Komplexe Plastiden
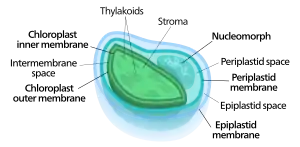
Komplexe Plastiden sind gekennzeichnet durch Membranhüllen, die anders als bei einfachen Plastiden aus drei oder mehr Membranen bestehen. Enthalten die Plastiden Chlorophyll, dann spricht man auch von komplexen Chloroplasten. Im Raum zwischen der zweiten und dritten Membran (von innen) befindet sich in manchen Fällen ein rudimentärer eukaryotischer Zellkern, genannt Nucleomorph. Dies ist beispielsweise bei Cryptomonaden, Chlorarachniophyten und vielen Apicomplexa der Fall.[1][2][3]
Die Endosymbiontentheorie erklärt die einfachen Chloroplasten (mit ihrer Doppelmembran) als Ergebnis einer primären Endosymbiose, bei der eine heterotrophe eukaryotische Zelle (Eucyte) ein photosynthetisch aktives (autotrophes) Cyanobakterium mit seiner doppellagigen gramnegativen Zellwand aufgenommen hat, ohne dieses zu ‚verdauen‘ (zu lysieren). Die komplexen Chloroplasten (ggf. mit ihrem Nucleomorph) werden als das Ergebnis einer sekundären oder Endosymbiose erklärt, bei der eine heterotrophe eukaryotische Zelle eine (aus einer primären Endosymbiose hervorgegangene) autotrophe (ebenfalls eukaryotische) Algenzelle mitsamt Chloroplasten aufgenommen und als Endosymbiont behalten hat. Dieser Endosymbiont hatte zunächst neben dem oder den Chloroplasten einen eigenen Zellkern, eigene eukaryotische Ribosomen und Mitochondrien. Im Laufe einer langen Partnerschaft kam es jedoch zu einer Reihe von Vereinfachungen und dem Verlust redundanter Teile:
- Nach verschiedenen Anpassungen und endosymbiontischem Gentransfer (EGF) von Genen im Kern der phagozytierten Alge auf die äußere Wirtszelle[1] wurde ihr Zellkern zum Nucleomorph reduziert oder verschwand ganz, mit ihm die eigenen Ribosomen,
- Ähnlich erging es den eigenen Mitochondrien.
- Auch einzelne Lagen der vielschichtigen Membranhülle konnten verloren gehen.[1][4][5]
Die so entstandenen sekundären, komplexen Chloroplasten können daher drei oder vier Membranen aufweisen,[6] die beiden Cyanobakterienmembranen, manchmal die Zellmembran der inkorporierten Algen und die Phagosomenvakuole aus der Zellmembran des Wirts.[1] Selbst die Fähigkeit zur Photosynthese konnte verloren gehen, wenn der Endosymbiont aufgrund diverser anderer Leistungen für die Wirtszelle weiter von Nutzen war, so dass der Endosymbionten dann allgemeiner einen komplexen Plastiden darstellt. Da komplexe Plastiden von eukaryotischen Einzellern (Eucyten) abstammen, können diese im sog. Periplastidalraum (außerhalb des inneren, primären Plastiden) neben dem Nukleomorph noch weitere eukaryotische Strukturen (heißt eigene Organellen) aufweisen, wie man sie bei einfachen Plastiden (mit ihrem cyanobakteriellen Ursprung) nicht findet, z. B. ein Endoplasmatisches Retikulum (siehe Prymnesium parvum).[7]
Sekundäre und tertiäre Endosymbiose
Wurde eine solche Alge mit sekundärem Plastiden nach Art der Matrjoschka-Puppen nochmals von einer heterotrophen Eucyte aufgenommen, entstand eine tertiäre Endosymbiose. Je nachdem, wie viele solche verschachtelten Endosymbiosen zu einem komplexen Chloroplasten (oder Plastiden) geführt haben, wird dieser als sekundärer oder tertiärer Chloroplast (Plastid) bezeichnet, der Wirt entsprechend etwa zum Sekundärwirt.[8] Chloroplasten und Plastiden finden sich heute in einer extrem breiten Reihe von Organismen, von denen etliche nicht einmal direkt miteinander verwandt sind. Dies wird als eine Folge vieler sekundärer und sogar tertiärer endosymbiotischer Ereignisse erklärt. Diese Ereignisse schufen eine Vielzahl von eukaryotisch-eukaryotischen Chimären, die als Meta-Algen (en. meta-algae) bezeichnet werden.[9]
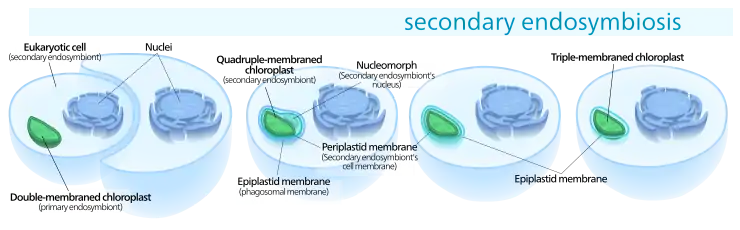
Eine eukaryotische Alge wird einverleibt von einem anderen Eukaryoten und wird damit zu einem Chloroplasten mit drei oder vier Membranen.
Alle sekundären Chloroplasten stammen von Grün- und Rotalgen ab; es wurden (bisher) keine von Glaucophyten abstammenden sekundären Chloroplasten gefunden – wahrscheinlich weil Glaucophyten in der Natur relativ selten sind und es daher unwahrscheinlich ist, dass sie von einem anderen Eukaryoten aufgenommen wurden.[1]
Eine alternative Darstellung dieser komplexen Evolution findet sich bei Sven. B. Gould et al. (2008).[9]
Von Grünalgen abstammende Chloroplasten
Grünalgen wurden von den Eugleniden, Chlorarachniophyten, einer Linie von Dinoflagellaten[10] und dem möglichen gemeinsamen Vorfahren der „CASH-Linie“ (Cryptophyten, Alveolaten, Stramenopilen und Haptophyten)[11] in drei oder vier verschiedenen Endosymbioseereignissen aufgenommen.[12] Viele der der von Grünalgen abstammenden Chloroplasten enthalten Pyrenoide, aber anders als Chloroplasten in ihren Grünalgen-Vorfahren sammelt sich die Stärke als Speicherprodukt außerhalb des Chloroplasten als Granulat.[2]
Euglenophyten
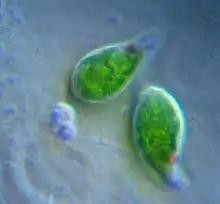
Euglenophyten (Euglenida) sind eine Gruppe von Flagellaten (taxonomisch genauer: Excavata bzw. Discoba), die von Grünalgen abstammende sekundäre Chloroplasten enthalten.[1] Die Chloroplasten der Euglenophyten haben drei Membranen. Man nimmt an, dass die Membran der aufgenommenen Grünalge (das ist die zweite von außen) verlorengegangen ist und die beiden Membranen der Cyanobakterien innen und die Phagosomenmembran des sekundären Wirts außen übriggeblieben sind.[1] Euglenophyten-Chloroplasten haben ein Pyrenoid und Thylakoide in Dreiergruppen gestapelt. Das Photosyntheseprodukt wird in Form von Paramylon gespeichert, das in membrangebundenen Granula im Zytoplasma des Euglenophyten enthalten ist.[2][10]
Chlorarachniophyten
Chlorarachniophyten sind eine kleine Gruppe von Organismen, die ebenfalls von Grünalgen abstammende sekundäre Chloroplasten,[1] obwohl ihre Geschichte komplizierter ist als die der Euglenophyten. Es wird angenommen, dass der Vorfahre der Chlorarachniophyten ein Eukaryot mit einem ursprünglichen aus Rotalgen abstammenden Chloroplasten war. Dieser ging später verloren, worauf es erneut zu einer sekundären Endosymbiose durch Aufnahme einer Grünalge kam, aus der sich der heutige komplexe Chloroplast entwickelte.[10]
Die (heutigen) Chloroplasten der Chlorarachniophyten sind von vier Membranen umgeben, außer in der Nähe der Zellmembran, wo sie zu einer Doppelmembran verschmelzen; ihre Thylakoide sind in losen Stapeln angeordnet.[2] Chlorarachniophyten erzeugen ein Polysaccharid namens Chrysolaminarin, das sie im Zytoplasma speichern[10], sich aber häufig um das Chloroplasten-Pyrenoid ansammelt, das in das Zytoplasma hinausragt.[2]
Die Chloroplasten der Chlorarachniophyten zeichnen sich dadurch aus, dass die Grünalge, von der sie abstammen, noch nicht vollständig abgebaut wurde – ihr Kern bleibt als Nucleomorph zwischen der zweiten und der dritten Chloroplastenmembran bestehen. Dieser entspricht dem Zytoplasma der Grünalge und wird Periplastidraum (englisch periplastid space) genannt.[1][2][10]
Von Prasinophyten abstammende Chloroplasten der Dinoflagellaten
Unter den Dinoflagellaten (alias Dinophyten) gibt es eine Gruppe naher Verwandter ihres Mitglieds Lepidodinium viride (früher zur Gattung Gymnodinium gestellt[13]), die ebenfalls ihren ursprünglichen (das rote Pigment Peridinin enthaltenden, näheres s. u.) Chloroplasten verloren und durch einen von Grünalgen (genauer gesagt Prasinophyten) abstammenden Chloroplasten ersetzt haben.[14][2]
Durch diese Ersetzung sind Lepidodinium und seine nahen Verwandten die einzigen Dinoflagellaten, die einen Chloroplasten haben, der nicht aus der Rhodoplastenlinie stammt. Dieser Chloroplast ist von zwei Membranen umgeben und hat kein Nucleomorph – alle Nucleomorph-Gene wurden auf den Dinophytenkern übertragen.[14] Es ist wahrscheinlicher, dass das endosymbiotische Ereignis, das zum heutigen Chloroplasten führte, eher eine solche serielle sekundäre Endosymbiose war als eine tertiäre Endosymbiose (s. u.), d. h. der Endosymbiont war eine Grünalge, die einen primären Chloroplasten enthielt.[10]
Von Rotalgen abstammende Chloroplasten
Cryptophyten
Cryptophyceen (auch Cryptomonaden genannt) sind eine Verwandtschaftsgruppe von Algen, die einen von Rotalgen abstammenden Chloroplasten enthalten. Die Chloroplasten der Cryptophyceen enthalten ein Nucleomorph, das oberflächlich dem der Chlorarachniophyten ähnelt.[1] Diese Chloroplasten haben vier Membranen, von denen die äußerste mit dem rauen endoplasmatischen Retikulum zusammenhängt. Sie synthetisieren gewöhnliche Stärke, die in Granulatkörnern im Periplastidraum (dem Zytoplasma der Rotalge) außerhalb der ursprünglichen Doppelmembran des primären (von Cyanobakterien abstammenden) Chloroplasten liegt. In den Chloroplasten der Cryptophyceen befinden sich ein Pyrenoid und Thylakoide in Stapeln von je zwei.[2]
Diese Chloroplasten haben keine Phycobilisome,[2] aber sie haben Phycobilin-Pigmente, die sie in ihrem Thylakoidraum aufbewahren, anstatt sie an der Außenseite ihrer Thylakoidmembran zu verankern.[2][1]
Cryptophyceen könnten eine Schlüsselrolle bei der Verbreitung von Chloroplasten bilden, die von Rotalgen abstammen.[15][16]
Die nächsten Verwandten der Cryptophyceae sind die meist farblosen und phagotroph lebenden Katablepharidophyta (syn. Katablepharida).[17] Zusammen bilden sie das Taxon der Cryptophyta. Im Gegensatz zu den Cryptophyceen haben die Katablephariden ihren Rotalgen-Chloroplasten aber wieder verloren.
Haptophyten
Haptophyten (alias Prymnesiophyta) sind ähnlich und wohl eng verwandt mit den Cryptophyceen und Heterokonta.[10] Ihren Chloroplasten fehlt ein Nucleomorph,[2][1] ihre Thylakoide bilden Dreierstapel und sie synthetisieren das Polysaccharid Chrysolaminarin, das sie vollständig außerhalb des Chloroplasten im Zytoplasma (des Haptophyten) lagern.[2]
Heterokonta (Stramenopile)
Die Heterokonta (auch Heterokontophyta, Stramenopile, oder Chromista genannt) sind eine sehr große und vielfältige Gruppe von Eukaryoten. Zu ihnen gehört die photoautotrophe Abstammungslinie der Ochrophyta mit folgenden Untergruppen:
- Kieselalgen (syn. Diatomeen oder Bacillariophyta)
- Goldalgen (syn. Chrysophyceae oder Chrysophyta s. s.)
- Gelbgrünalgen (syn. Xanthophyceae)
- Braunalgen (syn. Phaeophyceae oder Phaeophyta)
Die ersten drei Gruppen werden gelegentlich auch als Chrysophyta im weiteren Sinne (s. l.) bezeichnet.
Gemeinsam sind diesen die von Rotalgen stammenden Chloroplasten.[10][18] Die Chloroplasten der Heterokonta sind denen der Haptophyten sehr ähnlich und enthalten ein Pyrenoid, Triplett-Thylakoide. Sie haben mit einigen Ausnahmen[2] eine vierschichtige Hülle, wobei die äußerste Epiplastidmembran mit dem Endoplasmatischen Retikulum verbunden ist. Wie die Haptophyten speichern die Heterokonta Polysaccharid (Zucker) als Chrysolaminaringranulat im Zytoplasma.[2] Die Chloroplasten enthalten Chlorophyll a und mit wenigen Ausnahmen[2] Chlorophyll c,[1] aber auch Carotinoide, wodurch ihnen ihre vielfältigen Farben verliehen werden.[18]
Alveotalta (Apicomplexa, Chromerida und Dinoflagellata)
Die Alveolaten sind eine große Gruppe einzelliger Eukaryoten mit autotrophen, heterotrophen und mixotrophen Mitgliedern. Das bemerkenswerteste gemeinsame Merkmal ist das Vorhandensein von kortikalen, d. h. unter der Zellmembran (Pellicula) befindlichen, flachen Vakuolen oder Vesikeln (sackartige Strukturen), sogenannte Alveolen. Diese sind gepackt in einer durchgehender Schicht direkt unter der Membran und stützen diese, wobei sie typischerweise ein flexibles Pellikel (dünne Haut) bilden. In Dinoflagellaten bilden sie jedoch häufig panzerplattenartige Strukturen. Viele Mitglieder enthalten einen von Rotalgen abstammenden Plastiden. Ein bemerkenswertes Merkmal dieser vielfältigen Gruppe ist der häufige Verlust der Photosynthese, wobei bei einem Großteil dieser Heterotrophen weiterhin ein nicht photosynthetisches Plastid verbleibt.[19]
Apicomplexa
Die Apicomplexa sind eine Untergruppe der Alveolata. Wie die Vertreter der Gattung Helicosporidium (siehe Protothekose) sind sie parasitär und haben einen nicht-photosynthetischen Chloroplasten.[10] Man nahm daher früher an, dass sie mit Helicosporidium verwandt wären, aber inzwischen ist bekannt, dass Helicosporidium zu den Grünalgen gehört und nicht Teil der CASH-Linie ist.[10] Zu den Apicomplexa gehört der Malariaparasit Plasmodium. Viele Apicomplexa haben von ihren Vorfahren ein von Rotalgen abstammendes komplexes Plastid geerbt,[20][10] Apicoplast genannt. Andere Apicomplexa wie Cryptosporidium haben dieses vollständig verloren.[20] Die Apicomplexa speichern ihre Energie in Amylopektin-Granula, die sich in ihrem Zytoplasma befinden, auch wenn sie nicht photosynthetisch aktiv sind.[2]
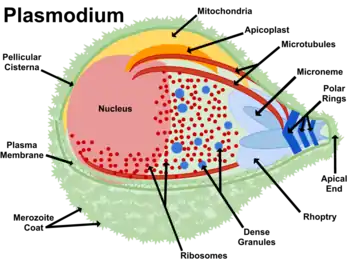
Die Apicoplasten haben alle photosynthetischen Funktionen verloren und enthalten keine photosynthetischen Pigmente oder echte Thylakoide. Sie sind an vier Membranen gebunden, aber die Membranen sind nicht mit dem Endoplasmatischen Retikulum (ER) verbunden.[2] Die Tatsache, dass Apicomplexa ihren nicht photosynthetischen Chloroplasten immer noch behalten haben, bezeugt, dass die Chloroplasten neben der Photosynthese wie bereits oben erwähnt weitere wichtige Funktionen ausüben – pflanzliche Chloroplasten versorgen Pflanzenzellen neben Zucker mit vielen wichtigen Stoffen und Apicoplasten unterscheiden sich darin nicht – sie synthetisieren Fettsäuren, Isopentenylpyrophosphat und Eisen-Schwefel-Cluster. Außerdem und führen sie einen Teil der Häm-Synthese (englisch heme pathway) aus.[20] Diese machen Apicoplasten zu einem interessanten Angriffsziel für Medikamente zur Heilung von Krankheiten, die von Apicomplexa (wie dem Malariaerreger) hervorgerufen werden.[8] Die wichtigste Funktion der Apicoplasten ist die Synthese von Isopentenylpyrophosphat. In der Tat sterben Apicomplexa, wenn etwas diese Funktion verhindert.[20]
Chromerida
Die Chromerida bilden eine relativ neu entdeckte Algengruppe, die man bei australischen Korallen gefunden hat. Sie sind enge, aber photosynthetisch aktive Verwandte der Apicomplexa. Das erste Mitglied, Chromera velia, wurde 2001 entdeckt und erstmals isoliert. Die Entdeckung von Chromera velia mit einer ähnlichen Struktur wie die Apicomplexa lieferte ein wichtiges Bindeglied in der Evolutionsgeschichte der Apicomplexa und Dinoflagellaten. Die Chloroplasten der Chromerida haben wie die Apicoplasten vier Membranen, es fehlt das Chlorophyll c. C .velia verwendet den RuBisCO-Typ II, diese Form ist homolog zu der bei Dinoflagellaten. Offenbar hat diese Abstammungslinie das RuBisCO Typ II durch horizontalen Gentransfer von einem Proteobakterium erhalten.[21]
Dinoflagellaten
Die Dinoflagellaten (syn. Dinophyten oder Panzergeißler) sind eine weitere sehr große und vielfältige Gruppe von Eukaryoten, von denen etwa die Hälfte (zumindest teilweise) photosynthetisch aktiv ist.[18][14]
Die Plastiden der Dinoflagellatenbilden keine einheitliche Abstammungslinie. Stattdessen kann man die Dinoflagellaten wie folgt klassifizieren:[1][10][14]
- Bei den meisten ihrer Chloroplasten handelt es sich um sekundäre Chloroplasten, die von Rotalgen abstammen.
- Viele andere Dinophyten haben ihre Chloroplasten verloren und gehören deshalb zur nicht photosynthetischen Gruppe der Dinoflagellaten.
- Teilweise wurden die sekundären Plastiden durch eine tertiäre Endosymbiose ersetzt[22]. D. h. nach dem Verlust des Chloroplasten haben Vertreter der obigen zweiten Gruppe eine andere eukaryotische Alge in sich aufgenommen, die einen von Rotalgen abstammenden Chloroplasten enthielt.
- Andere Dinoflagellaten ersetzten ihren ursprünglichen Chloroplasten durch einen aus Grünalgen abgeleiteten.
Die meisten Chloroplasten der Dinoflagellaten enthalten RuBisCO vom Typ II, und ggf. neben anderen die photosynthetischen Pigmente Chlorophyll a, Chlorophyll c2, Beta-Carotin, und dazu mindestens ein Dinophyten-spezifisches Xanthophyll wie Peridinin, Dinoxanthin oder Diadinoxanthin, wodurch viele ihre goldbraune Farben erhalten.[19][14] Alle Dinophyten speichern Stärke im Zytoplasma und ihre Chloroplasten haben meisten Thylakoide, die Dreierstapeln angeordnet sind.[2]
Der meisten Dinoflagellaten-Chloroplasten sind vom Peridinin-Typ, charakterisiert durch das Carotinoid-Pigment Peridinin zusammen mit Chlorophyll a und Chlorophyll c2.[1][14]
Peridinin kommt in keiner anderen Gruppe von Chloroplasten vor.[14]
Peridinin-Chloroplasten haben drei Membranen (gelegentlich nur zwei),[2], d. h. sie haben die ursprüngliche Zellmembran des Rotalgen-Endosymbionten verloren haben.[1][10]
Die äußerste Membran ist nicht mit dem endoplasmatischen Retikulum verbunden.[2][14]
Sie enthalten ein Pyrenoid und haben Thylakoide im Dreierstapel. Die Stärke wird außerhalb des Chloroplasten gefunden.[2]
Ein wichtiges Merkmal dieser Chloroplasten ist, dass ihre DNA stark reduziert und in viele kleine Ringe fragmentiert ist, ein Phänomen, wie man es ähnlich bei Mitochondrien der Menschenlaus Pediculus humanus und anderen Echten Tierläusen (Anoplura), nicht aber bei Kieferläusen (Mallophaga) beobachtet hat.[23] Das Genom ist größtenteils in den Zellkern gewandert, und im Chloroplasten verblieben sind nur kritische Gene, die mit der Photosynthese zusammenhängen.[14]
Es wird angenommen, dass die Peridinin-Chloroplasten die „ursprünglichen“ Chloroplasten der Dinoflagellaten darstellen,[14] und in mehreren anderen Dinoflagellaten-Abstammungslinien entweder verloren gingen, reduziert wurden, ersetzt wurden oder „Gesellschaft“ bekommen haben (s. u.).[10]
Fucoxanthinhaltige (von Haptophyten abstammende) Chloroplasten der Dinoflagellaten
In den Fucoxanthin-Dinophyten-Linien (einschließlich Karlodinium und Karenia)[10] verloren die Dinoflagellaten ihren ursprünglichen, aus Rotalgen stammenden Chloroplasten und ersetzten ihn durch einen neuen Chloroplasten, der von Haptophyten abstammt. Karlodinium und Karenia nahmen wahrscheinlich unterschiedliche Heterokonta auf.[10] Da die (sekundären) Chloroplasten der Haptophyten vier Membranen besitzen, wäre eigentlich zu erwarten, dass diese tertiäre Endosymbiose zu einem aus sechs Membranen bestehenden Chloroplasten führen würde, wobei die Zellmembran des Haptophyten und die phagosomale Vakuole des Dinophyten zu den ursprünglich vier Membranen hinzukommen.[24] Der Haptophyt wurde allerdings stark reduziert und hat einige Membranen und seinen Kern verloren, so dass nur noch der Chloroplasten (mit seiner ursprünglichen Doppelmembran) und je nachdem eine oder zwei zusätzliche Membranen darum übrig blieben.[10][24]
Fucoxanthinhaltige Chloroplasten sind dadurch gekennzeichnet, dass sie das Pigment Fucoxanthin (genauer 19′-Hexanoyloxyfucoxanthin[25] und/oder 19′-Butanoyloxyfucoxanthin[26]), aber kein Peridinin aufweisen. Fucoxanthin wird auch in Chloroplasten der Haptophyten gefunden, was auf die beschriebene Abstammung hinweist.[14]
Dinoflagellaten mit von Diatomeen abstammenden Chloroplasten

Einige Dinoflagellaten wie Kryptoperidinium und Durinskia (beide Peridiniaceae, auch englisch dinotoms)[10] haben einen von Diatomeen (Heterokontophyta) abgeleiteten Chloroplasten.[1] Diese Chloroplasten sind von bis zu fünf Membranen umgeben,[1] je nachdem, ob man den gesamten Diatomeen-Endosymbionten als den Chloroplasten ansieht oder nur die darin enthaltene Rotalge als Chloroplast zählt. Der Diatomeen-Endosymbiont ist relativ wenig reduziert worden – er behält immer noch seine ursprünglichen Mitochondrien[10] und verfügt über endoplasmatisches Retikulum, eukaryotische Ribosomen, einen Zellkern und natürlich den von Rotalgen abstammenden komplexen (sekundären) Chloroplasten – praktisch eine vollständige Zelle[27] – alles im Innern des Wirts.[10] Der Diatomeen-Endosymbiont kann jedoch keine eigenen Reserven speichern – sein Speicherpolysaccharid befindet sich stattdessen außen in Granulaten im Zytoplasma des Dinophyten.[2][27] Der Kern des Diatomeen-Endosymbionten ist – wie bereits erwähnt – vorhanden, kann aber wahrscheinlich noch nicht einmal als Nucleomorph bezeichnet werden, da er keine Anzeichen einer Genomreduktion aufweist und möglicherweise sogar erweitert wurde.[10] Diatomeen wurden mindestens dreimal von Dinoflagellaten als Endosymbionten vereinnahmt.[10]
Der Diatomeen-Endosymbiont ist durch eine einzige Membran begrenzt.[14] In ihrem Inneren befinden sich Chloroplasten mit vier Membranen. Wie der Diatomeen-Endosymbiont-Vorfahr haben die Chloroplasten Dreifach-Thylakoide und Pyrenoide.[27]
In einigen der Gattungen dieses Typs sind die Chloroplasten des Diatomeen-Endosymbionten nicht die einzigen Chloroplasten im Dinophyt. Ihr ursprünglicher Drei-Membran-Peridinin-Chloroplast ging nämlich gar nicht verloren, sondern wurde zu einem Augenfleck umgebildet.[1][10]
Siehe auch
- Kleptoplastiden
- Apicomplexa mit Apicoplasten
- Ocelloid
- Zooxanthelle
Weblinks
- Wilfried Probst: Frühe Evolution und Symbiose, Europa-Universität Flensburg, Institut für Biologie und Sachunterricht und ihre Didaktik: §Sekundäre Plastiden, abgerufen am 19. April 2019
Einzelnachweise
- Patrick J. Keeling: Diversity and evolutionary history of plastids and their hosts. In: American Journal of Botany. 91, Nr. 10, 2004, S. 1481–93. doi:10.3732/ajb.91.10.1481. PMID 21652304.
- Anna Stina Sandelius, Henrik Aronsson (Hrsg.); E. Kim, John M. Archibald;: Diversity and Evolution of Plastids and Their Genomes. In: The Chloroplast (= Plant Cell Monographs), Band 13 2009, ISBN 978-3-540-68692-7, S. 1–39, doi:10.1007/978-3-540-68696-5_1.
- Geoffrey I. McFadden: Chloroplast Origin and Integration. In: Plant Physiology. 125, Nr. 1, 2001, S. 50–3. doi:10.1104/pp.125.1.50. PMID 11154294. PMC 1539323 (freier Volltext).
- Jacques Joyard, Maryse A. Block, Roland Douce: Molecular aspects of plastid envelope biochemistry. In: Eur. J. Biochem.. 199, Nr. 3, 1991, S. 489–509. doi:10.1111/j.1432-1033.1991.tb16148.x. PMID 1868841.
- Chloroplast. In: Encyclopedia of Science. Abgerufen am 20. März 2019.
- Balbir K. Chaal, Beverley R. Green: Protein import pathways in 'complex' chloroplasts derived from secondary endosymbiosis involving a red algal ancestor. In: Plant Molecular Biology. 57, Nr. 3, Februar 2005, S. 333–342. doi:10.1007/s11103-004-7848-y. PMID 15830125.
- Orietta Beltrami, Marcela Escobar, Gloria Collantes: New record of Prymnesium parvum f. patelliferum (Green, Hibberd & Piennar) Larsen stat. nov. (Prymnesiophyceae) from Valparaíso Bay, in: Invest. Mar., Valparaíso, Band 35, Nr. 1, S. 97–104, Mai 2007, ISSN 0717-7178, doi:10.4067/S0717-71782007000100009, PDF
- Robert R. Wise (Hrsg.); J. Kenneth Hoober: The structure and function of plastids. Springer, Dordrecht 2006, ISBN 978-1-4020-4061-0, S. 3–21.
- Sven B. Gould, Ross F. Waller, Geoffrey I. McFadden: Plastid Evolution, in: Annu. Rev. Plant Biol., Band 59, 2008, S. 491–517, doi:10.1146/annurev.arplant.59.032607.092915. Siehe insbes. Fig. 1
- P. J. Keeling: The endosymbiotic origin, diversification and fate of plastids. In: Philosophical Transactions of the Royal Society B: Biological Sciences. 365, Nr. 1541, 2010, S. 729–48. doi:10.1098/rstb.2009.0103. PMID 20124341. PMC 2817223 (freier Volltext).
- Ahmed Moustafa, Bánk Beszteri, Uwe G. Maier, Chris Bowler, Klaus Valentin, Debashish Bhattacharya: Genomic Footprints of a Cryptic Plastid Endosymbiosis in Diatoms. In: Science. 324, Nr. 5935, 2009, S. 1724–1726. bibcode:2009Sci...324.1724M. doi:10.1126/science.1172983. PMID 19556510.
- Matthew B. Rogers, Paul R. Gilson, Vanessa Su, Geoffrey I. McFadden, Patrick J. Keeling: The Complete Chloroplast Genome of the Chlorarachniophyte Bigelowiella natans: Evidence for Independent Origins of Chlorarachniophyte and Euglenid Secondary Endosymbionts. In: Molecular Biology and Evolution. 24, Nr. 1, 2006, S. 54–62. doi:10.1093/molbev/msl129. PMID 16990439.
- Gert Hansen, Lizeth Botes, Miguel De Salas: Ultrastructure and large subunit rDNA sequences of Lepidodinium viride reveal a close relationship to Lepidodinium chlorophorum comb. nov. (= Gymnodinium chlorophorum). In: Phycological Research. 55, 2007, S. 25–41 doi:10.1111/j.1440-1835.2006.00442.x (pdf)
- Jeremiah D. Hackett, Donald M. Anderson, Deana L. Erdner, Debashish Bhattacharya: Dinoflagellates: A remarkable evolutionary experiment. In: American Journal of Botany. 91, Nr. 10, 2004, S. 1523–34. doi:10.3732/ajb.91.10.1523. PMID 21652307.
- Rafael Isaac Ponce Toledo: Origins and early evolution of photosynthetic eukaryotes (Thesis). Université Paris-Saclay, 5. März 2018, doi:10.1111/brv.12340
- Andrzej Bodył: Did some red alga-derived plastids evolve via kleptoplastidy? A hypothesis. In: Biological Reviews. 93, Nr. 1, 23. Mai 2017, ISSN 1464-7931, S. 201–222. doi:10.1111/brv.12340.
- Okamoto N., Inouye I. (2005): The Katablepharids are a distant sister group of the Cryptophyta: A proposal for Katablepharidophyta divisio nova/Kathablepharida phylum novum based on SSU rDNA and beta-tubulin phylogeny. Protist 156: S. 163–179
- Biology, 8th Edition, Campbell & Reece. Benjamin Cummings (Pearson), 2009, ISBN 978-0-321-54325-7, S. 582–592.
- Jan Janouškovec, Gregory S. Gavelis, Fabien Burki, Donna Dinh, Tsvetan R. Bachvaroff, Sebastian G. Gornik, Kelley J. Bright, Behzad Imanian, Suzanne L. Strom, et al.: Major transitions in dinoflagellate evolution unveiled by phylotranscriptomics. In: Proceedings of the National Academy of Sciences. 114, Nr. 2, 10. Januar 2017, ISSN 0027-8424, S. E171–E180. doi:10.1073/pnas.1614842114. PMID 28028238. PMC 5240707 (freier Volltext).
- Sethu C. Nair, Boris Striepen: What Do Human Parasites Do with a Chloroplast Anyway?. In: PLoS Biology. 9, Nr. 8, 2011, S. e1001137. doi:10.1371/journal.pbio.1001137. PMID 21912515. PMC 3166169 (freier Volltext).
- Antonietta Quigg, Eva Kotabová, Jana Jarešová, Radek Kaňa, Jiří Šetlík, Barbora Šedivá, Ondřej Komárek, Ondřej Prášil: Photosynthesis in Chromera velia Represents a Simple System with High Efficiency. In: PLOS ONE. 7, Nr. 10, 10. Oktober 2012, ISSN 1932-6203, S. e47036. bibcode:2012PLoSO...747036Q. doi:10.1371/journal.pone.0047036.
- Richard G. Dorrell, Alison G. Smith: Do Red and Green Make Brown?: Perspectives on Plastid Acquisitions within Chromalveolates. In: Eukaryotic Cell. 10, Nr. 7, 2011, S. 856–868. doi:10.1128/EC.00326-10. PMID 21622904. PMC 3147421 (freier Volltext).
- Renfu Shao, Ewen F. Kirkness, Stephen C. Barker: The single mitochondrial chromosome typical of animals has evolved into 18 minichromosomes in the human body louse, Pediculus humanus. In: Genome Research. 19, Nr. 5, Mai 2009, S. 904–912. doi:10.1101/gr.083188.108. PMID 19336451. PMC 2675979 (freier Volltext).
- Torstein Tengs, Ole J. Dahlberg, Kamran Shalchian-Tabrizi, Dag Klaveness, Knut Rudi, Charles F. Delwiche, Kjetill S. Jakobsen: Phylogenetic analyses indicate that the 19'Hexanoyloxy-fucoxanthin-containing dinoflagellates have tertiary plastids of haptophyte origin. In: Molecular Biology and Evolution. 17, Nr. 5, 2000, S. 718–29. doi:10.1093/oxfordjournals.molbev.a026350. PMID 10779532.
- Externe Identifikatoren von bzw. Datenbank-Links zu 19′-Hexanoyloxyfucoxanthin: CAS-Nummer: 60147-85-5, PubChem: 6443044, ChemSpider: 4947086, Wikidata: Q85748832.
- Externe Identifikatoren von bzw. Datenbank-Links zu 19′-Butanoyloxyfucoxanthin: CAS-Nummer: 111234-30-1, PubChem: 14160128, ChemSpider: 24823142, Wikidata: Q90309501.
- Eberhard Schnepf, Malte Elbrächter: Dinophyte chloroplasts and phylogeny – A review. In: Grana. 38, Nr. 2–3, 1999, S. 81–97. doi:10.1080/00173139908559217.