Lambeosaurinae
Die Lambeosaurinae ist ein Taxon der Hadrosaurier. Sie sind auf den Kontinenten Nordamerika und Asien aus der Oberkreide fossil nachweisbar. Das wichtigste Merkmal war ein Kamm, der aus dem Zwischenkieferbein (Prämaxillare) und dem Nasenbein (Nasale) gebildet wurde.[3] Die Lambeosaurinae sind die Hadrosaurier, die näher mit Lambeosaurus als mit Hadrosaurus verwandt sind.
| Lambeosaurinae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
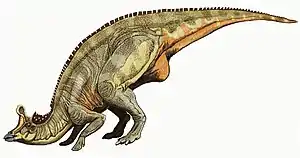
Zeichnerische Lebendrekonstruktion von Lambeosaurus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (spätes Santonium bis Maastrichtium)[1] | ||||||||||||
| 85,2 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Lambeosaurinae | ||||||||||||
| Parks, 1923 |
Merkmale
Rumpf und Größe
Die Lambeosaurinae waren robuster gebaut als die Hadrosaurinae.[4] Sie wiesen im Gegensatz zu ihren Gegenstücken hohe Dornfortsätze auf; die den Rücken und den Schwanzansatz deutlich erhöhten.[5] Bis auf Olorotitan mit 18 Halswirbeln wiesen die Lambeosaurinae wie alle anderen Hadrosaurier bis zu 15 Halswirbel auf.[6] Ebenfalls ein Merkmal ist, dass die Speiche (Radius) länger als der Oberarmknochen (Humerus) war.[3]
Die Lambeosaurinae waren im Vergleich zu den Hadrosaurinae relativ große Tiere. Die meisten Gattungen waren 9 bis 10 m lang.[6][7][8][9] Charonosaurus wird auf eine Länge von 13 m geschätzt,[10] Lambeosaurus sogar auf eine Größe von 15 bis 16 m – damit wäre Lambeosaurus der wohl größte Lambeosaurinae.[3] Zum Vergleich: Der größte Hadrosaurinae, Shantungosaurus, wurde etwa 14 m lang.[3] Der bisher älteste Vertreter, Nipponosaurus, war mit 8 m ein eher kleiner Lambeosaurinae.
Schädel

Das wichtigste Merkmal der Lambeosaurinae ist der aus Zwischenkieferbein (Prämaxillare) und Nasenbein (Nasale) gebildete Knochenkamm.[3] Der meistens nach hinten gerichtete Kamm variiert in der Form von knochenzapfenartig (z. B. Parasaurolophus)[11] bis beilförmig, patten-[12] bzw. helmartig (z. B. Corythosaurus).[8] Bei der Gattung Tsintaosaurus ist die Kammform umstritten, wird aber häufig als einhornartig gedeutet.[9] Allerdings kann der Kamm auch kaum vorhanden sein.[3] Der Größendimorphismus legt bei einer Art nahe, dass Männchen und Weibchen zwei verschiedene Kämme hatten.[3] Die Kämme wurden Röhren durchzogen, mit denen Töne erzeugt werden konnten. In den Kämmen waren 4 Röhren, zwei verliefen nach oben, zwei nach unten; die Luft musste die ganzen Röhren durchqueren.[13]
Wie bei allen alle Hadrosauriern war die Schnauze breit, allerdings schmaler als bei den Hadrosaurinae.[14] Bei Lambeosaurus hatte ein zur Schnauze weisender Auswuchs des Jochbeins (Jugale) eine dreieckige Form.[3]
Systematik und Evolution
Äußere Systematik
Die Lambeosaurinae wird der Hadrosaurinae gegenübergestellt. Der Hauptunterschied ist dabei, dass die Hadrosaurinae keinen Kamm besitzt.[3] Beide Taxa bilden wiederum das Taxon Euhadrosauria. Kladogramm laut Vecchia:[15]
| Hadrosaurier (Hadrosauridae) |
| ||||||||||||||||||||||||||||||
Evolution und Paläobiogeographie
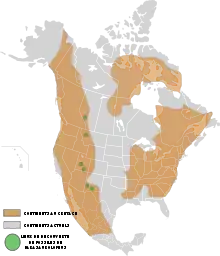

In den 1980er-Jahren löste der Hadrosaurier-ähnliche Schädel der Gattung Ouranosaurus eine Diskussion aus. Laut Philippe Taquet ähnelte der Schädel von Ouranosaurus den Hadrosauriern so sehr, dass er ein Vorfahr der Hadrosauridae sein könnte, und somit auch ein Vorfahr der Lambeosaurinae. Jack Horner meinte 1990, Ouranosaurus habe ähnliche Schädelmerkmale wie die Lambeosaurinae; somit wurden Ouranosaurus und die Lambeosaurinae als Lambeosauria zusammengefasst. Die Hadrosaurinae dagegen seien näher mit Iguanodon verwandt. Sollte diese Theorie stimmen, so bilden die Hadrosaurier keine monophyletische Klade mehr, sondern die Lambeosaurinae und die Hadrosaurinae hätten sich aus separaten Vorfahren und anderen Zeiten entwickelt. Diese Theorie ist aber nicht bewiesen.[12]
Der Ursprung der Lambeosaurinae selbst war wahrscheinlich Asien. Dort spalteten sich die Gattungen Aralosaurus und Jaxartosaurus ab. Ebenfalls wanderte ein Teil einer Population in Richtung China; dort entstand die Gattung Tsintaosaurus. Der andere Teil der Population wanderte über die Beringstraße nach Nordamerika ein. Dort spalteten sich die Parasaurolophini von den anderen Lambeosaurinae, den Corythosaurini, ab und bildeten durch Anpassung an die Natur die zapfenartigen Kämme. Da die Lambeosaurinae eher im Norden als im Süden Nordamerikas gefunden worden sind, spricht dies ebenfalls für diese Entwicklung. Die nordamerikanischen Lambeosaurinae wanderten wahrscheinlich auch wieder zurück nach Asien. Ein gutes Beispiel für diese Theorie sind Charonosaurus und Parasaurolophus: Beide weisen schnorchelartige Kämme auf. Da Charonosaurus geologisch jünger als Parasaurolophus ist, könnte Parasaurolophus nach Asien eingewandert sein. Die Lambeosaurinae starben in Nordamerika noch vor dem Ende der Kreide, in Asien dagegen starben sie mit allen anderen Nicht-Vogel-Dinosauriern am Ende der Kreidezeit vor rund 66 Mio. Jahren aus.[16]
Innere Systematik
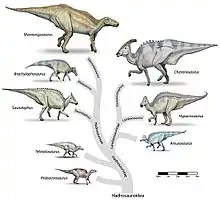
Die Lambeosaurinae lassen sich generell in zwei Tribus einteilen: In die Parasaurolophini, mit den schnorchelartigen Kämmen, und in die Corythosaurini mit den „runden“ Kämmen, zu denen auch die Gattung Lambeosaurus gehört. Zu den basalen Lambeosaurinae befinden sich unter anderem Tsintaosaurus und Olorotitan.[4]
| Lambeosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paläobiologie
Funktion der Kämme
Die Funktion(en) der Kämme der Lambeosaurier ist umstritten. Früher wurden die Hadrosaurier als amphibische Tiere gedeutet; der Kamm könnte, vor allem bei Parasaurolophus eine Art Schnorchel gewesen sein. Andere Vorschläge sind, dass der Kamm als Salzdrüse oder erweiterten Geruchssinn diente.[4] Man geht inzwischen davon aus, dass die Kämme bunt waren und in der Balz dienten.[3] Die Hohlräume waren an die Luftröhre und an die Nase angeschlossen. Damit könnten Töne erzeugt werden.[4]
Lebensraum und Paläoökologie
Früher ging man davon aus, dass die Lambeosaurinae wie andere Hadrosaurier amphibisch lebten. Richard Deckert stellte die Gattung Corythosaurus im Wasser mit horizontalen Körper dar, an Land allerdings rekonstruierte Deckert ihn auf zwei Beinen stehend. Der Erstbeschreiber Barnum Brown schloss aus dem hohen, seitlich abgeflachten Schwanz auf eine schwimmende Lebensweise. Diese Theorie hielt sich über Jahrzehnte.[12] Eine weitere Stütze dieser Theorie der amphibischen Lebensweise war der schnorchelartige Kamm des Parasaurolophus.[4]
Heute weiß man, dass die Lambeosaurinae terrestrische, also an Land lebende Tiere waren. In Nordamerika war die Landschaft in Alberta im Norden von Flüssen und sumpfigen Auen durchzogen und es gab ein vom Western Interior Seaway marin beeinflusstes Klima,[17] in New Mexico im Süden war Nordamerika durch das Zurückziehen des Western Interior Seaways eine Überflutungsebene.[18]
In Asien gab es sowohl Wüste als auch Wald. Unter anderem gab es auch einige Seen.[19]
Paläoökologie
In der Dinosaur-Park-Formation in Nordamerika, in der Lambeosaurinae gefunden wurden, wurden auch viele Ceratopsier gefunden, darunter Centrosaurus, Styracosaurus und Chasmosaurus, aber die nordamerikanischen Lambeosaurinae teilten ihren Lebensraum sowohl mit den Hadrosaurinae Prosaurolophus und Gryposaurus, den Ankylosauriden Edmontonia und Euoplocephalus, als auch den Tyrannosauroiden Gorgosaurus.[20] In der Horseshoe-Canyon-Formation, ebenfalls Nordamerika, teilten sich die Lambeosaurinae den Lebensraum mit den Hadrosaurinae Edmontosaurus regalis und Saurolophus osborni, den Ceratopsier mit den Arten Pachyrhinosaurus canadensis, Anchiceratops ornatus, Arrhinoceratops brachyops und Eotriceratops xerinsularis. Eher selten waren Ankylosaurier, darunter Euoplocephalus tutus und Edmontonia longiceps oder Pachycephalosaurier wie Stegoceras edmontonense. Ebenfalls ein Ornithischier ist der Ornithopode Parksosaurus warrenae. Die Theropoden waren mit den Ornithomimiden wie Dromiceiomimus brevitertius, Ornithomimus edmontonicus und Struthiomimus altus sehr häufig. Der Spitzenprädator war der Tyrannosauride Albertosaurus sarcophagus. Kleinere Räuber waren Ricardoestesia isosceles, der Caenagnathiden Chirostenotes pergracilis, den Troodontide Troodon formosus und die Dromaeosauriden Atrociraptor marshalli, Dromaeosaurus sp. und Saurornitholestes sp.[21][22]
In Asien teilten sich die Lambeosaurinae den Lebensraum mit den Hadrosaurine Kerberosaurus,[6] den Protoceratopsiden Protoceratops,[23] einigen Pachycephalosaurier, den Ankylosaurier Saichania[24] den Lebensraum, Theropoden waren durch den Gattungen Tarbosaurus[25] und Velociraptor vertreten.

Ernährungsweise
Alle Lambeosaurinae waren Herbivoren. Die fortgeschrittenen Kiefer erlaubten mahlende Kaubewegungen. Die Kauleisten bildeten Zahnbatterien, in denen mehrere hundert Zähne angeordnet waren; es wurden immer mehrere Zähne benutzt. Abgenutzte Zähne wurden ständig ersetzt. Die Pflanzen wurden erst mit dem Schnabel abgerupft und anschließend in wangenartigen Mundpartien gehalten.[3] Der Paläontologe Robert Bakker meinte, die Lambeosaurinae hätten schmalere Schnauzen als die Hadrosaurinae. Bakker stellte die Theorie auf, dass die Lambeosaurinen Hadrosaurier wählerischere Pflanzenfresser waren.[14]
Entdeckungs- und Forschungsgeschichte
1910 bis 1930
Der erste bekannte Lambeosaurinae war der von Barnum Brown 1913 beschriebene Hypacrosaurus. Er war ursprünglich durch Rippen, Wirbel, Schulter- und Beckengürtel und Knochen von Gliedmaßen bekannt. Brown allerdings fiel der Riesenwuchs auf und sprach diesem Hadrosaurier eine „aufgerichtete Höhe“ von fast der des Tyrannosaurus und nannte es „fast größte Echse“. Die Form des Schädels wurde 1924 durch eine Beschreibung eines zweiten Exemplars bekannt.[12] Im Jahr 1914 beschrieb Brown Corythosaurus; er wurde nach einem bemerkenswerten Fund bei Steveville beim Red Deer River, Alberta, 1912 benannt. Bis auf ein Teil der Vorderbeine und des Schwanzes vollständig erhalten. Bedeutend ist unter anderem, dass die Fossilien von Corythosaurus sehr gut erhalten sind, sie zeigen die Umrisse der Weichteile vom gesamten Tier, selbst die Hautpartien. Brown allerdings war aufmerksam auf parallel verlaufende, vertikale Falten; sie waren in der Schulter und im Rumpf gut erkennbar. Brown meinte, es seien „lose Hautfalten“, die beim lebenden Tier erkennbar sein müssen. Laut Brown habe das Corythosaurus-Fossil neben hohen Wirbelfortsetzungen vertikale Hautfurchen gezeigt. Richard Deckert rekonstruierte Corythosaurus für Browns Artikel mit einer gefalteten Hautoberfläche. Als Deckert Corythosaurus schwimmend und an Land aufrecht mit zwei Beinen stehend rekonstruierte, hielt sich die Theorie, Hadrosaurier seien amphibisch, im frühen 20. Jahrhundert.[12]
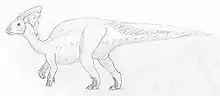
Als die University of Toronto eine Expedition nach Alberta machte, wurde, ebenfalls am Red Deer River, 1920 die Gattung Parasaurolophus entdeckt.[26] Das Exemplar ist der Holotyp mit der Exemplarnummer ROM 768. Es besteht aus einem Teilskelett mit Schädel, allerdings fehlen die Beinknochen unterhalb der Kniegelenke und der Großteil des Schwanzes. Das Tier wurde 1922 von William Parks beschrieben und nannte es Parasaurolophus walkeri. Allerdings wurde schon 1917 einige Parasaurolophus-Fossilien entdeckt, aber erst 1924 von Gilmore als Lambeosaurus sp. beschrieben.[27] Durch die Form des Kamms ging man davon aus, dass er als Schnorchel benutzt wurde, was die Theorie unterstützt, dass Hadrosaurier amphibische Tiere waren.[12] Parks glaubte, Parasaurolophus sei ein enger verwandter von Saurolophus, darum auch der Name „fast Saurolophus“. Allerdings bemerkte Parks, dass die Schnauze des Tieres der von Corythosaurus ähnelte und vermutete eine Verwandtschaft zwischen den beiden Gattungen, die sich auch bestätigte. Parks Vermutung, der Knochenzapfen sei mit einem Hautkragen mit dem Rücken verbunden, wurde allerdings nicht bestätigt, der Maler Charles Knight aber zeichnete Parasaurolophus nach Parks Vermutung.[12]
William Parks hat 1922 die neue Gattung Lambeosaurus. Die Fossilien wurden in der Judith-River-Gruppe im Dinosaur Provincial Park, Alberta. Dabei waren fast 20 relativ gut erhaltene Schädel, manchmal mit dazugehörenden Skeletten. Es wurden sowohl adulte (erwachsende) als auch Jungtiere gefunden. In der El-Gallo-Formation im mexikanischen Bundesstaat Baja California Norte wurde ein weiteres Teilskelett von Lambeosaurus freigelegt.[3]
1930 bis 1960
1931 wurde eine neue Art der Gattung Parasaurolophus entdeckt: Parasaurolophus tubicen. Die Art wurde von Carl Wiman beschrieben, allerdings schon 1921 entdeckt.[28] Der Artname tubicen kommt aus dem lateinischen und bedeutet „Trompeter“.[29]
Auf der Insel Sachalin wurde 1936 eine neue Art entdeckt, die als Nipponosaurus sachalinensis bekannt ist. Sie wurde vom japanischen Paläontologen Nagao beschrieben. Der Name bedeutet „Echse aus Japan“, da die Insel einmal zu Japan gehörte, heute wird sie von Russland regiert.[4]
Der Paläontologe Anatoli Nikolajewitsch Rjabinin beschrieb, je nach Quelle 1937[3] oder 1939,[4] die neue, wenig bekannte Art Jaxartosaurus aralensis.[3]
In den 1940er-Jahren wurden keine weiteren Lambeosaurinae entdeckt, erst 1958 beschrieb der chinesische Paläontologe Yang Zhongjian die neue Gattung Tsintaosaurus. Tsintaosaurus ist nach der Stadt Tsingtao (Qingdao) benannt. Die einzige Art ist Tsintaosaurus spinorhinus; der Artname spinorhinus bedeutet „Stachelnase“.[9] Er ist der erste bekannte Lambeosaurinae, der einen einhornartigen Kamm trägt.
1960 bis 2000
Anatoli Konstantinowitsch Roschdestwenski hatte 1968 die Art Aralosaurus tuberiferous beschrieben. Entdeckt wurden die Fossilien der Gattung in Kasachstan in der Beleutinsk-Formation nahe dem Aralsee. Er ist durch einen teilweise erhaltenen Schädel, einige Wirbel, Gliedmaßenknochen und einzelne Zähne bekannt.[3]
1993 wurde Pararhabdodon isonensis von Casanovas-Cladellas, Santafe-Llopis und Isidoro Llorens beschrieben. Er war ein möglicher verwandter des Tsintaosaurus. Ein mögliches Synonym ist Koutalisaurus.[4]
Neuere Entdeckungen
Im Jahr 2000 wurde eine neue Art beschrieben, die Parasaurolophus ähnlich sah: Charonosaurus jiayinensis. Er wurde in der Yuliangze-Formation schon in den Jahren 1916/1917 gefunden und ausgegraben. Der Holotyp ist ein fragmentarischer Schädel, der 2000 von Pascal Godefroit, Shuqin Zan und Liyong Jin als Charonosaurus beschrieben wurde.[16]
Im Jahr 2003 wurde der asiatische Olorotitan beschrieben. Er wurde 1991 in der Udurchukan-Formation (der untere Teil der Tsagayan-Group) entdeckt.[6]
Weitere Entdeckungen aus den Jahren 2006 bis 2010 waren Koutalisaurus kohlerorum, Nanningosaurus dashiensis, Velafrons coahuilensis, Sahaliyania elunchunorum, Arenysaurus ardevoli, Angulomastacator daviesi und Blasisaurus canudoi.[4]
Im November wurde die Entdeckung der ersten afrikanischen Hadrosaurierart bekannt gegeben. Ajnabia odysseus gehört zu den Lambeosaurinae und lebte während des Maastrichtiums. Kieferknochen und Zähne der Art wurden in der Nähe von Casablanca gefunden.[2]
In der Populärkultur
Obwohl die meisten Lambeosaurinae recht unbekannt sind, gibt es einige Arten, die häufig in Dokumentationen, Spielfilmen etc. vorkommen. Der berühmteste Dinosaurier dieser Unterfamilie ist Parasaurolophus. Er kommt in den Spielfilmen In einem Land vor unserer Zeit (obwohl eigentlich ein Saurolophus),[30] im Abschnitt „Le sacre du Printemps“ aus Disneys Fantasia,[31] in allen drei Jurassic-Park-Filmen[32] und Disneys Dinosaurier[33] vor. Auch als Spielzeug ist der Parasaurolophus beliebt, so wird er z. B. in der Firma Schleich hergestellt.[34] Die Gattung Corythosaurus kommt in sehr vielen populärwissenschaftlichen Büchern vor. Andere, weniger bekannte Gattungen sind unter anderem Lambeosaurus und Tsintaosaurus.
Weblinks
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, S. 306–316, ISBN 978-0-691-13720-9, Online (Memento vom 13. Juli 2015 im Internet Archive).
- Nicholas R. Longrich et al. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretaceous Research, published online November 2, 2020; doi: 10.1016/j.cretres.2020.104678
- John R. Horner, David B. Weishampel, Catherine A. Forster: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 438–463.
- Lambeosaurinae auf thescelosaurus.com (Memento vom 4. August 2009 im Internet Archive)
- Richard S. Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America (= The Geological Society of America. Special Papers. Nr. 40, ISSN 0072-1077). The Geological Society of America, New York NY 1942, S. 209–213.
- Pascal Godefroit, Yuri Bolotsky, Vladimir Alifanov: A remarkable hollow-crested hadrosaur from Russia: an Asian origin for lambeosaurines. In: Comptes Rendus Palevol. Bd. 2, Nr. 2, 2003, ISSN 1631-0683, S. 143–151, doi:10.1016/S1631-0683(03)00017-4.
- Donald F. Glut: Parasaurolophus. In: Donald F. Glut: Dinosaurs. The Encyclopedia. McFarland & Co, Jefferson NC u. a. 1997, ISBN 0-89950-917-7, S. 678–684.
- www.enchantedlearning.com - Corythosaurus (en)
- Tsintaosaurus auf Dinoruss.com (webarchive) (Memento vom 24. Mai 2008 im Internet Archive)
- Charonosaurus. In: Luis V. Rey’s Art Gallery Dinosaurs and Paleontology. Archiviert vom Original am 7. Juli 2009; abgerufen am 22. Oktober 2014 (englisch).
- Robert M. Sullivan, Thomas E. Williamson: A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus (= New Mexico Museum of Natural History and Science. Bulletin. 15, ISSN 1524-4156). New Mexico Museum of Natural History and Science, Albuquerque NM 1999, Digitalisat.
- Darren Naish: Die faszinierende Entdeckung der Dinosaurier. Konrad Theiss Verlag GmbH, Stuttgart 2010, ISBN 978-3-8062-2325-5.
- Michael J. Benton: Paläontologie der Wirbeltiere. Pfeil, München 2007, ISBN 978-3-89937-072-0, S. 226.
- Robert T. Bakker: The Dinosaur Heresies. New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. William Morrow, New York NY 1986, ISBN 0-688-04287-2, S. 194.
- Fabio M. Dalla Vecchia: Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. In: Journal of Vertebrate Paleontology. Bd. 29, Nr. 4, 2009, ISSN 0272-4634, S. 1100–1116, doi:10.1671/039.029.0428.
- Pascal Godefroit, Shuqin Zan, Liyong Jin: Charonosaurus jiayinensis n.g., n.sp.,a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China. In: Comptes Rendus de l'Académie des Sciences. Série 2, Fascicule A: Sciences de la Terre et des Planètes. Bd. 330, Nr. 12, 2000, ISSN 0764-4450, S. 875–882, doi:10.1016/S1251-8050(00)00214-7.
- David A. Eberth: The Geology. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A spectacular ancient Ecosystem revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 54–82.
- Dale A. Russell: An odyssey in time. The dinosaurs of North America. NorthWord Press u. a., Minocqua WI u. a. 1989, ISBN 1-55971-038-1, S. 160–164.
- Dinosaurier – Im Reich der Giganten: Das Rätsel der Riesenklaue. BBC, 2002.
- David B. Weishampel, Paul M. Barrett, Rodolfo Coria, Jean Le Loeuff, Xing Xu, Xijin Zhao, Ashok Sahni, Elizabeth Gomani, Christopher R. Noto: Dinosaur distribution. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–683, hier S. 517–606.
- David A. Eberth: Edmonton Group. In: Philip J. Currie (Hrsg.): Encyclopedia of Dinosaurs. Academic Press, San Diego CA u. a. 1997, ISBN 0-12-226810-5, S. 199–204.
- Edmonton Group (www.dinoruss.com / webarchive) (Memento vom 27. Juni 2008 im Internet Archive)
- You Hailu, Peter Dodson: Basal Ceratopsia. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 478–493.
- Matthew K. Vickaryous, Teresa Maryańska, David B. Weishampel: Ankylosauria. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 363–392.
- Tomasz Jerzykiewicz, Dale A. Russell: Late Mesozoic stratigraphy and vertebrates of the Gobi Basin. In: Cretaceous Research. Bd. 12, Nr. 4, 1991, ISSN 0195-6671, S. 345–377, doi:10.1016/0195-6671(91)90015-5.
- William A. Parks: Parasaurolophus Walkeri. A new genus and species of crested Trachodont dinosaur (= University of Toronto Studies. Geology Series. Bd. 13, ISSN 0372-4913). University of Toronto – University Library, Toronto 1922, Digitalisat.
- David C. Evans, Robert R. Reisz, Kevin Dupuis: A juvenile Parasaurolophus (Ornithischia: Hadrosauridae) Braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. In: Journal of Vertebrate Paleontology. Bd. 27, Nr. 3, 2007, S. 642–650, doi:10.1671/0272-4634(2007)27[642:AJPOHB]2.0.CO;2.
- Carl Wiman: Parasaurolophus tubicen n. sp. aus der Kreide in New Mexico (= Nova acta Regiae Societatis Scientiarum Upsaliensis. Serie 4, Bd. 7, Nr. 5, ZDB-ID 210356-4). Almqvist & Wiksell, Uppsala 1931.
- David P. Simpson: Cassell's Latin – English, English – Latin Dictionary. 5th edition. Cassell u. a., London u. a. 1979, S. 883.
- Die „In einem Land vor unserer Zeit“-DVD (Memento vom 17. Februar 2009 im Internet Archive)
- Fantasia (1940). In: Movie Mistakes. Abgerufen am 19. August 2009.
- Parasaurolophus. In: Park Pedia. Abgerufen am 19. August 2009.
- Disney Dinosaur Interviews David Krentz. In: The DINOSAUR Interplanetary Gazette. Archiviert vom Original am 28. Oktober 2010; abgerufen am 8. Juli 2013.
- Parasaurolophus auf Schleich.com (Memento vom 24. September 2015 im Internet Archive) (en)