Anhinga pannonica
Anhinga pannonica ist eine ausgestorbene Art der Schlangenhalsvögel (Anhinga), die während des Mittleren und Oberen Miozäns weite Teile Europas und Afrikas bewohnte. Die Art ist der einzige Vertreter der Gattung, dessen Verbreitungsgebiet auch Europa umfasste.
| Anhinga pannonica | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
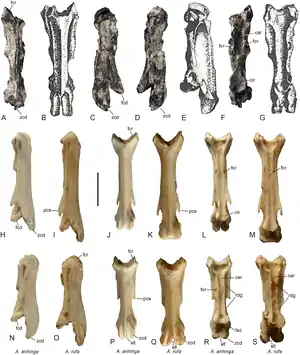
5. oder 6. Halswirbel von Anhinga pannonica (A–G; Holotypus B, E, G) im Vergleich zu 4. (H–M) und 6. Halswirbeln (N–S) rezenter Schlangenhalsvögel. | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittleres bis Oberes Miozän | ||||||||||||
| 16 bis 6 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Anhinga pannonica | ||||||||||||
| (Lambrecht, 1916) | ||||||||||||
Forschungsgeschichte und Etymologie
Die Erstbeschreibung der Art erfolgte 1916 durch den ungarischen Paläontologen Kálmán Lambrecht unter der Bezeichnung Plotus pannonicus.[1] Holotypus der Art ist ein sechster Halswirbel aus dem Tortonium (= Pannonium der regionalen chronostratigraphischen Gliederung der zentralen Paratethys) von Brusturi (Ungarisch: Tataros) im heutigen Kreis Bihor in Rumänien (ehemaliges Komitat Bihar im Königreich Ungarn). Ein Carpometacarpus von derselben Fundstelle wurde von Lambrecht ebenfalls der neuen Art zugeordnet.[1][2] Der von Lambrecht gewählte Artzusatz pannonicus bezieht sich auf die chronostratigraphische Einstufung der Fundschicht.[1]
Pierce Brodkorb stellte die Art 1963 als Anhinga pannonica in die Gattung Anhinga.[3][4]
Erst 1972 beschrieb Patricia Vickers-Rich weiteres Fossilmaterial (ein Halswirbel und ein unvollständiger Humerus) aus der Beglia-Formation Tunesiens als zu Anhinga pannonica gehörend.[5][2] Gleichzeitig legte sie den von Lambrecht 1916 beschriebenen Halswirbel von Brusturi als Lectotypus der Art fest, in der Annahme Lambrechts Erstbeschreibung hätte sich sowohl auf den Halswirbel als auch auf den Carpometacarpus der Typlokalität bezogen. Da sich Lambrechts Erstbeschreibung jedoch eindeutig auf den Halswirbel von Brusturi bezieht, war eine solche Festlegung unnötig und wurde von späteren Autoren als ungültig gewertet.[4]
Weitere europäische Funde, ein fast vollständiger rechter sowie das Fragment eines linken Humerus, stammen aus einer Sandgrube bei Götzendorf an der Leitha in Niederösterreich und wurden 1991 von Jiří Mlíkovský beschrieben.[6] Ein ähnlicher Fund wurde 2006 von der Fundstelle Hambach 6C des Tagebaus Hambach in der Niederrheinischen Bucht gemeldet.[2][7]
Im Jahr 2008 wurden, unabhängig voneinander, zwei Funde von großen Schlangenhalsvögeln von Toros Menalla im Tschad[8] und aus der Ngorora-Formation in Kenia[9] als vergleichbar mit Anhinga pannonica (Anhinga cf. pannonica) beschrieben.

Das bislang umfangreichste Belegmaterial stammt aus der Tongrube Hammerschmiede bei Pforzen im bayerischen Landkreis Ostallgäu. Die Funde umfassen postcraniale Skelettelemente von mindestens fünf Individuen und lassen sich den Fundhorizonten HAM 4 und HAM 5 der lokalen stratigraphischen Gliederung zuordnen. Diese Funde wurden 2020 durch Gerald Mayr, Thomas Lechner und Madelaine Böhme im Detail beschrieben. Gleichzeitig wurden die ursprünglich durch Ludwig von Ammon bereits 1918 als Kormoran (Phalacrocorax praecarbo) und Reiher (Ardea brunhuberi und Botaurites avitus) beschriebenen Fossilien[10] aus dem Mittleren Miozän von Regensburg-Dechbetten sowie die Überreste eines großen, zunächst nicht näher bestimmten Schlangenhalsvogels (Anhinga sp.)[11] aus dem obersten Miozän der Sahabi-Formation in Libyen ebenfalls Anhinga pannonica zugeordnet. Funde aus dem asiatischen Raum (Pakistan,[12] Thailand[13]), die ursprünglich als Anhinga pannonica oder damit vergleichbar beschrieben worden waren, wurden von den Autoren hingegen als nicht zu dieser Art gehörend gewertet.[2]
Zeitliche und geographische Verbreitung
(In Klammer eingefügte Ziffern beziehen sich auf nebenstehende Kartenskizze.)
Die beiden nördlichsten Fundstellen, Hambach (1) und Regensburg-Dechbetten (2) sind gleichzeitig auch die ältesten und lassen sich mit Altern von ~15 Ma beziehungsweise ~16 Ma in das frühe Mittlere Miozän datieren. Der südlichste Fundpunkt in Kenia (3) ist etwas jünger und fällt in das späte Mittlere Miozän.[2]
Die übrigen europäischen Fundorte, Hammerschmiede (4), Götzendorf (5) und Brusturi (6) datieren, ebenso wie die Funde aus Tunesien (7), in das frühe Obere Miozän, was einem Alter von etwa 11,6–9,8 Ma entspricht. Die beiden weiteren afrikanischen Funde aus Libyen (8) und dem Tschad (9) lassen sich mit Altern von 7–6 Ma hingegen dem Messinium, der obersten Stufe des Miozäns, zuordnen.[2]
Das geographische Verbreitungsgebiet erstreckt sich damit über etwa 50 Breitengrade, vom Äquator in Afrika bis in die Mittleren Breiten Europas. Die einzelnen Fossilbelege decken einen ungewöhnlich langen zeitlichen Rahmen von etwa 10 Millionen Jahren ab.[2]
Merkmale
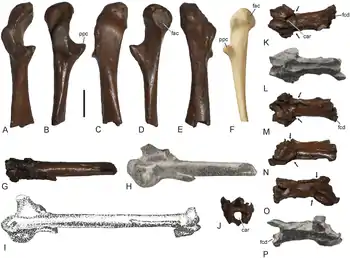

(In Klammer eingefügte Abkürzungen beziehen sich auf die Abbildungen rechts.)
Anhinga pannonica war ein relativ großer Vertreter der Schlangenhalsvögel. Alle hinreichend gut erhaltenen Skelettelemente der Gliedmaßen weisen Dimensionen auf, die jene der rezenten Anhinga-Arten um rund 15 % übertreffen und damit eher jenen eines Kormorans (Phalacrocorax carbo) entsprechen. Der kleinste Schaftumfang des Oberschenkelknochens lässt Rückschlüsse auf eine Körpermasse von etwa 3,3 kg zu. Der Wert übertrifft die Körpermasse von 1,0–1,8 kg der rezenten Schlangenhalsvögel zwar deutlich, bleibt jedoch weit hinter den Schätzwerten für einige ausgestorbene, südamerikanische Vertreter der Anhingidae zurück.[2]
Die fünften und/oder sechsten Halswirbel von Anhinga pannonica entsprechen in ihrer schmalen, langgestreckten Gestalt im Wesentlichen jenen der rezenten Schlangenhalsvögel. Die Rippenfortsätze (Processus costales; „pcs“) sind mit dem Wirbelkörper verwachsen und bilden entlang der ventrolateralen Seiten des Wirbelkörpers charakteristische, gratförmige Plattformen, die ein jeweils lateral liegendes Foramen („for“) des Wirbelkörpers begrenzen. Die Kombination dieser Merkmale gilt als diagnostische Apomorphie der Anhingidae.[2]
Das Rabenbein unterscheidet sich von dem der rezenten Schlangenhalsvögel in einigen Details, vor allem in Bezug auf die Knochenfortsätze an seinem oberen, zur Schulter hin gerichteten, Ende („Extremitas omalis“; nach dem altgriechischen ὦμος (ōmos) = „Schulter“[14]). Der Processus acrocoracoideus, ein craniomedial ausgerichteter, hakenförmiger Fortsatz,[14] ist dorsoventral schmaler und die Facies articularis clavicularis („fac“), die Gelenkfläche zum Schlüsselbein, ist sterno-omal (in Richtung zwischen Brustbein und Schulter) länger als bei den rezenten Arten. Die Facies articularis scapularis, die Gelenkfläche zum Schulterblatt, ist bei Anhinga pannonica leicht konkav gewölbt, während sie bei den rezenten Schlangenhalsvögeln weitgehend flach ausgeprägt ist. Der Processus procoracoideus („ppc“) ist verhältnismäßig lang und nicht knaufförmig, wie bei den rezenten Schlangenhalsvögeln, sondern spitz zulaufend. Der mediale Rand der Extremitas omalis des Rabenbeins bildet eine scharfe Kante. Als typisch Merkmal der rezenten Schlangenhalsvögel findet sich auch bei Anhinga pannonica auf der ventralen Seite der Extremitas omalis, unmittelbar schulterwärts zur Facies articularis clavicularis, eine deutlich ausgeprägte Vertiefung, die den nahe verwandten Kormoranen fehlt. Die Morphologie der Extremitas sternalis (zum Brustbein hin gerichtetes Ende des Rabenbeins) entspricht ebenfalls weitgehend jener der rezenten Schlangenhalsvögel.[2]
Die Ulna entspricht in ihren wesentlichen Merkmalen ebenfalls jenen der rezenten Schlangenhalsvögel, zeigt jedoch auch einige Abweichungen im Detail. Der Condylus dorsalis („cdd“; einer der Gelenkknorren am distalen Ende der Ulna, welche die Verbindung zu den Ossa carpi bilden) ist bei Anhinga pannonica etwas stärker konvex gewölbt als bei den rezenten Arten. Das Tuberculum carpale („tbc“), ein Knochenfortsatz im Bereich des zweiten Gelenkknorrens (Condylus ventralis; „cdv“) weist im Unterschied zu den rezenten Schlangenhalsvögeln einen eher geraden cranialen Rand auf. Der ganze Knochenfortsatz ist vergleichsweise schlank und eher spitz zulaufend. Dieses Merkmal unterscheidet Anhinga pannonica nicht nur von den rezenten Schlangenhalsvögeln, sondern auch von der ähnlich großen Art Anhinga grandis aus dem späten Miozän von Nordamerika.[2]
Dem Carpometacarpus scheint, im Gegensatz zu den rezenten Anhinga-Arten, ein charakteristischer, kammartiger Knochenfortsatz am distalen Ende des Os metacarpale majus zu fehlen.[2]
Abgesehen von der Größe entspricht auch der Oberschenkelknochen in seinen Proportionen weitgehend dem der rezenten Schlangenhalsvögel und ist damit deutlich langgestreckter als etwa der robuste Oberschenkelknochen der Gattung Macranhinga, die im Miozän Südamerikas ebenfalls mit riesenhafte Formen vertreten war.[2]
Palökologie

Anhinga pannonica ernährte sich vermutlich, ebenso wie die rezenten Schlangenhalsvögel, überwiegend von Fischen, die er tauchend erbeutete.[6] Zur Reduktion des Auftriebs besitzen rezente Schlangenhalsvögel ein stark benetzbares Gefieder, was sie andererseits jedoch anfällig für Wärmeverluste während und nach diesen Tauchgängen macht. Die Vögel verbringen deshalb viel Zeit mit dem Sonnenbaden in charakteristischer Pose mit weit abgespreizten Flügeln. Durch seine Größe und das damit günstigere Oberfläche-Volumen-Verhältnis war Anhinga pannonica diesbezüglich wohl im Vorteil.[2]
Rezente Schlangenhalsvögel bevorzugen tropische bis subtropische Süßwasserhabitate[2] und die europäischen Funde von Anhinga pannonica lassen sich in einem dementsprechenden paläoklimatologischen Zusammenhang interpretieren.
Die beiden ältesten Fossilbelege von Regensburg-Dechbetten (~16 Ma) und Hambach 6C (~15 Ma) fallen zeitlich noch in die Spätphase des Miozänen Klimaoptimums (~18–14,7 Ma[15][16]). Während dieser Zeit herrschten in Mitteleuropa tropische bis subtropische Bedingungen mit einer Jahresmitteltemperatur von bis zu 22 °C und durchschnittlichen Jahresniederschlagsmengen von mehr als 1200 mm.[15]
Die zweite Altersgruppe an europäischen Fossilbelegen (Hammerschmiede, Brusturi und Götzendorf) korreliert mit einer Phase, die als „Erstes Waschküchenklima“ („first washhouse climate“), mit subtropischen Temperaturen und durchschnittlichen Jahresniederschlagsmengen von mehr als dem Doppelten des heutigen Wertes,[17] bekannt ist. Das Verschwinden von Schlangenhalsvögeln in Mitteleuropa wird mit dem zunehmend kühleren Klima ab dem späteren Neogen in Zusammenhang gebracht. Die Gründe für das Aussterben von Anhinga pannonica im afrikanischen Teil des Verbreitungsgebietes sind hingegen unklar.[2]
Einzelnachweise
- K. Lambrecht: Die Gattung Plotus im ungarischen Neogen. In: Mitteilungen aus dem Jahrbuche der königlich ungarischen geologischen Anstalt, Band 24, 1916, S. 1–24 (Digitalisat).
- G. Mayr, T. Lechner & M. Böhme: The large-sized darter Anhinga pannonica (Aves, Anhingidae) from the late Miocene hominid Hammerschmiede locality in Southern Germany. In: PLoS ONE, Band 15, Nummer 5, 2020, Artikel e0232179, doi:10.1371/journal.pone.0232179.
- P. Brodkorb: Catalogue of Fossil Birds - Part 1 (Archaeopterygiformes through Ardeiformes). In: Bulletin of the Florida State Museum, Band 7, Nummer 4, 1963, S. 256 (Digitalisat).
- J. Mlíkovský: Cenozoic Birds of the World - Part 1: Europe. Ninox Press, Prag, 2002, ISBN 80-901105-3-8, S. 73 (Digitalisat).
- P. V. Rich: A fossil avifauna from the Upper Miocene Beglia formation of Tunisia. In: Notes du Service Géologique, Band 35, Nummer 5, 1972, S. 29–66.
- Jiri Mlíkovský: Late Miocene birds of Götzendorf/Leitha, Austria. In: Annalen des Naturhistorischen Museums in Wien, Band 92, Serie A, 1991, S. 97–100 (zobodat.at [PDF]).
- J. Dalsätt, T. Mörs & P. G. P. Ericson: Fossil birds from the Miocene and Pliocene of Hambach (NW Germany). In: Palaeontographica Abteilung A, Band 277, Lieferung 1–6, 2006, S. 113–121 (Abstract).
- A. Louchart, Y. Haile-Selassie, P. Vignaud, A. Likius & M. Brunet: Fossil birds from the Late Miocene of Chad and Ethiopia and zoogeographical implications. In: ORYCTOS, Band 7, 2008, S. 147–167 (Digitalisat).
- G. J. Dyke & C. A. Walker: New Records of Fossil ‘Waterbirds’ from the Miocene of Kenya. In: American Museum Novitates, Nummer 3610, 2008, S. 1–10 (Digitalisat).
- L. von Ammon: Tertiäre Vogelreste von Regensburg und die jungmiocäne Vogelwelt. In: Abhandlungen des Naturwissenschaftlichen Vereins (früher Zoologisch-Mineralogischen Vereins) in Regensburg, 12. Heft, 1918, S. 1–69 (Digitalisat).
- P. Ballmann: A Fossil Bird Fauna From the Pliocene Sahabi Formation in Libya. In: N. T. Boaz, A. El-Arnauti, A. W. Gaziry, J. de Heinzelin & D. D. Boaz (Hrsg.): Neogene Paleontology and Geology of Sahabi, Libya, Alan R. Liss, New York, 1987, ISBN 0-8451-4214-3, S. 113–117 (abrufbar).
- C. J. O. Harrison & C. A. Walker: Fossil birds from the Upper Miocene of northern Pakistan. In: Tertiary Research, Band 4, Nummer 2, 1982, S. 53–69.
- J. Cheneval, L. Ginsburg, C. Mourer-Chauvire & B. Ratanasthien: The Miocene avifauna of the Li Mae Long locality, Thailand: systematics and paleoecology. In: Journal of Southeast Asian Earth Sciences, Band 6, Nummer 2, 1991, S. 117–126 (Digitalisat).
- B. Vollmerhaus (Hrsg.): Lehrbuch der Anatomie der Haustiere - Band 5: Anatomie der Vögel. 3., durchgesehene Auflage, Parey Verlag, Stuttgart, 2004, ISBN 3-8304-4153-3, S. 97 (Leseprobe).
- M. Böhme: The Miocene Climatic Optimum: evidence from ectothermic vertebrates of Central Europe. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 195, 2003, S. 389–401 (Digitalisat).
- M. Böhme, M. Winklhofer & A. Ilg: Miocene precipitation in Europe: Temporal trends and spatial gradients. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 304, 2011, S. 212–218 (Digitalisat).
- M. Böhme, A. Ilg & M. Winklhofer: Late Miocene „washhouse“ climate in Europe. In: Earth and Planetary Science Letters, Band 275, 2008, S. 393–401 (Digitalisat).