Tanystropheus
Tanystropheus (Syn.: Tribelesodon, trivial auch „Giraffenhalssaurier“[1]) ist eine stammesgeschichtlich „primitive“ aber morphologisch sehr eigentümliche Gattung der Archosauromorpha aus der mittleren und oberen Trias Eurasiens. Typisch ist ihr stark verlängerter Hals bei einem insgesamt recht grazilen Körperbau. Mit seinen extremen Proportionen gilt Tanystropheus als biomechanisches und palökologisches Rätsel.[2]
| Tanystropheus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion eines großen Exemplars von Tanystropheus im Paläontologischen Museum Zürich. Durch die Perspektive wirkt der Hals im Verhältnis zum Rest des Körpers noch deutlich länger als er ohnehin ist. | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittel- bis Obertrias | ||||||||||||
| 247,2 bis 208,5 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tanystropheus | ||||||||||||
| von Meyer, 1852 | ||||||||||||
Etymologie
Der Name der Gattung wurde von Hermann von Meyer im Jahre 1852 geprägt.[3] Er ist altgriechisch und mittels des männlichen Suffixes -εύς -eus vom Kompositum aus dem Präfix τανύ- tany- ‚lang(gestreckt)‘ und dem Nomen στρόφος strophos ‚Strang, Band, Schnur, Gürtel‘ abgeleitet. Er bedeutet folglich soviel wie ‚der Langbändige‘ oder ‚der Langsträngige‘, was sich wahrscheinlich auf die außergewöhnliche Länge der Wirbel und den von diesen gebildeten Abschnitt der Wirbelsäule bezieht.[4] Von Meyer wusste seinerzeit allerdings noch nicht, dass es sich bei dem ‚langen Strang‘ um den Hals handelte (siehe Funde und taxonomische Geschichte).
Merkmale


Kennzeichnendstes Merkmal von Tanystropheus ist der stark verlängerte Hals, der bei den Individuen von T. hydroides mindestens genauso lang war wie Rumpf und Schwanz zusammen. Er enthielt jedoch nur 12[2][4] oder 13[5][6] amphicoele Wirbel, von denen die meisten allerdings ausgesprochen lang sind. Vermutlich aufgrund ihrer Länge, weil sie hohl sind und wegen der flügelartig abstehenden Zygapophysen an ihren Enden, die in rücken- (dorsaler) und bauchseitiger (ventraler) Ansicht entfernt an die Epiphysen von Langknochen erinnern, sind die ersten gefundenen Halswirbel seinerzeit für Extremitätenknochen gehalten worden.[3] Tatsächlich sind die längsten Halswirbel bei Tanystropheus ungefähr genauso lang wie die schwach s-förmig gekrümmten Oberschenkelknochen.[5] Bei weitgehend vollständig erhaltenen Exemplaren gelenken die hinteren Zygapophysen (Postzygapophysen) von „oben“ (dorsal) mit den vorderen Zygapophysen (Präzygapophysen) des nachfolgenden Wirbels. Die Ebene dieser Gelenkung ist relativ stark gegen die Horizontale geneigt. Die Neuralbögen sind weitgehend reduziert und der Rückenmarkskanal verläuft innerhalb der Wirbelkörper (Centra).[2] Nur an den vorderen und hinteren Enden der Wirbel befinden sich rückenseitig niedrige kleine Dornfortsätze, was nahelegt, dass die Nackenmuskulatur nicht besonders kräftig und auf die Intervertebralmuskeln beschränkt war.[2] Jeder Halswirbel gelenkte an seinem vorderen Ende bauchseitig (ventral) mit einem Paar langer, einköpfiger (monocotyler), stäbchenartiger, parallel zur Halswirbelsäule ausgerichteter Rippen. Die Rumpf- und Schwanzwirbel sind „normal“ ausgebildet und nicht verlängert.
Aufgrund der geringen Anzahl von Wirbeln und der versteifend wirkenden langen Rippen war der Hals vermutlich nicht allzu beweglich. Die relativ steil stehenden Gelenkflächen der Zygapophysen dürften dafür gesorgt haben, dass er in der vertikalen Ebene flexibler war als in der horizontalen Ebene. Falls Schädel und Halsskelett leicht genug waren, hätte Tanystropheus den Kopf trotz der schwachen Nackenmuskeln ein ganzes Stück über den Rumpf heben können.[2]
Tanystropheus erreichte eine Gesamtkörperlänge von drei bis mindestens fünf Metern. Der „katapsid“ gefensterte Schädel (unterer Temporalbogen reduziert) ist im Verhältnis zum Rest des Skelettes sehr klein. Die Kieferknochen sind mit zahlreichen spitzkonischen (das Maxillare bei T. longobardicus mit dreispitzigen[6]) Zähnen bestückt, die bei geschlossenem Maul ineinander griffen.[4] Je nach Art sind ein oder mehrere Knochen des Gaumendaches mit kleinen spitzen Zähnen (Vomer, Palatinum) oder Dentikeln (Pterygoid) besetzt. Die Vordergliedmaßen sind vergleichsweise kurz und im Bereich der Handwurzel „minimal“[2] verknöchert. Die hinteren Extremitäten sind ca. 1,7 mal so lang wie die vorderen und haben eine gut verknöcherte Fußwurzel mit normal ausgebildetem Astragalus-Calcaneus-Komplex.[2][4]
Lebensweise

Bezüglich der Lebensweise von Tanystropheus und der Funktion des langen, nicht sehr biegsamen Halses herrscht bis heute Uneinigkeit. Als relativ sicher gilt, dass Tanystropheus eine enge Beziehung zum Meer hatte, denn seine fossilen Überreste wurden ausschließlich in marinen Sedimentgesteinen gefunden. Auf eine solche Beziehung deuten auch die Hartteile von wahrscheinlichen Beutetieren hin, die in der vermuteten Magengegend von Tanystropheus-Skeletten gefunden wurden, darunter Fischschuppen und Fangarmhaken (Onychiten) von Belemniten. Wie stark die Bindung an das Wasser als Lebensraum war, ist jedoch umstritten und hängt unmittelbar von der Interpretation des Knochenbaus ab. Bearbeiter, die den Hals von Tanystropheus als extrem starr und den Rest des Körpers als relativ gut angepasst an eine schwimmende Fortbewegung betrachten, favorisieren eine aquatische Lebensweise, wobei der Vortrieb im Wasser vor allem durch Schlängelbewegungen (laterale Undulation) von Rumpf und Schwanz sowie, untergeordnet, durch Paddeln mit den Hinterbeinen erzeugt worden sein soll.[7][4] Andere Autoren vermuten eine moderate Flexibilität des Halses und sehen faktisch keine Anpassung der Extremitäten oder des Schwanzskelettes an eine schwimmende Fortbewegung. Indirekte Fossilbelege[2] für die Präsenz einer großen, schweren Muskelmasse am Schwanzansatz (siehe Funde und taxonomische Geschichte) böten eine Lösung für das biomechanische Problem der Kopflast, das an Land bei einem Tier mit einem so extrem langem Hals bei erhobenem Kopf zwangsläufig auftritt. Tanystropheus könnte demnach ein Lauerjäger gewesen sein, der z. B. auf einem Felsen an der Küste, trockenen Fußes oder in sehr flachem Wasser mit erhobenem Kopf nach vorbeischwimmenden Beutetieren Ausschau hielt.[2]
Traditionell als Jungtiere der Art T. longobardicus interpretierte kleine Individuen hatten im Verhältnis zum Rest des Körpers einen wesentlich kürzeren Hals[7] und dreispitzige Zähne (namensgebend für das Synonym Tribelesodon) auf dem Maxillare und im hinteren Teil des Dentale. Da Fischfresser üblicherweise spitzkonische Zähne haben, wurde vermutet, dass diese Individuen eine weniger meeresbezogene Lebensweise hatten und sich von Insekten ernährten. Eine neuere alternative Hypothese sieht die dreispitzigen Zähne als Analogon zu den mehrspitzigen Zähnen einiger Hundsrobben-Arten und postuliert deshalb auch für diese kleinen Vertreter eine aquatische Lebensweise. Aufgrund der Unterscheide im Gebiss und auch im Bau diverser Knochen sowie anhand von Ergebnissen morphometrischer Untersuchungen wurde in Erwägung gezogen, dass die kleinen Individuen eine andere Spezies repräsentieren könnten als die großen aus der alpinen Obertrias bekannten Individuen,[4][7] was sich taxonomisch schließlich in der Erstbeschreibung von T. hydroides für die großen Exemplare niederschlug.[6]
Funde und taxonomische Geschichte
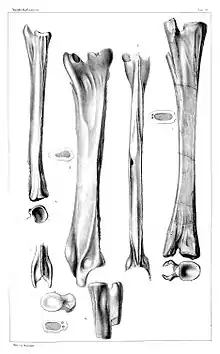



In den 1830er Jahren fand Georg Graf zu Münster bei seinen privaten Grabungen im Oberen Muschelkalk von Bayreuth hohle, stabförmige Knochen, die eine Länge von knapp 30 Zentimetern erreichten. Diese wurden durch Hermann von Meyer als Schwanzwirbel von „Macrotrachelen“, Reptilien aus der näheren Verwandtschaft von Nothosaurus, identifiziert und 1852* unter dem Namen Tanystropheus conspicuus beschrieben.[3] Im Jahre 1896 erwähnte Eberhard Fraas isolierte Zähne und Knochen aus dem Muschelkalk-Keuper-Grenzbonebed von Crailsheim, die er unter dem Namen „Nothosaurus blezingeri “ beschrieb.[9] Olivier Rieppel (1996)[10] zufolge gehören aber zumindest die in diesem Material enthaltenen Zähne zu Tanystropheus. Um die Mitte des ersten Jahrzehntes des 20. Jahrhunderts beschrieb Friedrich von Huene längliche Wirbelknochen aus dem süddeutschen, lothringischen und oberschlesischen Muschelkalk, auf die er die neuen Arten Thecodontosaurus latespinatus und Tanystrophaeus (sic!) antiquus fußte.[11][12] Auch von Huene interpretierte diese Fossilien als Schwanzwirbel, ordnete sie aber frühen Theropoden aus der engeren Verwandtschaft von Coelophysis zu.[11][12] Während „Thecodontosaurus latespinatus “ später mit Tanystropheus conspicuus synonymisiert wurde, ist Tanystropheus antiquus aufgrund seines relativ stark abweichenden Baus der Halswirbelsäule (weniger und massigere Wirbel)** und seines höheren geologischen Alters (Unterer statt Oberer Muschelkalk) in eine eigene Gattung, Protanystropheus, gestellt worden.[13] Von Huene war nicht der einzige, der Tanystropheus und frühe Theropoden „vermischte“. In einer Arbeit aus dem Jahr 1887 hatte der bekannte US-amerikanische Paläontologe Edward D. Cope Material unter den Bezeichnungen „Tanystrophaeus willistoni “, „Tanystrophaeus “ bauri und „Tanystrophaeus longicollis “ beschrieben,[14] das heute samt und sonders unter dem Namen Coelophysis bauri geführt wird.[15]
Bei und noch lange nach der Errichtung der Gattung durch von Meyer war der tatsächliche Körperbau dieser Tiere also unbekannt und ihre fossilen Überreste wurden mehrfach fehlgedeutet oder gar nicht erst erkannt. Erst im September 1929 konnte eine Arbeitsgruppe um den Zürcher Paläontologen Bernhard Peyer in den Bitumenschiefern („scisti bituminosi“, „Grenzbitumenzone“) des Monte San Giorgio im Tessin nach einer Sprengung in einem Tagebau erstmals ein fast vollständiges Skelett bergen.[16] Es handelte sich bei diesem Fund um ein Exemplar der 1886 vom Italiener Francesco Bassani anhand eines schlecht erhaltenen Skelettes aus der Umgebung von Besano, unweit des Monte San Giorgio, beschriebenen und als Flugsaurier gedeuteten Art Tribelesodon longobardicus.[17] Deren Zugehörigkeit zur Gattung Tanystropheus wurde somit klar.[16] Auch konnten zum einen die verlängerten Wirbel nunmehr korrekt als Halswirbel*** erkannt werden und zum anderen wurde offensichtlich, dass Tanystropheus kein allzu enger Verwandter von Nothosaurus und erst recht kein Dinosaurier war, sondern einer anderen Gruppe mesozoischer Reptilien angehörte, die Peyer „Tanysitrachelia“ nannte[4] und den Sauropterygia unterordnete.
Im Jahr 1975 beschrieb der hungaro-rumänische Paläontologe Tibor Jurcsák die Art Tanystropheus bihoricus anhand von Halswirbeln aus mitteltriassischen Kalksteinen der Region Bihor in den Karpaten Nordwestrumäniens. Diese Art wurde 1980 von Rupert Wild erstmals in Synonymie mit Tanystropheus longobardicus gestellt.[4] Im gleichen Jahr beschrieb Wild, der bereits in den frühen 1970ern T. longobardicus neu bearbeitet hatte, anhand relativ unvollständiger Exemplare zwei neue Tanystropheus-Arten, T. meridensis und T. fossai, aus dem Meride-Kalkstein des Monte San Giorgio bzw. dem Riva-di-Solto-Tonstein (Argillite di Riva di Solto) des Friaul (Nordost-Italien). Aber die Gültigkeit auch dieser beiden Arten ist nachfolgend in Zweifel gezogen worden. So wurde T. meridensis als Synonym von T. longobardicus eingestuft[4][18] und von T. fossai wird sogar bezweifelt, dass es sich um einen Vertreter der Gattung Tanystropheus handelt.[2][19] Wild war es auch, der den etablierten Gattungsnamen Tanystropheus von der International Commission on Zoological Nomenclature (ICZN) zum Nomen protectum erklären ließ, nachdem der angeblich durch Münster geprägte Name Macroscelosaurus von Oskar Kuhn in mehreren Arbeiten als älteres und damit prioritäres Synonym für Tanystropheus verwendet worden war.[20][21] In Einheit mit dem Schutz des Namens Tanystropheus wurde der Name Macroscelosaurus von der ICZN zum Nomen suppressum erklärt.[20][21] Des Weiteren verwarf Wild Peyers Tanysitrachelia und ordnete Tanystropheus bei den Prolacertiformes ein, die seinerzeit als „Stammgruppe“ der Lacertilia galten.
Die ersten Überreste von Tanystropheus außerhalb Europas kamen bereits in den 1950ern bei Aufsammlungen im „Muschelkalk“ des Machtesch Ramon in der Negev-Wüste in Israel zum Vorschein. Entdecker war der österreichisch-israelische Zoologe Georg Haas, der die Funde seinerzeit bei Peyer meldete und sie teilweise auch nach Zürich weitergab.[22] Auf Grundlage von mehreren unvollständigen Halswirbeln, die bei späteren Exkursionen dort gefunden wurden, beschrieb Olivier Rieppel 2001 die neue Art Tanystropheus haasi.[23] Im Jahr 2005 veröffentlichte der italienische Paläontologe Silvio Renesto die Beschreibung eines als Tanystropheus cf. longobardicus bestimmten Neufundes eines Skelettes aus dem Meride-Kalkstein des Monte San Giorgio, in dessen „Lenden“-Region deutlich sichtbar Abdrücke der Haut mit viereckigen Schuppen erhalten sind. Zudem liefern carbonatisch-phosphatische Knollen in der proximalen Schwanzregion dieses Exemplars indirekte Hinweise darauf, dass sich dort zu Lebzeiten und noch zu Beginn der Zersetzung des Kadavers eine ungewöhnlich große Muskelmasse befunden haben könnte.[2] Zwei Jahre später erschien eine Monographie der Italienerin Stefania Nosotti, in der sie Neufunde von u. a. weitgehend vollständigen Skeletten zweier Individuen von Tanystropheus longobardicus aus der Umgebung von Besano detailliert beschreibt.[4]
Im gleichen Jahr wurde der zweite Nachweis von Tanystropheus außerhalb Europas vermeldet. Es handelt sich um ein unvollständiges aber im anatomischen Zusammenhang überliefertes Skelett eines juvenilen Individuums aus dem Mittel-/Obertrias-Grenzbereich der karbonatischen Falang-Formation der Guizhou-Provinz (S-China).[24] Ein zweites Exemplar aus der Region ist 2006 gefunden und 2010 publiziert worden. Dieses relativ große, als Tanystropheus cf. longobardicus klassifizierte Exemplar stammt aus einem etwas jüngeren Intervall der gleichen Formation und besteht aus einem Rumpfskelett einschließlich der proximalen Partien der Hals- und Schwanzwirbelsäule. Es bestätigt, dass Tanystropheus in der jüngeren Trias in seiner Verbreitung offenbar nicht nur auf den Westen Laurasiens beschränkt war.[5]
Im Jahr 2015 wird von Hans-Dieter Sues und Paul E. Olsen ein einzelner, 76 Millimeter langer, unvollständiger, aber diagnostischer vorderer oder mittlerer Halswirbel aus der ? mitteltriassischen Economy-Formation als „erster Nachweis eines langhalsigen Tanystropheiden in Nordamerika“ unter dem Namen cf. Tanystropheus sp. in einer Arbeit über die Wirbeltierfossilgesellschaften des Fundy-Beckens (Nova Scotia, Ost-Kanada) abgebildet.[25] Allerdings wird die Gattung bereits 1988 eher beiläufig von Olsen in einer Zusammenschau der Fossilfaunen der gesamten Newark-Supergruppe gelistet,[26] und im Jahr 1993 wird diesbezüglich ein „einzelnes, diagnostisches [Wirbel-]Zentrum“ in der Literatur erwähnt.[27]
Im August 2020 veröffentlichen Stephan Spiekman und Kollegen die Erstbeschreibung der Art Tanystropheus hydroides aus der Grenzbitumenzone des Monte San Giorgio. Die neue Art beinhaltet Exemplare, die vormals als ausgewachsene Individuen von T. longobardicus aufgefasst wurden. Eingehendere Untersuchungen des Materials, insbesondere anhand Computertomographie-basierter dreidimensionaler Schädelrekonstruktionen, zeigten jedoch deutliche Unterschiede zwischen den Schädelproportionen und den Gebissen großer und kleiner Exemplare (u. a. eine flachere Schnauze und sehr lange Fangzähne im vorderen Teil der Schnauze bei den erstgenannten). Vor allem aber konnten knochenhistologische Untersuchungen belegen, dass die kleinen Exemplare keine Jungtiere, sondern ausgewachsen waren. Dies rechtfertigte schließlich die Errichtung einer eigenen Spezies für die großen Exemplare.[6]
Systematik
Äußere Systematik
Tanystropheus ist die Typusgattung der Familie Tanystropheidae, die weitere, sehr ähnliche Formen umfasst und auf die Trias beschränkt ist. Als relativ eng verwandt mit Tanystropheus gilt dabei die nordamerikanische Gattung Tanytrachelos. Ein etwas entfernterer Verwandter ist die chinesische Gattung Dinocephalosaurus, die deutlich stärkere Anpassungen an eine aquatische Lebensweise zeigt als Tanystropheus.
Die Tanystropheiden wiederum gehören zu einer eigenartigen permo-triassischen Linie vom Stammgruppen-Archosauriern (basalen Archosauromorpha), die unter den Namen Protorosauria oder Prolacertilia bekannt ist. Als Angehörige der Vogel-Krokodil-Linie stehen die langhalsigen Tanystropheiden somit, anders als im 20. Jahrhundert von vielen Paläontologen vermutet, weder in engerer Beziehung zu den langhalsigen mesozoischen Meeresreptilien, die in der Gruppe Sauropterygia vereint sind und als Angehörige der Schlangen-„Echsen“-Linie (Lepidosauromorpha) gelten, noch in enger Beziehung zu den „Echsen“ (Lacertilia) selbst.
Innere Systematik
Bis heute sind eine ganze Reihe von Tanystropheus-Arten beschrieben oder in der Literatur erwähnt worden (siehe Funde und taxonomische Geschichte), von denen neben der Typus-Art jedoch nur drei aktuell (Stand 2020) weitgehend anerkannt bzw. tatsächlich dieser Gattung zuzuordnen sind:[4][6]
- Tanystropheus conspicuus von Meyer, 1852 – Mitteltrias – Süddeutschland, Lothringen (Frankreich); Typus-Art
- Tanystropheus longobardicus (Bassani, 1886) – Obertrias – Tessin (Schweiz), Varese (NW-Italien), Bihor (NW-Rumänien) [Guizhou (S-China)]
- Tanystropheus hydroides Spiekmann et al., 2020 (vormals der große Morphotyp von T. longobardicus) – Obertrias – Tessin (Schweiz), Varese (NW-Italien)
- Tanystropheus haasi Rieppel, 2001 – Mitteltrias – Israel
T. longobardicus und T. hydroides sind die bekannteste Arten. Von ihnen liegt mit Abstand das meiste und vollständigste Material vor, und ein Großteil dessen, was über die Gattung bekannt ist, entstammt dem Studium von Exemplaren dieser beiden Spezies.
Weblinks
- 150 Year-Old Mystery Solved: Bizarre Giraffe-Necked Reptile Was a Surprisingly Adaptable Hunter – Bericht auf SciTechDaily vom 25. September 2020 im Zusammenhang mit der Veröffentlichung der Arbeit von Spiekman et al. (2020)
Einzelnachweise
- Rupert Wild: Der Giraffenhals-Saurier. In: Die Naturwissenschaften. Bd. 62, Nr. 4, 1975, S. 149–153, doi:10.1007/BF00608696.
- Silvio Renesto: A new specimen of Tanystropheus (Reptilia Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus. In: Rivista Italiana di Paleontologia e Stratigrafia. Bd. 111, Nr. 3, 2005, S. 377–394, (PDF 2,29 MB); siehe auch darin zitierte Literatur.
- Hermann von Meyer: Zur Fauna der Vorwelt. Die Saurier des Muschelkalkes mit Rücksicht auf die Saurier aus buntem Sandstein und Keuper. Verlag von Heinrich Keller, Frankfurt am Main 1847–1855, doi:10.3931/e-rara-43030, S. 41 f.
- Stefania Nosotti: Tanystropheus longobardicus (Reptilia, Protorosauria): re-interpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, northern Italy). In: Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano. Bd. 35, Nr. 3, 2007 (Volltext auf ResearchGate); siehe auch darin zitierte Literatur.
- Olivier Rieppel, Da-Yong Jiang, Nicholas C. Fraser, Wei-Cheng Hao, Ryosuke Motani, Yuan-Lin Sun, Zuo-Yu Sun: Tanystropheus cf. T. longobardicus from the early Late Triassic of Guizhou Province, southwestern China. In: Journal of Vertebrate Paleontology. Bd. 30, Nr. 4, S. 1082–1089, doi: 10.1080/02724634.2010.483548
- Stephan N. F. Spiekman, James M. Neenan, Nicholas C. Fraser, Vincent Fernandez, Olivier Rieppel, Stefania Nosotti, Torsten M. Scheyer: Aquatic habits and niche partitioning in the extraordinarily long-necked Triassic reptile Tanystropheus. In: Current Biology. Bd. 30, Nr. 19, 2020, S. 3889–3895, doi:10.1016/j.cub.2020.07.025
- Karl Tschanz: Allometry and heterochrony in the growth of the neck of Triassic prolacertiform reptiles. In: Palaeontology. Bd. 31, Nr. 4, 1988, S. 997–1011 (PalAss).
- Franz Nopcsa: Neubeschreibung des Trias-Pterosauriers Tribelesodon. In: Palaeontologische Zeitschrift. Bd. 5, Nr. 3, 1923, S. 161–181, doi:10.1007/BF03160365.
- Eberhard Fraas: Die schwäbischen Trias-Saurier. Schweizerbart, Stuttgart 1896, urn:nbn:de:bsz:21-dt-46495, S. 11.
- Olivier Rieppel: A revision of the genus Nothosaurus (Reptilia: Sauropterygia) from the Germanic Triassic, with comments on the status of Conchiosaurus clavatus. In: Fieldiana Geology, new series. Nr. 34, 1996 doi:10.5962/bhl.title.2691, S. 61.
- Friedrich von Huene: Die Trias-Dinosaurier Europas. In: Zeitschrift der Deutschen Geologischen Gesellschaft. Bd. 57, 1905, S. 345–349 (BHL), S. 349.
- Friedrich von Huene: Die Dinosaurier der europäischen Triasformation, mit Berücksichtigung der außereuropäischen Vorkommnisse. In: Geologische und Palæontologische Abhandlungen. Suppl.-Bd. 1, 1907–1908 (archive.org).
- Andrey G. Sennikov: New tanystropheids (Reptilia: Archosauromorpha) from the Triassic of Europe. In: Paleontological Journal. Bd. 45, Nr. 1, 2011, S. 90–104, doi:10.1134/S0031030111010151 (alternativer Volltextzugriff: ResearchGate).
- Edward D. Cope: A Contribution to the History of the Vertebrata of the Trias of North America. In: Proceedings of the American Philosophical Society. Bd. 24, Nr, 126, 1887, (BHL), S. 221 ff.
- Matthew T. Carrano, Roger B. J. Benson, Scott D. Sampson: The phylogeny of Tetanurae (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology. Bd. 10, Nr. 2, 2012, S. 211–300, doi:10.1080/14772019.2011.630927 (alternativer Volltextzugriff: ResearchGate), S. 227.
- Emil Kuhn-Schnyder: Die Triasfauna der Tessiner Kalkalpen. Neujahrsblatt 1974, Naturforschende Gesellschaft in Zürich, 1973 (PDF 25 MB), S. 50–57.
- Francesco Bassani: Sui fossili e sull’età degli schisti bituminosi triasici di Besano in Lombardia. In: Atti della Società Italiana di Scienze Naturali. Bd. 19, 1886, S. 15–72 (BHL), S. 25 ff.
- Nicholas C. Fraser, Olivier Rieppel: A new protorosaur (Diapsida) from the Upper Buntsandstein of the Black Forest, Germany. In: Journal of Vertebrate Paleontology. Bd. 26, Nr. 4, 2006, S. 866–871, doi:10.1671/0272-4634(2006)26[866:ANPDFT]2.0.CO;2, S. 866.
- Silvio Renesto: A reappraisal of the diversity and biogeographic significance of the Norian (Late Triassic) reptiles from the Calcare di Zorzino. S. 445–456 in: Jerry D. Harris, Spencer G. Lucas, Justin A. Spielmann, Martin G. Lockley, Andrew R.C. Milner, James I. Kirkland (Hrsg.): The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin. Bd. 37, 2006 (online).
- Rupert Wild: Tanystropheus H. von Meyer, [1852] (Reptilia): Revised request for conservation under the plenary powers. Z.N.(S.)2084. In: Bulletin of Zoological Nomenclature. Bd. 33, Nr. 2, 1976, S. 124–126 (BHL).
- R. V. Melville: Opinion 1186. Tanystropheus H. von Meyer, [1852] (Reptilia) conserved. In: Bulletin of Zoological Nomenclature. Bd. 38, Nr. 1, 1981, S. 188–190 (BHL).
- Bernhard Peyer: Demonstration von Trias-Vertebraten aus Palästina. In: Eclogae Geologicae Helvetiae. Bd. 48, Nr. 2, 1955, S. 486–490 (PDF 98 MB; umfasst den gesamten wissenschaftlichen Teil des Berichtes zur 34. Jahresversammlung der Schweizerischen Paläontologischen Gesellschaft).
- Olivier Rieppel: A new species of Tanystropheus (Reptilia: Protorosauria) from the Middle Triassic of Makhtesh Ramon, Israel. In: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. Bd. 221, Nr. 2, 2001, S. 271–287, doi:10.1127/njgpa/221/2001/271
- Li Chun: A juvenile Tanystropheus sp. (Protorosauria, Tanystropheidae) from the Middle Triassic of Guizhou, China. In: Vertebrata PalAsiatica. Bd. 45, Nr. 1, 2007, S. 37–42 (PDF 550 kB).
- Hans-Dieter Sues, Paul E. Olsen: Stratigraphic and temporal context and faunal diversity of Permian-Jurassic continental tetrapod assemblages from the Fundy rift basin, eastern Canada. Atlantic Geology. Bd. 51, Nr. 1, 2015, S. 139–205, doi:10.4138/atlgeol.2015.006 (Open Access), Fig. 10
- Paul E. Olsen: Paleontology and paleoecology of the Newark Supergroup (early Mesozoic, eastern North America). S. 185–230 in: Warren Manspeizer (Hrsg.): Triassic-Jurassic Rifting: Continental Breakup and the Origin of the Atlantic Ocean and Passive Margins. Developments in Geotectonics 22. Elsevier, Amsterdam/New York 1988, ISBN 0-444-42903-4, S. 189 (Fig. 8-4), 221
- Phillip Huber, Spencer G. Lucas, Adrian P. Hunt: Vertebrate biochronology of the Newark Supergroup Triassic, eastern North America. S. 179–186 in: Spencer G. Lucas, Michael Morales (Hrsg.): The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin. Bd. 3, 1993 (online), S. 180.
- Susan E. Evans: The early history and relationships of the Diapsida. S. 221–260 in: Michael J. Benton (Hrsg.): The Phylogeny and Classification of the Tetrapoda. Volume 1: Amphibians, Reptiles, Birds. Systematics Association Special Volume 35A. Clarendon Press, Oxford 1988 (Digitalisat auf ResearchGate), S. 227.