund 0,12 ha groß ist. Die randlichen Bereiche werden nur selten aufgesucht. Ein über einen Zeitraum von 50 Stunden beobachtetes Tier bewegte sich in einem Umkreis von 39 m. Die Aktivitätszeiten dauerten bis zu fünf Stunden an, beschränken sich dabei jedoch nicht auf eine bestimmte Tageszeit. Dadurch bestand auch keine Synchronität zwischen benachbarten Individuen. Die längste Schlafphase währte rund vier Stunden.[11] Die Tiere sind außerdem gute Schwimmer und können Gebiete nach Überschwemmungen rasch neu besiedeln. Lediglich Jungtiere, die das mütterliche Gebiet verlassen, sind im Sommer stärker oberirdisch aktiv.[6][2][3][4]
Ernährung
Die Hauptnahrung des Pazifischen Maulwurfs besteht aus Regenwürmern. Als weitere Komponenten kommen Schnecken, Hundertfüßer, pflanzliches Material und verschiedenen Alt- und Jungstadien von Insekten, darunter Schmetterlinge, Zweiflügler, Käfer, Heuschrecken und Hautflügler, hinzu. Nach Untersuchungen von 108 Mageninhalten aus British Columbia enthielten rund 93 % Regenwürmer und nur jeweils 2 % Larven beziehungsweise ausgewachsene Insekten.[5] Eine vergleichbare Studie an 25 Mageninhalten aus Oregon erbrachte in 92 % Regenwürmer, während 48 % Hundertfüßer und 28 % Schnecken enthielten. Im Volumenanteil der Nahrung erreichten Regenwürmer hingegen 56 %, Hundertfüßer und Ameisen brachten es jeweils rund 6 %. Auch wurden Reste von kleinen Säugetieren gefunden. Pflanzliches Material war mit insgesamt 4 % vertreten, einschließlich Pilze.[12] Wiederum acht analysierte Mageninhalte aus Oregon bestanden zu 69,9 % aus Regenwürmern, 25,1 % aus Zweiflüglern, 1,6 % aus Schmetterlingen, 0,9 % aus Käfern, 0,6 % aus Hundertfüßern und der Rest aus Heuschrecken, Hautflüglern sowie Pflanzen.[13] In Gefangenschaft gehaltene Tiere fraßen 100 bis 150 g Regenwürmer täglich, was das bis zum Zweifachen ihres Körpergewichts entspricht, benötigten dafür aber kein Wasser. Hochrechnungen zufolge vertilgen in Gebieten mit hoher Populationsdichte vier Individuen auf rund 0,4 ha Fläche jährlich bis zu 219.000 Regenwürmer, was bis zu 150 Würmer pro Individuum und Tag sind. Der Pazifische Maulwurf ist dadurch kontinuierlich auf Nahrungssuche und unterbricht diese Aktivität eher selten für maximal eine Stunde.[5] Sie erfolgt meist unterirdisch, nur unter Umständen kommt ein Tier dafür an die Erdoberfläche.[14][2][3][4]
Fortpflanzung
Die Paarung erfolgt normalerweise zwischen Ende Januar und Anfang März. Während dieser Zeit vergrößern sich die Hoden der Männchen merklich bis auf Bohnengröße mit einem Durchmesser von gut 10 bis 15 mm. Die Männchen erweitern dann ihre Tunnelsysteme, um die Territorien der Weibchen zu erreichen. Wahrscheinlich pflanzen sie sich dadurch polygyn fort. Die Dauer der Tragzeit ist unbekannt, der einzige Wurf im Jahr kommt aber im Zeitraum von März bis April zur Welt. In der Regel umfasst er zwei bis vier Jungen. Die Größe ist wahrscheinlich abhängig vom Alter des Muttertiers, da einzelnen Beobachtungen zufolge junge Weibchen zwei, ältere hingegen vier Neugeborene zur Welt bringen. Diese sind rund 5,5 cm lang, 13 bis 14 g schwer und nackt. Sie verbleiben zunächst in einem Nest aus grobem Pflanzenmaterial, das sich 15 cm unter der Oberfläche befindet und einen Durchmesser von rund 20 cm aufweist. Nach der Entwöhnung gehen sie auf Nahrungssuche im Gangsystem der Mutter für möglicherweise mehrere Wochen. Ein im April untersuchtes Gangsystem enthielt drei Jungen von je 32 g Körpergewicht, was etwa der Hälfte eines ausgewachsenen Individuums entspricht. Die Geschlechtsreife wird mit neun bis zehn Monaten erreicht. Das individuelle Höchstalter beträgt wahrscheinlich drei bis vier Jahre. Das Geschlechterverhältnis bei bisher gefangenen Individuen lag bei etwa 1:1. Von 940 untersuchten Tieren aus British Columbia waren rund 45 % einjährig und rund 6 % über drei Jahre alt.[5][2][3][4]
Fressfeinde und Parasiten
Bedeutende Fressfeinde des Pazifischen Maulwurfs sind mit mehreren Vögeln dokumentiert. Dazu gehören unter anderem die Schleiereule und die Waldohreule. Bei letzterer liegt der Anteil der Maulwurfsart im gesamten Beutespektrum nach Untersuchung von rund 140 Gewöllen aus Oregon bei unter 1 %.[15] Nur unwesentlich mehr beträgt der Anteil bei ersterer. Basis hierfür bilden mehr als 11.700 Gewöllreste aus British Columbia. Der Großteil der Individuen wird im Frühjahr und Sommer geschlagen, was wohl mit den Wanderungsbewegungen der Jungen zusammenhängt, die das mütterliche Nest verlassen.[16][17] Ein vergleichbares Bild ergab sich bei der Studie von über 720 Gewöllen der Schleiereule in Oregon. Insgesamt fanden sich hierbei 28 Reste des Pazifischen Maulwurfs.[18] Unter den Schlangen kommt die Gummiboa als Beutegreifer hinzu. Gelegentlich töten auch Hauskatzen und Haushunde einen Pazifischen Maulwurf, fressen diesen aber nicht.[6] Als Kommensalen treten Feldmäuse und die Hausmaus in Erscheinung, die die Tunnel des Pazifischen Maulwurfs nutzen.[19][2]
Es sind überwiegend äußere Parasiten belegt, ein größerer Teil davon repräsentiert Milben. In erster Linie handelt es sich um Raubmilben sowie deren Verwandte wie Androlaelaps, Eulaelaps, Echinonyssus und Haemogamasus, untergeordnet sind auch Prostigmata wie Pygmephorus präsent. Des Weiteren kommen einzelne nicht-parasitäre Milben vor.[12][20] Flöhe werden durch Corypsylla, Nearctopsylla und Catallagria angezeigt, Zecken wiederum durch Ixodes und Hirstionyssus.[21] Als innere Parasiten wurden Fadenwürmer benannt, so unter anderem Tricholinstowia.[22][23][2]
Systematik
Innere Systematik der Westamerikanischen Maulwürfe nach Álvarez-Castañeda und Cortes-Calva 2021[24]
|
Der Pazifische Maulwurf ist eine Art innerhalb der Gattung der Westamerikanischen Maulwürfe (Scapanus). Der Gattung werden insgesamt fünf Vertreter zugewiesen. Sie gehört wiederum zur Familie der Maulwürfe (Talpidae) und innerhalb dieser zusammen mit einigen weiteren Gruppen aus Nordamerika und Asien zur Tribus der Neuweltmaulwürfe (Scalopini). Die Neuweltmaulwürfe repräsentieren vergleichbar den Eigentlichen Maulwürfen (Talpini) grabende Angehörige der Familie, andere Vertreter leben nur teils unterirdisch, bewegen sich auf der Erdoberfläche fort oder verfolgen eine semi-aquatische Lebensweise. Nach molekulargenetischen Untersuchungen spalteten sich die Neuweltmaulwürfe im Oberen Eozän vor rund 39 bis 35 Millionen Jahren von den anderen Triben der Maulwürfe ab.[25] Die Tribus besteht aus zwei Entwicklungslinien: die Parascalopina und die Scalopina, die beide vor allem an der Ausprägung des Metastylids am unteren zweiten Molar differenzierbar sind.[26][27] Den Scalopina fehlt das Metastylid, bei den Parascalopina hingegen kommt es vor. Beide Linien entwickelten sich bereits seit dem Unteren Miozän vor 21,4 Millionen Jahren getrennt voneinander. Die Westamerikanischen Maulwürfe sind den Scalopina zuzurechnen und gehören somit in das nähere Verwandtschaftsumfeld des Ostamerikanischen Maulwurfs (Scalopus). Die Scalopina diversifizierten sich im Mittleren Miozän vor gut 14 Millionen Jahren, die deutlicher Auffächerung der Westamerikanischen Maulwürfe reicht in den Übergang vom Miozän zum Pliozän vor gut 6 Millionen Jahren zurück. Als Schwesterart des Pazifischen Maulwurfs ist der Townsend-Maulwurf (Scapanus townsendii) aufzufassen.[28][29]

Die wissenschaftliche Erstbeschreibung des Pazifischen Maulwurfs geht auf Frederick William True aus dem Jahr 1896 zurück. Sie erfolgte im Rahmen einer größeren Revision der damals bekannten Maulwurfsarten in Nordamerika. True verwies auf die Ähnlichkeit zum Townsend-Maulwurf, stellte jedoch als abtrennendes Merkmal die geringere Körpergröße heraus. Das Typusexemplar bildet ein weibliches Tier, das bereits im August 1855 an der „Shoalwater Bay“ im US-amerikanischen Bundesstaat Washington von James Graham Cooper aufgesammelt worden war. Das Fundgebiet wird heute als Willapa Bay bezeichnet und befindet sich im Pacific County. Es stellt die Typusregion der Art dar. Mit dem Artepitheton orarius verwies True auf die küstennahe Verbreitung der Tiere (aus dem Lateinischen für „Küste“).[30][6][31]
True schloss damals auch die Populationen weiter im Landesinneren in seine Art mit ein. Knapp 20 Jahre später, im Jahr 1915, setzte Hartley H. T. Jackson diese während einer erneuten Überprüfung der nordamerikanischen Maulwürfe als eigene Unterart ab, was er mit den durchschnittlich größeren Körpermaßen, der helleren Fellfärbung und dem kürzeren und breiteren Rostrum begründete. Als Terra typica wies er Walla Walla im gleichnamigen County in Washington aus. Das Belegexemplar hierfür, ein männliches Individuum, war wiederum im Jahr zuvor von Theodore H. Scheffer entdeckt worden, nach dem Jackson seine neue Form benannte.[1] Dadurch sind heute zwei Unterarten des Pazifischen Maulwurfs anerkannt:[2][32][4][31]
- S. o. orarius True, 1896; Nominatform, Pazifikküste Nordamerikas vom westlichen Washington über das westliche und zentrale Oregon bis in das nordwestliche Kalifornien
- S. o. schefferi Jackson, 1915; vom südwestlichen British Columbia über das zentrale und östliche Washington bis in das nordöstliche Oregon und westzentrale Idaho
Der Status der östlichsten Populationen jenseits des Kaskadengebirges wird teilweise als problematisch erachtet, da die Tiere dort weitgehend nur aus Museumsbeständen dokumentiert sind.[3] Im Jahr 1944 etablierten Walter W. Dalquest und Victor B. Scheffer mit S. o. yakimensis eine weitere Unterart anhand eines nicht ganz ausgewachsenen Jungtiers aus dem Yakima County in Washington. Dieses stand bezüglich Körpergröße und Fellfärbung intermediär zu den beiden anderen Unterarten.[33] Sie wird heute als synonym zu S. o. schefferi aufgefasst.[2][31]
Es sind keine Fossilfunde des Pazifischen Maulwurfs überliefert. Mit Scapanus hagermanensis liegt aus den älteren Abschnitten der Glenns-Ferry-Formation im US-Bundesstaat Idaho eine ausgestorbene Form vor, die der Verwandtschaftsgruppe aus dem Townsend-Maulwurf und dem Pazifischen Maulwurf näher als dem Kalifornischen Maulwurf (Scapanus latimanus). Sie gehört der Hagerman fauna an und datiert in das Pliozän. Die Gattung Scapanus selbst ist erstmals im Verlauf des Miozäns nachgewiesen.[34]
Bedrohung und Schutz
Der Pazifische Maulwurf wird von der IUCN als „nicht gefährdet“ (least concern) eingestuft. Er ist weit verbreitet ist, kommt relativ häufig vor und hat sich an eine Vielzahl verschiedener Lebensräume angepasst. Bestandsgefährdende Bedrohungen sind nicht bekannt.[35] Lokal bestehen nur wenige Konflikte mit dem Menschen, da die Art häufiger in Wäldern auftritt und seltener auf Wiesen und in Gärten. In letzteren beiden Arealen können die Tiere durch ihre Grabungsaktivitäten Pflanzen schädigen. Zur Bestandskontrolle erwiesen sich lediglich Tierfallen (überwiegend Klapp- und Springfallen) als effektiv.[5] Andere Kontrollmechanismen wie die Anreicherung der Böden mit Stickstoff zur Ausdünnung der Beutetiere zeigten bisher keine ausreichende Wirkung.[36]
Literatur
- Gregory D. Hartman und Terry L. Yates: Scapanus orarius. Mammalian Species 253, 1985, S. 1–5
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 599) ISBN 978-84-16728-08-4
- B. J. Verts und Leslie N. Carraway: Land mammals of Oregon. University of California Press, Berkeley, 1998, S. 69–71
Einzelnachweise
- Hartley H. T. Jackson: A review of the American moles. North American Fauna 38, 1915, S. 1–100 (S. 61–64)
- Gregory D. Hartman und Terry L. Yates: Scapanus orarius. Mammalian Species 253, 1985, S. 1–5
- B. J. Verts und Leslie N. Carraway: Land mammals of Oregon. University of California Press, Berkeley, 1998, S. 69–71
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 599) ISBN 978-84-16728-08-4
- R. Glendenning: Biology and control of the Coast mole, Scapanus orarius True, in British Columbia. Canadian Journal of Animal Science 39, 1959, S. 34–44
- Chris Maser, Bruce R. Mate, Jerry F. Franklin und C. T. Dyrness: Natural history of Oregon Coast mammals. United States Department of Agriculture, Forest Service, General Technical Report PNW-133, 1981, S. 1–496 (S. 77–79)
- Valentin H. Schaefer und R. M. F. S. Sadleir: Concentrations of carbon dioxide and oxygen in mole tunnels. Acta Theriologica 24 (21), 1979, S. 267–271
- Kevin L. Campbell, Jay F. Storz, Anthony V. Signore, Hideaki Moriyama, Kenneth C. Catania, Alexander P. Payson, Joseph Bonaventura, Jörg Stetefeld und Roy E. Weber: Molecular basis of a novel adaptation to hypoxic-hypercapnia in a strictly fossorial mole. BMC Evolutionary Biology 10, 2010, S. 214, doi:10.1186/1471-2148-10-214
- Anthony V. Signore, Jörg Stetefeld, Roy E. Weber und Kevin L. Campbell: Origin and mechanism of thermal insensitivity in mole hemoglobins: a test of the ‘additional’ chloride binding site hypothesis. Journal of Experimental Biology 215 (3), 2012, S. 518–525, doi:10.1242/jeb.063669
- Valentin H. Schaefer und R. M. F. S. Sadleir: Factors influencing molehill construction by the coast mole (Scapanus orarius True). Mammalia 45 (1), 1981, S. 31–38
- Valentin H. Schaefer: Movements and diel activity of the coast mole Scapanus orarius True. Canadian Journal of Zoology 60, 1982, S. 480–182
- John O. Whitaker, Jr., Chris Maser und Richard J. Pedersen: Food and Ectoparasitic Mites of Oregon Moles. Northwest Science 53 (4), 1979, S. 268–273
- A. W. Moore: Food habits of Townsend and Coast moles. Journal of Mammalogy 14, 1933, S. 36–40
- Vladimir Dinets: Surface foraging in Scapanus moles. Mammalia 82 (1), 2018, S. 48–53, doi:10.1515/mammalia-2016-0091
- Chris Maser und Edmund D. Brodie, Jr.: A Study of Owl Pellet Contents from Linn, Benton and Polk Counties, Oregon. he Murrelet 47 (1), 1966, S. 9–14
- I. McT. Cowan: Food habits of the barn owl in British Columbia. The Murrelet 23, 1942, S. 48–53
- R. Wayne Campbell, David A. Manuwal und Alton S. Harestad: Food habits of the common barn owl in British Columbia. Canadian Journal of Zoology 65 (3), 1987, S. 578–586
- Richard D. Giger: Surface Activity of Moles as Indicated by Remains in Barn Owl Pellets. The Murrelet 46 (3), 1965, S. 32–36
- Leo K. Couch: Mice and Moles. Journal of Mammalogy 5 (4), 1924, S. 264
- John O. Whitaker, Jr. und Chris Maser: Mites (excluding chiggers) of mammals of Oregon. Great Basin Naturalist 45 (1), 1985, S. 67–76
- Terry L. Yates, Danny B. Penc und Glenn K. Launchbaugh: Ectoparasites from seven species of North American moles (Insectivora: Talpidae). Journal of Medical Entomology 16, 1979, S. 166–168
- Marie-Claude Durette-Desset und Claude Vaucher: Nématodes Héligmosomes parasites d’insectivores Talpidés de la région holarctique. Annales de Parasitologie 49 (2), 1974, S. 191–200
- John M. Kinsella und Vasyl V. Tkach: Checklist of helminth parasites of Soricomorpha (= Insectivora) of North America north of Mexico. Zootaxa 1969, 2009, S. 36–58
- Sergio Ticul Álvarez-Castañeda und Patricia Cortes-Calva: Revision of moles in the genus Scapanus. Therya 12 (2), 2021, S. 275–281, doi:10.12933/therya-21-1174
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang, Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- J. Howard Hutchinson: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117 (S. 58–96)
- Achim Schwermann, Kai He, Benjamin J. Peters, Thorsten Plogschties und Gabrielle Sansalone: Systematics and macroevolution of extant and fossil scalopine moles (Mammalia, Talpidae). Palaeontology 62 (4), 2019, S. 661–676, doi:10.1111/pala.12422
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Zhong-Zheng Chen, Shui-Wang He, Wen-Hao Hu, Wen-Yu Song, Kenneth O. Onditi, Xue-You Li und Xue-Long Jiang: Morphology and phylogeny of scalopine moles (Eulipotyphla: Talpidae: Scalopini) from the eastern Himalayas, with descriptions of a new genus and species. Zoological Journal of the Linnean Society, 2021, S. zlaa172, doi:10.1093/zoolinnean/zlaa172
- Frederick W. True: A revision of the American moles. Proceedings of the United States National Museum 19 (1101), 1896, S. 1–112 (S. 52)
- Neal Woodman: American Recent Eulipotyphla. Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithsonian Institution Scholary Press, 2018, S. 1–107 (S. 27)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Walter W. Dalquest und Victor B. Scheffer: A New Mole from Washington State. The Murrelet 25 (2), 1944, S. 27–28
- J. Howard Hutchinson: Late Pliocene (Blancan) Scapanus (Scapanus) (Talpida: Mammalia) from the Glenns Ferry Formation of Idaho. PaleoBios 12 (45), 1987, S. 1–7
- F. Cassola: Scapanus orarius (errata version published in 2017). The IUCN Red List of Threatened Species 2016. e.T41474A115188698 (); zuletzt aufgerufen am 10. April 2021
- Valentin H. Schaefer: A Test of the Possible Reduction of the Digging Activity of Moles in Pastures by Increasing Soil Nitrogen. Acta Theriologica 26 (7), 1981, S. 118–123
Weblinks
- Scapanus orarius in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: F. Cassola, 2016. Abgerufen am 10. April 2021.