Siamotyrannus
Siamotyrannus ist eine ausgestorbene Gattung der Allosauroidea aus der Gruppe der Theropoden. Die einzige bekannte Art der bislang monotypischen Gattung ist Siamotyrannus isanensis aus der Sao-Khua-Formation (Barremium; vor ca. 130,7 bis 126,3 Millionen Jahren) von Thailand.[1]
| Siamotyrannus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Lebendrekonstruktion von Siamotyrannus am Sirindhorn Museum in Non Buri, Thailand | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unterkreide (Barremium) | ||||||||||||
| 130,7 bis 126,3 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Siamotyrannus | ||||||||||||
| Buffetaut, Suteethorn & Tong, 1996 | ||||||||||||
| Art | ||||||||||||
| ||||||||||||
Etymologie und Forschungsgeschichte
Der Gattungsname leitet sich ab vom bis 1939 im Westen gebräuchlichen Namen Thailands (Siam) und dem lateinischen Wort „tyrannus“ („Alleinherrscher“, „Gewaltherrscher“, „Despot“). Der Artzusatz „isanensis“ bezieht sich auf Isan (Thai: อีสาน), die nordöstliche Region Thailands.
1976 wurden in der Sao-Khua-Formation des Verwaltungs-Distriktes Phu Wiang in der Provinz Khon Kaen der Region Isan im Zuge einer Kampagne zur Uran-Prospektion erstmals Dinosaurier-Fossilien entdeckt.[2] Eine genauere Untersuchung des Gebietes durch ein Thailändisch-Französisches Forschungsteam im Jahr 1981 ergab eine reichhaltige Fossillagerstätte und das Fundgebiet wurde im Dezember 1991 als Nationalpark Phu Wiang unter Schutz gestellt.[3]

1993 entdeckte der thailändische Geologe Somchai Triamwichanon an der sogenannten Fundstelle Phu Wiang 9 (kurz „PW.9“ oder einfach „Site No. 9“) die Überreste eines großen Theropoden.[3] Nach der Fundbergung wurden die Fossilien durch Repliken ersetzt und die ursprüngliche Fundsituation originalgetreu wieder hergestellt. Die Fundstelle wurde überdacht und ist heute für Besucher des Nationalparks zugänglich.[2]
Die Erstbeschreibung erfolgte 1996 durch Buffetaut, Suteethorn & Tong. Die Autoren beschrieben Siamotyrannus isanensis zunächst als basalen Vertreter der Tyrannosauroidea.[4]
Ein einzelnes, isoliert gefundenes Schienbein sowie mehrere, ebenfalls isolierte, große Theropodenzähne wurden 1998 von Buffetaut & Suteethorn ebenfalls dieser Art zugeordnet. Ein stichhaltiger Beleg für die Zugehörigkeit zu Siamotyrannus isanensis konnte jedoch nicht erbracht werden.[5]
2001 vermerkt Holtz ebenfalls mehrere Synapomorphien mit den Tyrannosauridae, verweist jedoch gleichzeitig auch auf das Fehlen des, diagnostisch bedeutsamen, Schädels und sieht Siamotyrannus als möglichen Vertreter einer Vorläuferlinie, außerhalb der eigentlichen Tyrannosauridae.[6]
2003 widerspricht Rauhut dieser Befundung, findet keine Synapomorphien mit den Tyrannosauridae und wertet Siamotyrannus in seiner phylogenetischen Analyse der basalen Theropoden als eigenständige „operative taxonomische Einheit“ („Operational Taxonomic Unit“; OTU).[7] Dieser Einschätzung folgen 2004 schließlich auch Holtz et al.[8]
2012 stellen Carrano et al. Siamotyrannus gemeinsam mit Metriacanthosaurus (Typusgattung) und Sinraptor in die Gruppe der Metriacanthosaurinae.[9]
Fossilbeleg
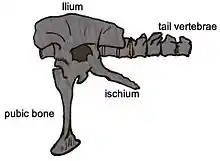
Nachdem das isolierte Schienbein und die Theropodenzähne aus der Sao-Khua-Formation nicht eindeutig zuordenbar sind, ist von Siamotyrannus isanensis dementsprechend bislang nur der Holotypus (PW.9-1) von Phu Wiang 9 gesichert bekannt. Dieser besteht aus einem artikulierten Teilskelett mit der linken Hälfte des Beckens, dem Kreuzbein, fünf Rückenwirbeln und 13 zum Rumpf hin gelegenen Schwanzwirbeln mit einigen Chevronknochen.[4]
Merkmale
Die gefundenen Knochen lassen auf einen Theropoden mit einer Länge von etwa 6,5 Metern schließen.[8]
Als hauptsächliches gattungs- und artbestimmendes, anatomisches Merkmal gelten zwei parallel verlaufende, vertikale Knochenrücken an der Außenseite des Iliums oberhalb der Hüftgelenksöffnung (Acetabulum). Diesem Merkmal kommt auch eine zentrale Rolle bei der systematischen Einordnung von Siamotyrannus zu.[5][7][9]
Systematik
Siamotyrannus wurde zunächst als primitiver Tyrannosauroide beschrieben. Ein wesentliches Argument für die Zuordnung war der oben beschriebene doppelte Knochenkamm an der Außenseite des Iliums. Ein ähnliches Merkmal weisen zwar auch die Vertreter der Tyrannosauroidea auf, bei diesen ist allerdings nur ein einfacher, dafür aber kräftiger ausgebildeter Kamm vorhanden. Das Merkmal ist zudem nicht nur auf die Tyrannosauroidea beschränkt, sondern insbesondere bei den basalen Vertretern der Tetanurae weit verbreitet.[7][9]
Andere Eigenschaften von Siamotyrannus, wie etwa ein spornartiger Fortsatz anterior zum eigentlichen Dornfortsatz der Schwanzwirbel, treten bei den Tyrannosauroidea überhaupt nicht auf, dafür aber zum Beispiel bei Allosaurus oder Sinraptor und einigen anderen Theropoden. Rauhut ordnete 2003 Siamotyrannus deshalb als basalen Carnosaurier ein.[7] Dieser Ansicht folgten 2004 im Wesentlichen auch Holtz et al. die in Fukuiraptor den nächsten bekannten Verwandten von Siamotyrannus sahen und beide gemeinsam in eine Klade stellten.[8]
2012 ergab eine umfassende phylogenetische Analyse der Tetanurae durch Carrano et al. einige in Bezug auf die systematische Stellung von Siamotyrannus wesentliche Ergebnisse:[9]
- Die Carnosauria werden von den Autoren als paraphyletische Gruppe gewertet und es wird eine Dreiteilung mit den monophyletischen Kladen Megalosauroidea, Allosauroidea und Coelurosauria vorgeschlagen. Allosauroidea und Coelurosauria lassen sich dabei weiter zusammenfassen zu den Avetheropoda, die als Schwestergruppe zu den Megalosauroidea innerhalb der Tetanurae gewertet werden.
- Das Taxon der Sinraptoridae Currie & Zhao, 1994[10] wird als Juniorsynonym der Metriacanthosauridae Paul, 1988[11] erkannt und gemäß der Prioritätsregel verworfen.
- Siamotyrannus wird gemeinsam mit Metriacanthosaurus und Sinraptor in die Gruppe der Metriacanthosaurinae in die Klade der Metriacanthosauridae innerhalb der Allosauroidea gestellt.
Siamotyrannus ist damit nicht einer der ältesten Vertreter der Tyrannosauroidea, sondern vielmehr einer der jüngsten Vertreter der Metriacanthosauridae.[12]
Systematische Stellung von Siamotyrannus innerhalb der Allosauroidea (nach Carrano et al., 2012[9]):
| Allosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
*) Juniorsynonym: Sinraptoridae
Allerdings ist auch diese systematische Zuordnung von Siamotyrannus nicht ganz unumstritten und einige Autoren halten es für möglich, dass es sich bei dieser Gattung eventuell auch um einen Vertreter der basalen Coelurosauria handeln könnte.[13]
Palökologie
Trotz des nur bruchstückhaften Fossilmaterials kann für Siamotyrannus von einer rein carnivoren Ernährungsweise ausgegangen werden. Siamotyrannus lebte annähernd zeitgleich in derselben Region mit dem Sauropoden Phuwiangosaurus, dem zumindest teilweise aquatisch lebenden Megalosauroiden Siamosaurus und dem Ornithomimosaurier Kinnareemimus.[14] Fossile Reste von Süßwasser-Haien, Knochenfischen, Schildkröten, Krokodilen und eines nicht näher bestimmten Vogels vervollständigen das Bild der Fauna der Sao-Khua-Formation.[15]
Die Sedimente der Sao-Khua-Formation wurden in einem niedrig-energetischen fluviatilen System mit mäandrierenden Flussläufen und ausgedehnten Schwemmebenen abgelagert. Als Liefergebiet der Sedimente wird das Qin-Ling-Gebirge (im heutigen Zentralchina) vermutet, das erst später im Zuge der Kollision des Indischen Subkontinents mit Eurasien räumlich abgetrennt wurde. Stabile Sauerstoffisotope aus Wirbeltierzähnen und Fischschuppen deuten auf ein wechselfeuchtes Tropenklima mit saisonal starken Niederschlägen bis zu mehreren tausend Millimetern pro Jahr hin.[16]
Pollenanalysen aus der Sao-Khua-Formation deuten auf einen Bewuchs aus häufigen Koniferen aus der Gruppe der Cheirolepidiaceae, Farnen aus der Ordnung der Schizaeales und anderen Vertretern der Echten Farne sowie untergeordnet auch Laubmoosen, Baumfarnen aus den Familien der Cyatheaceae oder der Dicksoniaceae, Königsfarngewächsen, Vertretern der Bärlappartigen und der Bennettitales, sowie Araukariengewächsen hin.[17]
Literatur
- David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Ausgabe. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2.
- Éric Buffetaut, Gilles Cuny, Jean Le Loeuff & Varavudh Suteethorn (Hrsg.): Late Palaeozoic and Mesozoic Continental Ecosystems in SE Asia, The Geological Society, London, Special Publications, No. 315, 306 S., 2009, ISBN 978-1-86239-275-5.
Einzelnachweise
- S. Tumpeesawan, Y. Sato, & S. Nakhapadungrat: A new species of Pseudohyria (Matsumotoina) (Bivalvia: Trigonioidoidea) from the Early Cretaceous Sao Khua Formation, Khorat Group, northeastern Thailand. In: Tropical Natural History, Vol. 10, S. 93–106, 2010. (Digitalisat)
- N. Boonchain, P. J. Grote & P. Jintasakul: Paleontological parks and museums and prominent fossil sites in Thailand and their importance in the conservation of fossils. In: J. H. Lipps & B. R. C. Granier (Hrsg.): PaleoParks – The protection and conservation of fossil sites worldwide. Carnets de Géologie / Notebooks on Geology, Kapitel 7, S. 75–95, Brest, 2009. (CG2009_BOOK_03/07) (Digitalisat)
- W. Songtham & B. Sektheera: Phuwiangosaurus sirindhornae. 106 S., Department of Mineral Resources, Ministry of Natural Resources and Environment, Bangkok, 2006. ISBN 978-974-226-410-9 (Digitalisat)
- E. Buffetaut, V. Suteethorn & H. Tong: The earliest known tyrannosaur from the Lower Cretaceous of Thailand. In: Nature, Vol. 381, S. 689–691, 1996. doi:10.1038/381689a0
- E. Buffetaut & V. Suteethorn: Early Cretaceous dinosaurs from Thailand and their bearing on the early evolution and biogeographical history of some groups of Cretaceous dinosaurs. In: S. G. Lucas, J. I. Kirkland & J. W. Estep (Hrsg.): Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History Bulletins, No. 14, S. 205–210, 1998. (Digitalisat)
- Th. Holtz: The Phylogeny and Taxonomy of the Tyrannosauridae. In: D. H. Tanke & K. Carpenter (Hrsg.): Mesozoic vertebrate life: new research inspired by the paleontology of Philip J. Currie. Kapitel 7, S. 64–83, Indiana University Press, Bloomington, 2001. (Digitalisat)
- O. W. M. Rauhut: The interrelationships and evolution of basal theropod dinosaurs. In: Special Papers in Palaeontology, No. 69, 213 S., The Palaeontological Association, London, 2003. (Digitalisat)
- T. R. Holtz Jr., R. E. Molnar & Ph. J. Currie: Basal Tetanurae. In: D. B. Weishampel, P. Dodson & Halszka Osmólska (Hrsg.): The Dinosauria. 2. Ausgabe, S. 71–110, University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2.
- M. T. Carrano, R. B. J. Benson & S. D. Sampson: The phylogeny of Tetanurae (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology, Vol. 10, No. 2, S. 211–300, 2012. (Digitalisat)
- P. J. Currie & X.–J. Zhao: A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. In: Canadian Journal of Earth Sciences, Vol. 30, No. 10, (1993), S. 2037–2081, 1994. (Abstract)
- G. S. Paul: Predatory Dinosaurs of the World. 464 S., Simon & Schuster, New York, 1988. ISBN 0671619462, ISBN 9780671619466
- Ch. Hendrickx, S. A. Hartman & O. Mateus: An Overview of Non-Avian Theropod Discoveries and Classification. In: PalArch’s Journal of Vertebrate Palaeontology, Vol. 12, No. 1, S. 1–73. ISSN 1567-2158, 2015. (pdf)
- A. Samathi, P. Chanthasit & P. M. Sander: A review of theropod dinosaurs from the Late Jurassic to mid-Cretaceous of Southeast Asia. In: Annales de Paléontologie, Band 105, Nummer 3, 2019, doi:10.1016/j.annpal.2019.03.003, S. 201–215, pdf.
- E. Buffetaut, V. Suteethorn & H. Tong: An early ‘ostrich dinosaur’ (Theropoda: Ornithomimosauria) from the Early Cretaceous Sao Khua Formation of NE Thailand. In: E. Buffetaut, G. Cuny, J. Le Loeuff & V. Suteethorn (Hrsg.): Late Palaeozoic and Mesozoic Continental Ecosystems in SE Asia, The Geological Society, London, Special Publications, No. 315, S. 229–243, 2009. doi:10.1144/SP315.16(pdf)
- H. Tong, J. Claude, V. Suteethorn, W. Naksri & E. Buffetaut: Turtle assemblages of the Khorat Group (Late Jurassic – Early Cretaceous) of NE Thailand and their palaeobiogeographical significance. In: E. Buffetaut, G. Cuny, J. Le Loeuff & V. Suteethorn (Hrsg.): Late Palaeozoic and Mesozoic Continental Ecosystems in SE Asia, The Geological Society, London, Special Publications, No. 315, S. 141–152, 2009. doi:10.1144/SP315.11 (pdf)
- R. Amiot, E. Buffetaut, Ch. Lécuyer, V. Fernandez, F. Fourel, F. Martineau & V. Suteethorn: Oxygen isotope composition of continental vertebrate apatites from Mesozoic formations of Thailand; environmental and ecological significance. In: E. Buffetaut, G. Cuny, J. Le Loeuff & V. Suteethorn (Hrsg.): Late Palaeozoic and Mesozoic Continental Ecosystems in SE Asia, The Geological Society, London, Special Publications, No. 315, S. 271–283, 2009. doi:10.1144/SP315.19(pdf)
- A. Racey & J. G. S. Goodall: Palynology and stratigraphy of the Mesozoic Khorat Group red bed sequences from Thailand. In: E. Buffetaut, G. Cuny, J. Le Loeuff & V. Suteethorn (Hrsg.): Late Palaeozoic and Mesozoic Continental Ecosystems in SE Asia, The Geological Society, London, Special Publications, No. 315, S. 69–83, 2009. doi:10.1144/SP315.6 (pdf)