Flagellum
Flagellen (lateinisch flagellum) oder Geißeln sind fadenförmige Gebilde auf der Oberfläche einzelner Zellen, die der Fortbewegung dienen. Sie sind bei Prokaryoten (Lebewesen ohne Zellkern) und bei Eukaryoten (Lebewesen mit Zellkern) hinsichtlich Struktur und Funktionsweise grundsätzlich verschieden:
- Prokaryoten besitzen gewendelte Proteinfäden außerhalb der Zellmembran, die sich nicht aktiv verformen, sondern an ihrem in der Zelle verankerten Ende durch einen Motor in Drehung versetzt werden und auf diese Weise – ähnlich wie ein Propeller – einen Schub oder Zug ausüben.
- Eukaryoten dagegen besitzen fädige, von der Zellmembran umschlossene Ausstülpungen der Zelle, in deren Inneren sich ein Bündel von Mikrotubuli befindet, und die durch aktive Formveränderung eine Bewegung bewirken.
Diese beiden völlig unterschiedlichen Organelltypen werden jeweils sowohl als Flagellen als auch als Geißeln bezeichnet, diese beiden Bezeichnungen werden also als Synonyme verwendet. Um jedoch dem unterschiedlichen Aufbau und der verschiedenen Funktion von Pro- und Eukaryotengeißeln nomenklatorisch Rechnung zu tragen, ist von deutschsprachigen Autoren folgende Sprachregelung vorgeschlagen worden:
„Die Geißeln der eukaryotischen Zellen haben einen sehr einheitlichen Aufbau. […] Der Begriff 'Flagelle' bleibt den ganz anders organisierten Fortbewegungsorganellen der Prokaryoten vorbehalten.“[1]
Im vorliegenden Artikel wird diese eindeutige Zuordnung der Bezeichnungen, also „Flagellum“ für Prokaryoten und „Geißel“ für Eukaryoten, weitgehend berücksichtigt (mit Ausnahme feststehender Begriffe wie Begeißelung). Zu bemerken ist aber, dass in anderen aktuellen Lehrbüchern diese Unterscheidung nicht gemacht wird.[2]
Die Flagellen der Prokaryoten
Aufbau
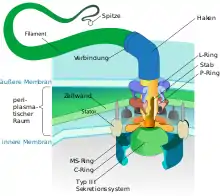
Bakterielle Flagellen sind extrazelluläre, wendelförmige Fäden („Filamente“), die über einen „Haken“ mit einem Motorkomplex in der Zellmembran (bzw. den Zellmembranen) und der Zellwand verankert sind. Die Flagellen einschließlich Haken und Motorkomplex bestehen vollständig aus Proteinen. Der Durchmesser der Filamente beträgt bei den meisten Flagellen etwa 15–20 nm, und sie sind hohl. Wegen ihres geringen Durchmessers sind sie nur mit Dunkelfeldmikroskopie und Elektronenmikroskopie sichtbar zu machen, mit normaler Lichtmikroskopie nicht; es gibt jedoch spezielle Färbeverfahren, durch die sie so weit verdickt werden, dass sie lichtmikroskopisch sichtbar werden.
Beim Aufbau der Filamente werden die Proteinmoleküle (Flagellin) durch den Hohlkanal der Flagellen bis zum äußeren Ende transportiert und dort angebaut. Falls ein ausreichend großer Vorrat an Flagellin in der Zelle vorhanden ist, kann der Aufbau eines Filaments sehr schnell geschehen.
Die Flagellen der Archaeen (auch Archaellen, im Singular Archaellum genannt) sind funktionell ähnlich aufgebaut wie die der Bakterien, bestehen jedoch aus anderen Proteinen und einem unterschiedlichen Motorkomplex, der mit ATP angetrieben wird. Beispiele finden sich bei Haloarcula marismortui und Sulfolobus acidocaldarius, Pyrococcus furiosus und Methanocaldococcus villosus.[3]
Begeißelungstypen
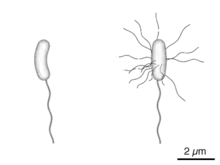
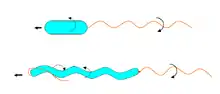
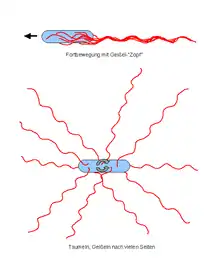
Nach Anordnung und Anzahl der Flagellen werden verschiedene Begeißelungstypen unterschieden (in der Reihenfolge absteigender Schwimmgeschwindigkeit):
- holotrich: Zahlreiche Flagellen sind gleichmäßig über die gesamte Zelloberfläche verteilt und umfassen die gesamte Körperoberfläche.
- peritrich: Viele Flagellen sind gleichmäßig über die Zelloberfläche verstreut.
- polytrich-bipolar: Die Flagellen stehen in zwei gegenüberliegenden Gruppen an den Zellpolen (auch als amphitrich bezeichnet).
- polytrich-monopolar: Die Flagellen stehen in einer Gruppe an einem der Zellpole (auch als lophotrich bezeichnet).
- monotrich: Die Zelle hat nur eine einzige Flagelle.
- polar: Das Flagellum bzw. die Flagellen stehen an einem oder beiden Polen der Zelle.
- lateral, seitliche Begeißelung: Flagellen stehen seitlich, nicht an den Polen der Zelle.
Die laterale Begeißelung ist oft nicht mit hoher Schwimmgeschwindigkeit verbunden, sie bietet aber den Vorteil, dass sich das Bakterium leichter in Hindernisse, wie hochviskose Flüssigkeiten oder Lücken zwischen Feststoffen, zwängen kann.
Bewegungsweise
Die Flagellen wirken durch ihre Wendelung ähnlich wie ein Propeller. Der Motorkomplex setzt einen Konzentrationsunterschied an Protonen zwischen den beiden Seiten der inneren Zellmembran in eine Drehbewegung des auf einem gekrümmten „Haken“ sitzenden gewendelten Filaments um und folgt damit einem ähnlichen Bauprinzip wie die ATP-Synthase. Die Flagellenmechanik stellt das bisher einzig bekannte echt rotierende Gelenk in der gesamten Biologie dar. Die Drehfrequenz liegt um 40–50 Hz.
Die Richtung der durch den Motor bewirkten Flagellen-Drehung in Kombination mit der Windungsrichtung der Flagellen-Wendel bestimmt, ob ein Schub oder ein Zug auf den Bakterienkörper ausgeübt wird. Die Richtung der durch den Motor bewirkten Drehung kann in sehr kurzer Zeit umgekehrt werden, so dass Schub und Zug schnell wechseln können.
In der Regel ist die Drehrichtung der Flagellen so, dass sie schieben. Das bedeutet, dass sie sich bei monopolar begeißelten Bakterien am Hinterende befinden. Der Bakterienkörper dreht sich dabei (langsamer) in entgegengesetzter Richtung (Erhaltung des Drehimpulses).
Bei bipolar begeißelten Bakterien drehen sich die Flagellen der beiden Enden gegensinnig. Dadurch wirken die Flagellen des Hinterendes schiebend, die Flagellen des Vorderendes sind nach hinten gebogen und drehen sich um das Vorderende des Bakterienkörpers und verstärken so den Schub. Wird die Drehrichtung der Flagellen umgekehrt, klappen die Filamente um, das Hinterende des Bakteriums wird zum Vorderende und das Vorderende zum Hinterende, das Bakterium schwimmt in die Gegenrichtung.
Die Flagellen peritrich begeißelter Bakterien drehen gleichsinnig und zwar in der Regel so, dass sie schieben. Dabei vereinigen sie sich zu einem nach hinten gerichteten gewendelten Bündel, auch als „Geißelzopf“ bezeichnet, welches das Bakterium vorwärts schiebt. Wird die Drehrichtung der Flagellen peritrich begeißelter Bakterien umgekehrt, richten sich die einzelnen Flagellen radial vom Bakterienkörper abstehend und ihre Zugwirkung auf den Bakterienkörper hebt sich im Mittel auf, wodurch das Bakterium in zufälliger Bewegung an einem Ort taumelt.
Die Umkehrung der Flagellen-Drehrichtung und die damit verbundene Änderung der Bewegungsrichtung spielt bei Taxien eine bedeutende Rolle (siehe beispielsweise Chemotaxis).
Die Geißeln der Eukaryoten
Aufbau
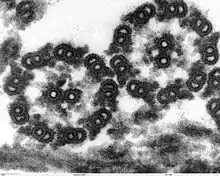
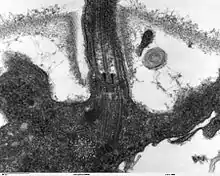
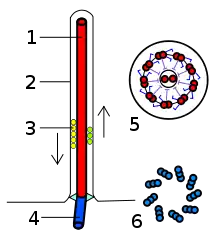
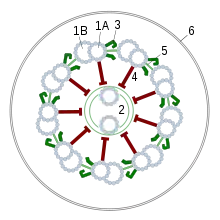
Die Geißeln der Eukaryoten sind fadenförmige Gebilde, die vom Körper nach außen in das umgebende Medium ragen; sie sind von der Zellmembran umgeben und von Zellplasma erfüllt. In ihrem Inneren liegen Mikrotubuli in einer speziellen Anordnung, der sogenannten 9×2+2: Neun doppelte Mikrotubuli bilden im Querschnitt einen Kreis, in dessen Mitte zwei einzelne Mikrotubuli liegen. Die doppelten Mikrotubuli bestehen aus je einem vollständigen Mikrotubulus (A-Tubulus) und einem unvollständigen Mikrotubulus (B-Tubulus). Am A-Tubulus befinden sich in gleichem Höhenabstand, etwa alle 20 nm, Paare von Protein-Armen, die als Dyneinarme bezeichnet werden. Das gesamte Mikrotubuli-Bündel wird als Axonem bezeichnet. Diese Struktur wird von verschiedenen Brückenproteinen stabilisiert (vor allem Nexin). An der Basis der Geißel, wo sie in den Zellkörper übergeht, befindet sich ein Basalapparat; dieser wird als Blepharoplast oder Kinetosom bezeichnet, oft allerdings auch als Centriol, da er strukturell einem solchen gleicht. Er besteht aus neun dreifachen Mikrotubuli in einem Kreis (9×3-Struktur), der quer zu einer zweiten, gleich strukturierten 9×3-Struktur liegt. An ihrem freien Ende sind Eukaryoten-Geißeln zugespitzt. Ihr Durchmesser beträgt etwa 250–300 nm, ihre Länge wenige Mikrometer bis mehr als 150 µm.
Ein anschauliches Beispiel für geißeltragende Zellen stellen Spermatozoen dar. Bei ihnen geht die Bewegung in einer Welle mit gleichbleibender Amplitude von der Basis zur Spitze der Geißel.
Gemeinsam mit den Zilien bezeichnet man die Geißeln der Eukaryoten auch als Undulipodien.
Bewegungsweise
Die zur hydrodynamischen Wirksamkeit erforderliche Formveränderung kommt nach dem bisherigen Wissensstand durch gegeneinander gerichtetes Gleiten der Doppelfibrillen zustande. Die Energie dafür soll von den Dyneinarmen bereitgestellt werden, und zwar durch hydrolytische Abspaltung von Phosphat aus ATP. Das die Dyneinarme bildende Protein Dynein hat ATPase-Aktivität. Das Mikrotubuli-Gleiten hat eine Formveränderung der Geißel zur Folge.
Die Formveränderungen der Geißeln sind je nach Geißeltyp verschieden. Sie können in einer über die Geißel fortlaufenden Welle (Undulation) in einer Ebene oder in Form einer Wendel mit kreis- bis ellipsenförmigen Bewegungen bestehen, sie können auch in einem Geißelschlag bestehen, wobei die Geißel sich in der einen Richtung krümmt und gewissermaßen das Medium unterwandert und in der Gegenrichtung ausgestreckt schlägt und damit eine Kraft ausübt. Geißeln, die sich in der zuletzt geschilderten Weise bewegen, werden Zilien genannt. Sie sind meistens kürzer als andere Geißeln und in größerer Dichte auf der Oberfläche der Zellen und Gewebe angeordnet.
Die Folge der Geißelbewegung kann eine Fortbewegung des Individuums sein, sie kann jedoch auch bei ruhendem Individuum eine Bewegung des angrenzenden Mediums oder in der Nähe befindlicher Partikel zur Folge haben. Beispiele für die Fortbewegung des Individuums sind frei bewegliche Ciliaten, Flagellaten und Spermatozoen. Beispiele für die Fortbewegung des angrenzenden Mediums oder von Partikeln sind festsitzende Ciliaten und das Flimmerepithel in der Luftröhre von Tieren.
Variationen, Begeißelungstypen
Bei einigen einzelligen Algen und Protozoen sind die Geißeln mit vielen seitlichen kurzen Filamenten, sogenannten Mastigonemen oder Flimmern, besetzt und werden als Flimmergeißeln bezeichnet. Die Mastigonemen können in einer Reihe (stichonematisch) oder in zwei Reihen (pantonematisch) auftreten.
Trägt eine Zelle mehrere gleichartige Geißeln, so spricht man von einer isokonten Begeißelung, so zum Beispiel bei Grünalgen.[4] Verschiedenartig begeißelte Zellen werden als heterokont oder anisokont bezeichnet, wobei meist eine lange, nach vorne gerichtete Flimmergeißel als Zuggeißel dient und eine kurze glatte Schleppgeißel nach hinten gerichtet ist, so zum Beispiel bei den Heterokontae.[5] Zellen ohne Geißel werden in Abgrenzung zu den begeißelten Formen als akont bezeichnet.[6]
Nach dem Insertionsort der Geißel unterscheidet man akrokont (am Vorderende, Zuggeißel), pleurokont (seitlich) und opisthokont (am Hinterende, Schubgeißel).
Einzelnachweise
- Kleinig, Maier: Zellbiologie, 4. Auflage, 1999, S. 151.
- So wird z. B. in der deutschen Übersetzung der 3. Auflage von Alberts’ Essential Cell Biology (2005) das englische Original „flagella“ mit „Flagellen (Geißeln)“ übersetzt und aufgrund dieser Gleichsetzung die Geißeln der Eukaryoten im Folgetext als „Flagellen“ bezeichnet, vgl. ebd. S. 625 ff.
- Biologists Investigate Smallest Propeller on Earth – Used by One of the Fastest Organisms on the Planet. Auf SciTechDaily vom 12. Februar 2022. Quelle: University of Exeter.
- Isokont im Springer Lexikon der Biologie
- Heterokont im Springer Lexikon der Biologie
- Akont im Springer Lexikon der Biologie
Weblinks
- Protonic NanoMachine Project
- Zur Evolution des Bakterienmotors: Die Entstehung bakterieller Flagellen ist erklärbar
- Die bakterielle Flagelle: Hinweise zu molekularem Aufbau, Diversität und Evolution
- Die Evolution des bakteriellen Motors: aktueller Stand der Forschung zur Evolution