Myxogastria
Die Myxogastria, meist als Myxomyceten, gelegentlich auch als Echte Schleimpilze[1] oder plasmodiale Schleimpilze bezeichnet, sind eine Untergruppe der Amoebozoa. Sie umfassen rund 900 Arten in fast 60 Gattungen.
| Myxogastria | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Hängende Fruchtkörper von Badhamia utricularis | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Myxogastria | ||||||||||
| L.S.Olive |
Alle Arten durchlaufen während ihres Lebens mehrere, morphologisch sehr verschiedene Phasen von der mikroskopisch kleinen Zelle über mit bloßem Auge sichtbare, schleimige, amorphe Organismen bis hin zu teils ungewöhnlich auffällig gestalteten Fruchtkörpern. Dabei können sie beträchtliche Größen und Gewichte erreichen.
Die Gruppe ist weltweit verbreitet, in gemäßigten Breiten ist sie jedoch deutlich häufiger und weist einen höheren Artenreichtum auf als in polaren Regionen, den Subtropen oder Tropen. Hauptsächlich sind sie in lichten Wäldern anzutreffen, finden sich aber auch an extremen Standorten wie Wüsten, unter geschlossenen Schneedecken oder sogar unter Wasser.
Merkmale und Lebenszyklus
%252C_Wachstumsphase%252C_01.jpg.webp)
%252C_Wachstumsphase%252C_02.jpg.webp)
%252C_Wachstumsphase%252C_03.jpg.webp)
%252C_Konzentrationsphase%252C_01.jpg.webp)
_Reifestadium%252C_03.jpg.webp)
_Reifestadium%252C_04.jpg.webp)
_Sporenstadium%252C_01.jpg.webp)
_Sporenstadium%252C_02.jpg.webp)
_Ende_Sporenstadium%252C_01.jpg.webp)
Grob lässt sich der haplo-diplontische Lebenszyklus der Myxogastria in drei Phasen unterteilen, zuerst eine einzellige, einkernige, haploide Phase, die mit der Bildung von diploiden Zygoten beendet wird. Dann folgt ein Stadium als amorphes, vielkerniges, diploides Plasmodium sowie abschließend die reproduktive Phase mit der Ausbildung des ebenfalls diploiden Fruchtkörpers, der sogenannten Fruktifikation, bei der wieder haploide Sporen endogen gebildet werden. In jedem Stadium ändert sich das jeweilige Erscheinungsbild dabei drastisch.
Sporen
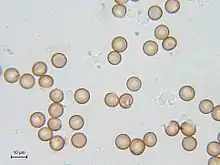
Die Sporen sind haploid, meist rund und messen zwischen 5 und 20, selten bis 24 Mikrometer im Durchmesser. Ihre Oberfläche ist üblicherweise netzartig, gratig, warzig bis stachelig strukturiert, nur sehr selten ist sie glatt. Durch die Struktur entsteht zugleich die wahrgenommene Färbung der Sporenmasse, die Sporen sind nicht pigmentiert. Bei einigen Arten (besonders bei Arten der Gattung Badhamia) bilden die Sporen Klumpen. Die „Farbe“ der Sporen, ihre Form, die Skulpturierung sowie der Durchmesser sind wichtige Merkmale zur Bestimmung.[2]
Entscheidende Faktoren für die Keimung sind vor allem Feuchtigkeit und Temperatur; welche Faktoren darüber hinaus eine Rolle spielen, ist unklar. Die Sporen bleiben meist mehrere Jahre lang keimfähig, einzelne Sporen von Herbarmaterial keimten sogar noch nach 75 Jahren. Die Sporen enthalten nach ihrer Bildung zunächst einen diploiden Zellkern, die Meiose findet in der Spore statt. Bei der Keimung öffnen sich die Sporenhüllen entweder entlang spezieller Keimporen oder Keimspalten oder reißen unregelmäßig auf und geben dann ein bis vier haploide Protoplasten frei.[2]
Myxamöben und Myxoflagellaten
Bei Arten, die sich sexuell vermehren, keimen aus den Sporen haploide Zellen. Aus der Spore keimt je nach Umgebungsbedingungen eine Myxamöbe oder ein Myxoflagellat: [3] Myxamöben bewegen sich amöboid, also auf dem Substrat kriechend fort und entstehen bei eher trockenen Bedingungen. Myxoflagellaten sind begeißelt und schwimmfähig, sie entstehen in eher feuchter bis nasser Umgebung. Myxoflagellaten weisen fast immer zwei Geißeln auf, eine Geißel ist meist kürzer und kann in einigen Fällen fast vollständig reduziert sein. Die Geißeln dienen nicht nur der Fortbewegung, sondern werden auch eingesetzt, um Nahrung heranzustrudeln. Verändert sich das Feuchtigkeitsregime, so können die Zellen zwischen den beiden Erscheinungsformen wechseln. Beide Formen besitzen keine Zellwand.[2] Dieses Entwicklungsstadium dient wie das nachfolgende der Ernährung und wird daher als erstes trophisches Stadium (Ernährungsstadium) bezeichnet. In dieser Phase ernähren sich Myxogastria von Bakterien und Pilzsporen sowie wahrscheinlich auch gelösten Stoffen. In diesem einzelligen Stadium vermehren sie sich durch einfache Zellteilung.[2]
Wenn sich in dieser Phase die Umweltbedingungen nachteilig verändern (zum Beispiel extreme Temperaturen, starke Trockenheit oder Nahrungsmangel), können Myxogastria in ein ausgesprochen langlebiges, dünnschaliges[3] Überdauerungsstadium wechseln, die sogenannte Mikrozyste. Dazu runden sich die Myxamöben ab und scheiden eine dünnwandige Zellwand aus.[4] In diesem Zustand können sie leicht ein Jahr und länger überleben. Verbessern sich die Lebensbedingungen, werden sie wieder aktiv.[5]
Zygotenbildung
Treffen in diesem Stadium zwei Zellen gleicher Art aufeinander, befruchten sie einander durch die Verschmelzung von Protoplasmen und Zellkernen zu einer diploiden Zygote. Die Bedingungen, die dies auslösen, sind nicht bekannt. Aus der diploiden Zygote entsteht durch vielfache Kernteilung ohne nachfolgende Zellteilung ein schließlich vielkerniges Plasmodium. Falls die Ausgangszellen noch begeißelt waren, wechseln sie ihre Gestalt vor der Verschmelzung von der begeißelten Form zur Myxamöbe. Für die Bildung einer Zygote sind Zellen unterschiedlichen Kreuzungstyps nötig (Heterothallie).[6][5]
Plasmodium

Mit der Bildung des Plasmodiums beginnt das zweite trophische Stadium. Der vielkernige Organismus nimmt ab nun so viel Nährstoffe per Phagozytose auf wie möglich. Dabei handelt es sich um Bakterien, Protisten, gelöste Stoffe, Schimmelpilze und höhere Pilze sowie kleine Partikel organischen Materials.[6] Sie ermöglichen der Zelle ein enormes Wachstum und ein vielfaches Teilen des Zellkerns, so dass die Zelle bald mit bloßem Auge sichtbar ist und eine Fläche – je nach Art – von bis zu einem Quadratmeter einnehmen kann (eine künstlich gezüchtete Zelle von Physarum polycephalum erreichte 1987 eine Fläche von 5,5 Quadratmeter)[7]. Myxogastria-Arten besitzen in ihrem trophischen Stadium als Plasmodium stets zahlreiche Zellkerne: bei den kleinen, nichtgeäderten Protoplasmodien sind es 8 bis 100 Kerne, bei den großen geäderten Netzwerken 100 Kerne bis 10 Millionen.[3] Alle sind jedoch nach wie vor Teil einer einzigen Zelle, die von zähflüssig-schleimiger Konsistenz und entweder transparent, weiß oder schrill gefärbt ist (orange, gelb, rosa).[5]
Die Zelle verfügt in dieser Phase über chemotaktische und (negativ) phototaktische Fähigkeiten, ist also beweglich und bewegt sich auf Substanzen wie zum Beispiel Nährstoffe zu bzw. von gefährlichen Stoffen sowie von Lichtquellen fort. Die Bewegung wird durch das körnige Plasma verursacht, das innerhalb der Zelle pulsierend in eine Richtung strömt. Dabei erreicht die Zelle eine ungewöhnlich hohe Geschwindigkeit von bis zu 1000 Mikrometer (1 mm) pro Sekunde (zum Vergleich: in Pflanzenzellen beträgt diese Geschwindigkeit 2 bis 78 Mikrometer pro Sekunde).[6] Auch in diesem Stadium kann ein Überdauerungsstadium gebildet werden, das sogenannte Sklerotium, eine ausgehärtete, widerstandsfähige Form aus zahlreichen sogenannten Makrozysten, die den Myxogastria in diesem Stadium z. B. das Überstehen von Wintern oder von Trockenheit[3] ermöglichen.[5]
Fruktifikation
Macbr.jpg.webp)


.jpg.webp)
Reife Plasmodien können dann unter angemessenen Umständen Fruchtkörper ausbilden. Die genauen Gründe, die diesen Prozess auslösen, sind nicht bekannt, bei einigen Arten werden aufgrund von Laborbeobachtungen Feuchtigkeits-, Temperatur- oder pH-Wert-Wechsel sowie Hungerphasen als auslösende Faktoren vermutet. Das Plasmodium stellt die Nahrungsaufnahme ein und kriecht – jetzt vom Licht angezogen (=„positive Phototaxis“) – an eine trockene, helle Stelle, von wo aus die spätere Verbreitung der Sporen optimal stattfinden kann. Sobald die Fruktifikation begonnen hat, ist sie nicht mehr rückgängig zu machen. Kommt es dabei zu Störungen, werden zwar sporentragende Fruchtkörper ausgebildet, diese sind allerdings häufig missgebildet.[6][5]

Die aus dem Plasmodium oder Teilen davon hervorgehenden Fruchtkörper können kleiner als einen Millimeter sein, im Extremfall aber auch Größen von bis zu einem Meter und ein Gesamtgewicht von bis zu 20 Kilogramm erreichen (Brefeldia maxima). Meist haben sie die Gestalt gestielter oder ungestielter Sporangien (wobei die Stiele nicht zellulär sind), können aber auch als geäderte bis genetzte Plasmodiokarpe, kissenförmige Aethalien oder scheinbar kissenförmige Pseudoaethalien auftreten. Fast immer weisen die Fruchtkörper am Ansatz einen Hypothallus auf. In den Fruchtkörpern finden sich die massenhaft produzierten Sporen, die bei fast allen Arten (Ausnahmen sind die Arten der Liceida sowie einige Vertreter der Gattung Echinostelium) in einer netzartigen bis fadenförmigen Struktur, dem sogenannten Capillitium, eingelagert sind.[8]
Nach Abtrocknung der geöffneten Fruchtkörper werden die Sporen hauptsächlich durch den Wind verbreitet, ebenso spielen kleine Tiere wie zum Beispiel Asseln, Milben oder Käfer eine gewisse Rolle, die entweder sporenbehaftet aus Berührungen mit Fruchtkörpern hervorgehen oder diese als Teil ihrer Nahrung aufnehmen und keimfähig wieder ausscheiden. Nur eine geringe Rolle spielt die Verbreitung durch Fließgewässer.[2]
Asexuelle Formen
Bei einigen Myxogastria gibt es asexuelle Formen, die sich apomiktisch fortpflanzen. Diese sind durchgehend diploid. Vor der Keimung der Sporen findet keine Meiose statt und die Bildung des Plasmodiums geht ohne Verschmelzung von zwei Zellen vonstatten.[5]
Verbreitung und Ökologie
Verbreitung
Myxogastria sind weltweit verbreitet, Nachweise existieren von allen Kontinenten. Grundsätzlich gilt jedoch, dass detaillierte Aussagen über die Verbreitung einzelner Taxa der Myxogastria darunter leiden, dass weite Teile der Welt nur unzureichend oder sogar noch gar nicht untersucht wurden. Als zumindest grundlegend floristisch erschlossen gelten allein Europa und Nordamerika. Die bisherigen Ergebnisse lassen darauf schließen, dass für die Mehrheit aller Arten eine kosmopolitische Verbreitung angenommen werden darf.[2]
Am häufigsten und mit dem größten Artenreichtum treten sie in gemäßigten Breiten auf, deutlich weniger häufig in polaren Regionen oder den Subtropen und Tropen.[9] Als entscheidend für die Präsenz der Arten an bestimmten Standorten gelten vor allem die physikalischen Eigenschaften des Substrates sowie klimatische Bedingungen, Endemismus ist selten.[10]
Das Verbreitungsgebiet reicht im Norden bis nach Alaska, Island, Nordskandinavien, Grönland und ins nördliche Russland. Dabei handelt es sich nicht nur um einzelne, spezielle Arten; eine Übersichtsstudie konnte allein für die arktischen und subarktischen Regionen Islands, Grönlands, Nordrusslands und Alaskas mehr als 150 verschiedene Arten nachweisen. Die Baumgrenze wird dabei deutlich überschritten, in Grönland erreicht das Verbreitungsgebiet sogar den 77. Breitengrad.[11]
In den Wäldern gemäßigter Zonen erreichen die Myxogastria dann ihren größten Artenreichtum und ihre höchste Häufigkeit, dort finden sich die idealen Voraussetzungen in Form der reichhaltigen Präsenz organischen Materials, der offenen Wälder mit hinreichender, aber nicht zu hoher Luftfeuchtigkeit und für nivicole (= schneebewohnende) Arten viele Bereiche mit einer längeren Zeit geschlossenen Schneedecke.
Nur wenige Arten besiedeln die Tropen und Subtropen. Als Gründe für die geringere Häufigkeit in tropischen Wäldern werden vor allem konstant hohe Luftfeuchtigkeiten genannt, die zum einen die notwendige Austrocknung der Fruchtkörper verhindern (nachteilig zur Ausbreitung der Sporen) und zum anderen einen Befall durch Schimmelpilze begünstigen. Als weitere Faktoren gelten die relative Lichtarmut (Beeinträchtigung positiver Phototaxis), Windstille, saure Böden, eine Vielfalt von Fressfeinden und häufige starke Regenfälle, die die Zellen abwaschen oder zerstören können.[9] Vor allem die umfangreichen Gruppen der boden- und totholzbewohnenden Arten gehen mit zunehmender Feuchtigkeit zurück. In einer vergleichenden Studie aus Costa Rica stellen die Funde im relativ trockenen Tropical Moist Forest 73 % der Gesamtfunde, jene aus den bereits deutlich feuchteren Tropical Premontane Wet Forests nur noch 18 % und im Regenwald-Typ Lower Premontane Rain Forest sank er auf nur noch 9 % Anteil.[12]
Im südlichen Polarkreis existieren nur vereinzelte Nachweise von den Südlichen Shetlandinseln[13], den Südlichen Orkneyinseln, Südgeorgien sowie der Antarktischen Halbinsel.[14] Die im Vergleich zum arktischen Polarkreis ausgesprochen geringe Anzahl antarktischer bzw. subantarktischer Nachweise illustriert jedoch eher, wie unerforscht die Myxomycetenflora dieses Raumes ist. Bis 1983 waren nur fünf Funde bekannt[14], auch danach gab es nur einzelne weitere Funde.[13] Die beiden einzigen gründlichen Untersuchungen von Myxomycetenfloren der Region weisen jedoch auf im Allgemeinen deutlich höhere Artenzahlen hin. So konnten Stichproben aus subantarktischen Wäldern des argentinischen Patagoniens und Feuerland 67 Arten nachweisen,[15] eine Erhebung auf der nur 128 Quadratkilometer großen Macquarieinsel bereits 22.[16]
Habitate

Die Mehrheit aller bekannten Myxogastria-Arten lebt terrestrisch in offenen Wäldern. Das wichtigste Mikrohabitat ist Totholz, darüber hinaus von Bedeutung ist die Rinde lebender Bäume, verrottendes Pflanzenmaterial des Streuhorizonts, Erdboden und Tierexkremente.[17]
Davon abweichend finden sich Myxogastria an zahlreichen ungewöhnlichen Orten. Die umfangreiche Gruppe der sogenannten nivicolen Myxogastria wächst unter geschlossenen Schneedecken heran, um bei Exposition (zum Beispiel bei Tauwetter) schnell zu fruktifizieren und ihre Sporen freizugeben.[9] Auch Wüsten (für die Sonora-Wüste allein sind 33 Arten nachgewiesen[9]) oder lebende Blätter von Pflanzen in den Tropen[18] sind Habitate von Myxogastria.[17]
Ungewöhnlich ist die aquatische Lebensweise einiger Arten. Einzelne Vertreter der Gattungen Didymium, Physarum, Perichaena, Fuligo, Comatricha und Licea konnten sowohl als Myxoflagellaten wie als Plasmodien unter Wasser lebend beobachtet werden. Zur Fruktifikation kamen sie erst, wenn das Wasser zurückging oder sie es verließen, mit Ausnahme der Art Didymium difforme, für die ein Nachweis untergetauchter Fruktifikation erbracht wurde.[19]
Beziehungen zu anderen Lebewesen

Die Beziehungen der Myxogastria zu anderen Lebewesen sind bisher nur ansatzweise erforscht. Als Fressfeinde sind eine Reihe von Gliederfüßern bekannt, insbesondere Käfer (so aus den Familien der Kurzflügler,[20] Schwammkugelkäfer,[21] Runzelkäfer[22], Purzelkäfer,[23] Punktkäfer,[24] Schienenkäfer,[25] Staubpilzkäfer,[26] Glattrindenkäfer,[27] Moderkäfer[28]), Milben und Springschwänze. Auch verschiedene Nematoden haben sich als Fressfeinde erwiesen; sie verankern sich mit ihrem Hinterende in den Plasmasträngen von Plasmodien oder leben sogar innerhalb der Stränge.[29][2]
Bei den Zweiflüglern haben sich ebenfalls einige Spezialisten herausgebildet, meist Vertreter der Pilzmücken sowie der Trauermücken und der Taufliegen. Besonders häufig anzutreffen ist die Art Epicypta testata, sie findet sich vielfach an Enteridium lycoperdon, Enteridium splendens, Lycogala epidendrum und Tubifera ferruginosa.[30]
Einige echte Pilze haben sich auf die Besiedelung von Myxogastria spezialisiert, fast immer handelt es sich um Vertreter der Schlauchpilze. Am häufigsten findet man Verticillium rexianum (meist an Arten von Comatricha oder Stemonitis), an Arten der Physarida finden sich vielfach Gliocladium album oder Sesquicillium microsporum. An Vertretern der Trichiida findet sich oft Polycephalomyces tomentosus. Spezialisiert auf die Gelbe Lohblüte (Fuligo septica) ist Nectriopsis violacea.[30]
Als sogenannte „bakterielle Assoziierte“ fanden sich bei einigen Untersuchungen Bakterien meist aus der Familie der Enterobacteriaceae am Plasmodium lebend. Erst dem Verbund aus Plasmodien und bakteriellen Assoziierten ist es möglich, atmosphärischen Stickstoff zu binden oder Enzyme zu produzieren, die die Zersetzung zum Beispiel von Lignin, Carboxymethylcellulosen oder Xylan ermöglichen. In einzelnen Fällen erwarben die Plasmodien durch die Assoziation Salz- oder Schwermetalltoleranzen.[31]
Taxonomie
Nomenklatur
Der Name Myxogastria (=„Schleimbäuche“) wurde 1970 von Lindsay Shepherd Olive eingeführt, verweisend auf die bereits 1899 von Thomas Huston Macbride als Familie beschriebenen Myxogastridae.[32] Der Name taucht ähnlich bereits 1829 bei Elias Magnus Fries auf, der sämtliche Schleimpilze als „Myxogasteres“ bezeichnete.[33]
Durch konkurrierende Systematiken und den parallelen Gebrauch der einander ausschließenden botanischen wie zoologischen Regelwerke existieren viele verschiedene Namen ähnlichen oder gar identischen Schnitts (zum Beispiel Myxogastromycetidae, Mycetozoa, Myxomycota). Am bekanntesten ist sicher der 1833 von Johann Heinrich Friedrich Link erstbeschriebene Name „Myxomycetes“, der, obwohl als Taxon heute nicht mehr anerkannt, als „Myxomyceten“ zum Trivialnamen wurde und so in wissenschaftlichen Veröffentlichungen bis heute in Gebrauch ist. Der irreführenderweise eine Verwandtschaft zu den Pilzen andeutende Trivialname „Schleimpilze“ hingegen ist auf diese Gruppe falsch angewandt, da mit ihm das höhere (heute allerdings nicht mehr anerkannte) Taxon der Eumycetozoa bezeichnet wird.
Umfang
Die Gruppe umfasst rund 900 bis 1000 Arten. Eine auf Vollständigkeit angelegte Erhebung im Jahr 2000 ergab 1012 gültig beschriebene Taxa, darunter 866 auf Art-Ebene.[34] Eine andere Arbeit schätzte im Jahr 2007 die Artenzahl auf deutlich mehr als 1000, danach umfassten die Myxogastria als bei weitem größte Gruppe der Schleimpilze weit über 900 Arten. Die kontinuierliche Beschreibung neuer Taxa macht deutlich, dass die Kenntnis um die Gruppe noch immer unvollständig ist. Schätzungen anhand sequenzierter Umweltproben gehen davon aus, dass die Gruppe mit gesamt 1200–1500 Arten deutlich größer ist als bisher bekannt. Von den bekannten 1012 Taxa sind nur wenige Arten häufig: 305 sind von einem einzigen Ort oder nur einer einzigen Aufsammlung bekannt, 258 Arten wurden zwischen 2 und 20 mal an wenigen Orten gesammelt und nur 446 der Arten waren mit über 20 Funden an mehreren Standorten häufig.[34][35]
Zahlreiche Neubeschreibungen leiden darunter, dass Myxogastria durch Umwelteinflüsse morphologisch sehr beeinflussbar sind und zugleich nur wenige diagnostische Merkmale mit geringem Formenreichtum besitzen.[36] In Kombination mit der „problematischen Gepflogenheit“[6] von Autoren, bereitwillig anhand nur weniger zur Verfügung stehender Exemplare ein neues Taxon zu beschreiben, führt dies zu zahlreichen Dubletten (gelegentlich sogar auf Gattungsebene wie im Fall von Squamuloderma nullifila, eigentlich einer Art der Gattung Didymium[36]).[2]
Systematik und Phylogenie
Die folgende Systematik orientiert sich hinsichtlich der Myxogastria selbst an Adl et al. 2005,[3] in den Rängen und der weiteren Unterteilung nimmt sie jedoch die Systematik von Dykstra und Keller 2000 auf, in der die Myxogastria als „Mycetozoa“ angesprochen werden.[37]
Das Schwestertaxon bilden die Dictyostelia.[38] Mit ihnen und den Protostelia zusammen bildeten die Myxogastria nach klassischer Ansicht das Taxon der Schleimpilze (Eumycetozoa). Von den beiden anderen Gruppen ließen sich die Myxogastria vor allem durch den Weg der Ausbildung der Fruchtkörper trennen: während bei Protostelia aus jeder einzelnen und stets nur einkernigen Zelle ein separater Fruchtkörper entsteht, bildeten Dictyostelia aus einzelnen Zellen bestehende Zellverbände, sogenannte Pseudoplasmodien, die sich dann zu Fruchtkörpern umbauen. Nach neuerem Verständnis lässt sich das übergeordnete Taxon der Eumycetozoa nicht mehr aufrechterhalten.[39][8]
- Ordnung Liceida
- Familie Liceaceae
- Familie Enteridiidae
- Ordnung Echinosteliida
- Familie Echinosteliedae
- Familie Clastodermidae
- Ordnung Trichiida
- Familie Dianemidae
- Familie Trichiidae
- Ordnung Stemonitida
- Familie Stemonitidae
Kladogramm der Myxogastria[40]
|
- Ordnung Physarida
- Familie Elaeomyxidae
- Familie Physaridae
- Familie Didymiidae
Sie entspricht weitgehend der traditionellen Klassifikation, die seit den Arbeiten von Lister und Lister Anfang des 20. Jahrhunderts üblich war. Molekulargenetische Untersuchungen bestätigen diese Einteilung und festigen sie durch gut belastbare Resultate. Als basalste Gruppe gelten danach die Echinosteliida, die anderen Gruppen lassen sich darüber hinaus in zwei Superkladen fassen, die nach der Sporenfarbe auch morphologisch abgrenzbar sind.[41]
Fossilbericht
Fossile Funde, die den Myxogastria zugeordnet werden, sind ausgesprochen selten. Aufgrund ihrer kurzlebigen Natur und der empfindlichen Struktur der Plasmodien und Fruchtkörper kommen Myxogastria für Versteinerungen oder ähnliche Prozesse nicht in Frage, nur ihre Sporen können mineralisieren. Die wenigen bekannten Fossilien der Lebensstadien sind daher sämtlich nur als Einschlüsse in Bernstein bekannt.[42]
Bis zum Jahr 2010 wurden dreimal Fruchtkörper beschrieben, zweimal Sporen und einmal ein Plasmodium. Zwei ältere Taxa (Charles Eugène Bertrands Myxomycetes mangini und Bretonia hardingheni von 1892) gelten als zweifelhaft und werden in der Regel heutzutage nicht mehr berücksichtigt.[43][44][42]
Friedrich Walter Domke beschrieb 1952 einen 35 bis 40 Millionen Jahre alten Fund aus baltischem Bernstein der noch heute vorkommenden Art Stemonitis splendens. Bemerkenswert ist der Erhaltungszustand und die Vollständigkeit der Fruchtkörper, die eine annähernd einwandfreie Bestimmung ermöglichen. Aus derselben Periode und ebenfalls anhand baltischen Bernsteins wurde 2003 durch Heinrich Dörfelt und Alexander Schmidt Arcyria sulcata erstbeschrieben, eine Art, die der heutigen Arcyria denudata stark ähnelt. Beide Funde weisen darauf hin, dass sich die Fruchtkörper der Myxogastria in den letzten 35 bis 40 Millionen Jahren nur wenig verändert haben.[42]
Die 2006 wiederum von Dörfelt und Schmidt aus baltischem Bernstein beschriebene Protophysarum balticum hingegen gilt als fragwürdig. Nicht allein, dass sie der Gattungsbeschreibung in vielen Punkten widersprach und durch das Fehlen einer lateinischen Diagnose keine gültige Publikation darstellt. Schnell wurde ihr Charakter als Art der Myxogastria in Frage gestellt, da – anders als bei den bisherigen Funden – wichtige Details der Fruchtkörper nicht sichtbar waren oder der Zuordnung widersprachen. Darum wird heute eher davon ausgegangen, dass es sich bei dem Fund um eine Flechte aus dem Umfeld der Gattung Chaenotheca handelt. Der einzige bisher berichtete Fund eines Plasmodiums stammt aus dominikanischem Bernstein und wurde den Physarida zugeordnet. Auch er gilt jedoch als zweifelhaft, vor allem, da die Publikation durch Mängel als unzureichend eingestuft wird.[42]
Der bisher mit Abstand älteste 2019 publizierte Fund stammt aus Bernstein aus Myanmar, ist ungefähr 100 Millionen Jahre alt, und konnte der noch heute existierenden Gattung Stemonitis zugerechnet werden.[45]
Als einzig bekannte mineralisierte Fossilien sind 1971 zwei Sporenfunde bekannt geworden, von denen ein nacheiszeitlicher als Trichia favoginea nahestehend eingestuft wurde. Die Anregung zur Aufnahme von Myxogastria-Sporen in Pollen- und Sporenanalysen blieb jedoch ohne Widerhall.[43]
Forschungsgeschichte

Aufgrund ihrer unscheinbaren Natur sind die Myxogastria erst spät gezielt erforscht worden. Erstmals erwähnt Thomas Panckow 1654 in seinem Herbarium Portatile, oder behendes Kräuter- und Gewächsbuch von einer Illustration begleitet einen Myxogastria, nämlich den Blutmilchpilz, unter dem Namen „Fungus cito crescentes“, versteht ihn also als Pilz. 1729 formulierte Pier Antonio Micheli dann erstmals den Gedanken der bis dato bekannten Arten als eine von den Pilzen zu unterscheidende Gruppe von Lebewesen, dem Heinrich Friedrich Link ab 1833 zur Durchsetzung verhalf. Bereits 1829 hatte Elias Magnus Fries das plasmodiale Stadium dokumentiert, Anton de Bary konnte dann 1864 das Stadium der Sporenkeimung beobachten. Da er zugleich auch die Bewegung ermöglichende Plasmaströmung in der Zelle beobachten konnte, sah er sie als den Tieren näherstehend und beschrieb sie demzufolge neu als Mycetozoa, wörtlich übersetzt „Pilztiere“, eine Ansicht, die bis in die zweite Hälfte des 20. Jahrhunderts dominierte.[2]
Von 1874 bis 1876 veröffentlichte Jósef Tomasz Rostafinski, ein Schüler De Barys, die erste umfangreiche Monographie zur Gruppe. 1894, 1911 sowie 1925 erschienen schließlich von Arthur und Guilielma Lister drei Monographien, die bis heute als „beispielhafte, grundlegende Arbeiten“ rezipiert werden, dasselbe gilt für das 1934 erschienene The Myxomycetes von Thomas H. Macbride und George Willard Martin. Wichtige Arbeiten des späten 20. Jahrhunderts waren die Monographien von George Willard Martin und Constantine John Alexopoulos 1969 sowie Lindsay S. Olive 1975. Insbesondere erstere gilt als bedeutend, da durch sie „die moderne Ära der Taxonomie der Myxogastria begann“.[34] Zur Kenntnis neuer Taxa und ihrer Systematik trugen insbesondere Persoon, Rostafinski, Lister, Macbride sowie Martin und Alexopoulos bei, ihre Arbeiten erhöhten die jeweiligen Artenzahlen maßgeblich.[34][2]
Neben den die Gruppe weltweit behandelnden Monographien sind darüber hinaus einige Großraumfloren von Bedeutung, insbesondere, da die Gruppe wegen ihrer disjunkten Verbreitung wesentlich weniger regional wirksame Beschränkungen aufweist. Wichtige Großraumfloren sind z. B. Robert Hagelsteins The Mycetozoa of North America (1944) und Marie Farrs Band für die Flora Neotropica 1973, neuere Werke dieser Art sind Bruce Ings The Myxomycetes of Britain and Ireland, Lado und Pandos Band für die Flora Mycologica Iberica von 1997 und Yamamotos The Myxomycete Biota of Japan von 1998. Ungewöhnlich ist die Entstehung des dreibändigen Werkes Die Myxomyceten Deutschlands und des angrenzenden Alpenraumes unter besonderer Berücksichtigung Österreichs, das von 1993 bis 2000 durch Hermann Neubert, Wolfgang Nowotny und Karlheinz Baumann verfasst und im Eigenverlag veröffentlicht wurde. Obwohl die Autoren formal Amateure waren, erfuhr ihr Werk sehr positive Kritiken durch Wissenschaftler und ist heute ein vielzitiertes Standardwerk. Im Jahr 2011 erschien das Werk Les Myxomycètes, bestehend aus einem Text- und einem Bildband. Die Autoren sind Michel Poulain, Marianne Meyer und Jean Bozonnet. Auch dieses Werk erlangte bereits kurze Zeit nach seiner Erscheinung den Status eines Standardwerkes.
Nutzung
Für den Menschen sind die Myxogastria weitgehend ohne wirtschaftliche oder kulturelle Bedeutung. Physarum polycephalum dient in der Biologie als häufiger Modellorganismus zur Untersuchung von Zellmotilität, Zellwachstum und Zelldifferenzierung. Vorteilhaft für die Handhabung sind die leichte Kultivierbarkeit und die Größe der Zelle.[46]
Plasmodien der Gelben Lohblüte (Fuligo septica) und Aethalien von Enteridium lycoperdon werden in der Gegend von Veracruz in Mexiko gegessen. Dort sind sie gegrillt unter der Bezeichnung „caca de luna“, zu deutsch „Mondkacke“, als Delikatesse bekannt.[30]
Nachweise
- H. O. Schwantes: Biologie der Pilze. Stuttgart 1996, ISBN 3-8252-1871-6, S. 308–311
- Beleg für den Absatz: Wolfgang Nowotny: Myxomyceten (Schleimpilze) und Mycetozoa (Pilztiere) – Lebensformen zwischen Tier und Pflanze In: Wolfgang Nowotny (Hrsg.): Wolfsblut und Lohblüte. Lebensformen zwischen Tier und Pflanze = Myxomycetes (= Stapfia. Band 73). Linz 2000, ISBN 3-85474-056-5, S. 7–37 (deutsch, englisch, französisch, spanisch). (zobodat.at [PDF]).
- Sina M. Adl, Alastair G. B. Simpson, Mark A. Farmer, Robert A. Andersen, O. Roger Anderson, John A. Barta, Samuel S. Bowser, Guy Brugerolle, Robert A. Fensome, Suzanne Fredericq, Timothy Y. James, Sergei Karpov, Paul Kugrens, John Krug, Christopher E. Lane, Louise A. Lewis, Jean Lodge, Denis H. Lynn, David G. Mann, Richard M. McCourt, Leonel Mendoza, Øjvind Moestrup, Sharon E. Mozley-Standridge, Thomas A. Nerad, Carol A. Shearer, Alexey V. Smirnov, Frederick W. Spiegel, Max F. J. R. Taylor: The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. The Journal of Eukaryotic Microbiology 52 (5), 2005; S. 399–451 (The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. Abgerufen am 28. Dezember 2019.)
- H. Raven, Ray F. Evert, Helena Curtis: Biologie der Pflanzen. 2. Auflage. Walter de Gruyter, 1988, ISBN 3-11-011476-3, S. 267–269.
- Beleg für den Absatz: Henry Stempen, Steven L. Stevenson: Myxomycetes. A Handbook of Slime Molds. Timber Press, 1994, ISBN 0-88192-439-3, S. 15–18.
- Wolfgang Nowotny: Myxomyceten (Schleimpilze) und Mycetozoa (Pilztiere) – Lebensformen zwischen Tier und Pflanze In: Wolfgang Nowotny (Hrsg.): Wolfsblut und Lohblüte. Lebensformen zwischen Tier und Pflanze = Myxomycetes (= Stapfia. Band 73). Linz 2000, ISBN 3-85474-056-5, S. 7–37 (deutsch, englisch, französisch, spanisch). (zobodat.at [PDF]).
- Wolfgang Richter: Alter Schleimer. In: ZEIT Wissen. Nr. 1, 2007 (Online).
- Beleg für den Absatz: Sina M. Adl, Alastair G. B. Simpson, Mark A. Farmer, Robert A. Andersen, O. Roger Anderson, John A. Barta, Samuel S. Bowser, Guy Brugerolle, Robert A. Fensome, Suzanne Fredericq, Timothy Y. James, Sergei Karpov, Paul Kugrens, John Krug, Christopher E. Lane, Louise A. Lewis, Jean Lodge, Denis H. Lynn, David G. Mann, Richard M. McCourt, Leonel Mendoza, Øjvind Moestrup, Sharon E. Mozley-Standridge, Thomas A. Nerad, Carol A. Shearer, Alexey V. Smirnov, Frederick W. Spiegel, Max F. J. R. Taylor: The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. The Journal of Eukaryotic Microbiology 52 (5), 2005; S. 399–451 (The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. Abgerufen am 28. Dezember 2019.)
- Henry Stempen, Steven L. Stevenson: Myxomycetes. A Handbook of Slime Molds. Timber Press, 1994, ISBN 0-88192-439-3, S. 49–58.
- Steven L. Stephenson, Martin Schnittler, Carlos Lado: Ecological characterization of a tropical myxomycete assemblage—Maquipucuna Cloud Forest Reserve, Ecuador In: Mycologia, Bd. 96, S. 488–497, 2004
- Beleg für den Absatz: Steven L. Stephenson, Yuri K. Novozhilov, Martin Schnittler: Distribution and Ecology of Myxomycetes in High-Latitude Regions of the Northern Hemisphere In: Journal of Biogeography, 27:3, 2000, S. 741–754
- Beleg für den Absatz: Steven L. Stephenson, Martin Schnittler, Carlos Lado: Ecological characterization of a tropical myxomycete assemblage—Maquipucuna Cloud Forest Reserve, Ecuador In: Mycologia, Bd. 96, S. 488–497, 2004
- J. Putzke, A. B. Pereira, M. T. L. Putzke: New Record of Myxomycetes to the Antarctica. In: V Simposio Argentino y I Latinoamericano de Investigaciones Antarticas, 2004, Buenos Aires – Argentina, Actas del V Simposio Argentino y I Latinoamericano de Investigaciones Antarticas, Bd. 1, S. 1–4, 2004, PDF online (Memento vom 14. Oktober 2012 im Internet Archive)
- B. Ing, R. I. L. Smith: Further myxomycete records from South Georgia and the Antarctic peninsula. In: British Antarctic Survey Bulletin 59, S. 80–81, 1983
- Diana W. de Basanta, Carlos Lado, Arturo Estrada-Torres, Steven L. Stephenson: Biodiversity of myxomycetes in subantarctic forests of Patagonia and Tierra del Fuego, Argentina In: Nova Hedwigia, 90:1-2, S. 45–79
- S.L. Stephenson, G.A. Laursen, R.D. Seppelt: Myxomycetes of subantarctic Macquarie Island. In: Australian Journal of Botany 55, S. 439–449
- Beleg für den Absatz: Henry Stempen, Steven L. Stevenson: Myxomycetes. A Handbook of Slime Molds. Timber Press, 1994, ISBN 0-88192-439-3, S. 49–58.
- Uno H. Eliasson: Myxomyceten auf lebenden Blättern im tropischen Regenwald Ecuadors; eine Untersuchung basierend auf dem Herbarmaterial höherer Pflanzen In: Wolfgang Nowotny (Hrsg.): Wolfsblut und Lohblüte. Lebensformen zwischen Tier und Pflanze = Myxomycetes (= Stapfia. Band 73). Linz 2000, ISBN 3-85474-056-5, S. 81 (deutsch, englisch, französisch, spanisch). (zobodat.at [PDF]).
- Beleg für den Absatz: Lora A. Lindley, Steven L. Stephenson, Frederick W. Spiegel: Protostelids and myxomycetes isolated from aquatic habitats In: Mycologia, Bd. 99, S. 504–509, 2007
- Rolf G. Beutel, Richard A. B. Leschen (Hrsg.): Coleoptera, Beetles (= Handbuch der Zoologie. Band 4: Arthropoda: Insecta). 1. Auflage. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). de Gruyter, 2005, ISBN 3-11-017130-9, ISSN 1861-4388, S. 304 (englisch).
- Rolf G. Beutel, Richard A. B. Leschen (Hrsg.): Coleoptera, Beetles (= Handbuch der Zoologie. Band 4: Arthropoda: Insecta). 1. Auflage. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). de Gruyter, 2005, ISBN 3-11-017130-9, ISSN 1861-4388, S. 269 (englisch).
- Rolf G. Beutel, Richard A. B. Leschen (Hrsg.): Coleoptera, Beetles (= Handbuch der Zoologie. Band 4: Arthropoda: Insecta). 1. Auflage. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). de Gruyter, 2005, ISBN 3-11-017130-9, ISSN 1861-4388, S. 147 (englisch).
- Rolf G. Beutel, Richard A. B. Leschen (Hrsg.): Coleoptera, Beetles (= Handbuch der Zoologie. Band 4: Arthropoda: Insecta). 1. Auflage. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). de Gruyter, 2005, ISBN 3-11-017130-9, ISSN 1861-4388, S. 433 (englisch).
- Rolf G. Beutel, Richard A. B. Leschen (Hrsg.): Coleoptera, Beetles (= Handbuch der Zoologie. Band 4: Arthropoda: Insecta). 1. Auflage. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). de Gruyter, 2005, ISBN 3-11-017130-9, ISSN 1861-4388, S. 438 (englisch).
- Richard A. B. Leschen, Rolf G. Beutel, John F. Lawrence (Hrsg.): Coleoptera, Beetles (= Handbook of Zoology. Arthropoda: Insecta). Band 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). de Gruyter, 2010, ISBN 978-3-11-019075-5, S. 61 (englisch).
- Richard A. B. Leschen, Rolf G. Beutel, John F. Lawrence (Hrsg.): Coleoptera, Beetles (= Handbook of Zoology. Arthropoda: Insecta). Band 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). de Gruyter, 2010, ISBN 978-3-11-019075-5, S. 300 (englisch).
- Richard A. B. Leschen, Rolf G. Beutel, John F. Lawrence (Hrsg.): Coleoptera, Beetles (= Handbook of Zoology. Arthropoda: Insecta). Band 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). de Gruyter, 2010, ISBN 978-3-11-019075-5, S. 424 (englisch).
- Richard A. B. Leschen, Rolf G. Beutel, John F. Lawrence (Hrsg.): Coleoptera, Beetles (= Handbook of Zoology. Arthropoda: Insecta). Band 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). de Gruyter, 2010, ISBN 978-3-11-019075-5, S. 483 (englisch).
- Courtney M. Kilgore, Harold W. Keller: Interactions Between Myxomycete Plasmodia and Nematodes. In: Inoculum. Band 59, Nr. 1, 2008, S. 1–3.
- Beleg für den Absatz: Henry Stempen, Steven L. Stevenson: Myxomycetes. A Handbook of Slime Molds. Timber Press, 1994, ISBN 0-88192-439-3, S. 59–68.
- Beleg für den Absatz: Indira Kalyanasundaram: A Positive Ecological Role for Tropical Myxomycetes in Association with Bacteria In: Systematics and Geography of Plants, 74:2, S. 239–242, 2004
- Lindsay S. Olive: The Mycetozoa: A Revised Classification, In: Botanical Review, 36:1, S. 59–89, 1970
- Hermann Neubert, Wolfgang Nowotny, Karlheinz Baumann: Die Myxomyceten Deutschlands und des angrenzenden Alpenraumes unter besonderer Berücksichtigung Österreichs. Band 1. Karlheinz Baumann Verlag, Gomaringen 1993, ISBN 3-929822-00-8, S. 11.
- M. Schnittler & D. W. Mitchell: Species Diversity in Myxomycetes Based on the Morphological Species Concept – a Critical Examination In: Wolfgang Nowotny (Hrsg.): Wolfsblut und Lohblüte. Lebensformen zwischen Tier und Pflanze = Myxomycetes (= Stapfia. Band 73). Linz 2000, ISBN 3-85474-056-5, S. 39–53 (deutsch, englisch, französisch, spanisch).
- Beleg für den Absatz: Sina M. Adl, Brian S. Leander, Alastair G. B. Simpson, John M. Archibald, O. Roger Anderson, David Bass, Samuel S. Bowser, Guy Brugerolle, Mark A. Farmer, Sergey Karpov, Martin Kolisko, Christopher E. Lane, Deborah J. Lodge, David G. Mann, Ralf Meisterfeld, Leonel Mendoza, Øjvind Moestrup, Sharon E. Mozley-Standridge, Alexey V. Smirnov, Frederick Spiegel: Diversity, Nomenclature, and Taxonomy of Protists. Systematic Biology, Band 56, 2007, S. 684–689.
- Jim Clark: The species problem in the myxomycetes In: Wolfgang Nowotny (Hrsg.): Wolfsblut und Lohblüte. Lebensformen zwischen Tier und Pflanze = Myxomycetes (= Stapfia. Band 73). Linz 2000, ISBN 3-85474-056-5, S. 39–53 (deutsch, englisch, französisch, spanisch). (zobodat.at [PDF]).
- Michael J. Dykstra, Harold W. Keller: Mycetozoa In: John J. Lee, G. F. Leedale, P. Bradbury (Hrsg.): An Illustrated Guide to the Protozoa. Band 2. Allen, Lawrence 2000, ISBN 1-891276-23-9, S. 952–981.
- Anna Maria Fiore-Donno, Sergey I. Nikolaev, Michaela Nelson, Jan Pawlowski, Thomas Cavalier-Smith, Sandra L. Baldauf: Deep Phylogeny and Evolution of Slime Moulds (Mycetozoa) In: Protist, 161:1, S. 55–70, 2010
- Adl, S. M., Simpson, A. G. B., Lane, C. E., Lukeš, J., Bass, D., Bowser, S. S., Brown, M. W., Burki, F., Dunthorn, M., Hampl, V., Heiss, A., Hoppenrath, M., Lara, E., le Gall, L., Lynn, D. H., McManus, H., Mitchell, E. A. D., Mozley-Stanridge, S. E., Parfrey, L. W., Pawlowski, J., Rueckert, S., Shadwick, L., Schoch, C. L., Smirnov, A. and Spiegel, F. W.: The Revised Classification of Eukaryotes. Journal of Eukaryotic Microbiology, 59: 429–514, 2012, PDF Online
- A.-M. Fiore-Donno, C. Berney, J. Pawlowski, S.L. Baldauf: Higher-Order Phylogeny of Plasmodial Slime Molds (Myxogastria) Based on Elongation Factor 1-A and Small Subunit rRNA Gene Sequences. In: Journal of Eukaryotic Microbiology, 52, S. 201–210, 2005
- Beleg für den Absatz: A.-M. Fiore-Donno, C. Berney, J. Pawlowski, S.L. Baldauf: Higher-Order Phylogeny of Plasmodial Slime Molds (Myxogastria) Based on Elongation Factor 1-A and Small Subunit rRNA Gene Sequences. In: Journal of Eukaryotic Microbiology, 52, S. 201–210, 2005
- Harold W. Keller, Sydney E. Everhart: Myxomycete species concepts, monotypic genera, the fossil record, and additional examples of good taxonomic practice In: Revista Mexicana de Micología, 27, 2008, S. 9–19
- Beleg für den Absatz: Alan Graham: The role of myxomyceta spores in palynology (with a brief note on the morphology of certain algal zygospores) In: Review of Palaeobotany and Palynology, 11:2, S. 89–99, 1971
- B.M. Waggoner, G.O. Poinar: A Fossil Myxomycete Plasmodium from Eocene-Oligocene Amber of the Dominican Republic. In: Journal of Eukaryotic Microbiology, 39, S. 639–642, 1992
- Jouko Rikkinen, David A. Grimaldi & Alexander R. Schmidt: Morphological stasis in the first myxomycete from the Mesozoic, and the likely role of cryptobiosis. In: Scientific Reports. Band 9, 2019, S. 19730, doi:10.1038/s41598-019-55622-9.
- Beleg für den Absatz: Henry Stempen, Steven L. Stevenson: Myxomycetes. A Handbook of Slime Molds. Timber Press, 1994, ISBN 0-88192-439-3, S. 19–21.
Weiterführende Literatur
- Hermann Neubert, Wolfgang Nowotny, Karlheinz Baumann, Heidi Marx: Die Myxomyceten Deutschlands und des angrenzenden Alpenraumes unter besonderer Berücksichtigung Österreichs. Bd. 1–3, Karlheinz Baumann Verlag, Gomaringen.
- George W. Martin, Constantine John Alexopoulos, Marie Leonore Farr: The genera of Myxomycetes, Iowa City, 1983, ISBN 0-87745-124-9.
- Wolfgang Nowotny (Hrsg.): Wolfsblut und Lohblüte. Lebensformen zwischen Tier und Pflanze = Myxomycetes (= Stapfia. Band 73). Linz 2000, ISBN 3-85474-056-5 (deutsch, englisch, französisch, spanisch). Online (19 PDF-Dateien): Wolfsblut und Lohblüte. – Lebensformen zwischen Tier und Pflanze. In: ZOBODAT.at. OÖ Landes-Kultur GmbH.
- Henry Stempen, Steven L. Stevenson: Myxomycetes. A Handbook of Slime Molds. Timber Press, 1994, ISBN 0-88192-439-3.