Parasaurolophus
Parasaurolophus ist eine Gattung von Dinosauriern aus der Gruppe der Hadrosaurier, die zu den Ornithischia (Vogelbeckendinosaurier) gezählt wird. Funde dieser Tiere stammen aus Gesteinsschichten der Oberkreide (spätes Campanium) in Alberta (Kanada), Utah und New Mexico (USA). Wie alle Hadrosaurier konnten sich die Pflanzenfresser wahrscheinlich zweibeinig und vierbeinig fortbewegen[2] und besaßen den charakteristischen zahnlosen „Entenschnabel“ sowie Mahlzähne im Kiefer. Das auffälligste Merkmal ist der Kopfschmuck, ein langer, nach hinten gerichteter Knochenzapfen. Parasaurolophus gehört aufgrund seiner auffälligen Erscheinung zu den in der Öffentlichkeit bekannteren Vertretern der Dinosaurier.
| Parasaurolophus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Parasaurolophus cyrtocristatus im Field Museum of Natural History in Chicago. | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (spätes Campanium)[1] | ||||||||||||
| 76,4 bis 72 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Parasaurolophus | ||||||||||||
| Parks, 1922 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Beschreibung

Im Gegensatz zu einigen anderen populärwissenschaftlich oft rezipierten Hadrosauriern wurden relativ wenige Parasaurolophus-Exemplare gefunden, wodurch das Skelett nicht vollständig bekannt ist. Das Holotypusexemplar von Parasaurolophus walkeri wird auf 9,5 m Länge geschätzt, wobei der Schädel zusammen mit dem Knochenzapfen etwa 1,6 m ausmacht. Schädel und Knochenzapfen des Holotypusexemplars von Parasaurolophus tubicen messen 2 m.[3] Das Gewicht des lebenden Tieres lag bei etwa 2,5 Tonnen.[4] Das einzige bekannte Vorderbein ist verglichen mit anderen Hadrosauriden relativ kurz, wobei die Schulterblätter kurz aber breit sind. Der Oberschenkelknochen (Femur) von Parasaurolophus walkeri ist 103 cm lang und für seine Länge ziemlich robust. Auch der Schulterbereich und die Beckenknochen waren recht schwer gebaut.[5][6]
Wie andere Lambeosaurinen wies Parasaurolophus hohe Dornfortsätze auf[5], die den Rücken und den Schwanzansatz deutlich erhöhten. Ihren höchsten Punkt erreichten die Dornfortsätze über den Hüften. Von Parasaurolophus walkeri sind darüber hinaus Hautabdrücke bekannt, die gleichförmige tuberkelähnliche Schuppen, jedoch keine größeren Strukturen zeigen.[7]

Das auffälligste Merkmal der Gattung war der Knochenzapfen am Schädel. Dabei handelt es sich um eine lange Röhre, die nach hinten über den Kopf hinausragte und aus dem Nasenbein und dem Zwischenkieferbein (Prämaxillare) gebildet wurde. Der Knochenzapfen war größtenteils hohl, allerdings führten Röhren von den Nasenlöchern bis zum Ende, machten eine Kurve und führten wieder in den Schädel zurück. Während diese Röhren bei Parasaurolophus walkeri vergleichsweise einfach gebaut waren, wurde der Aufbau bei Parasaurolophus tubicen komplizierter. Bei dieser Art zeigten die Röhren labyrinthförmige Aufspaltungen, Zusammenläufe und Sackgassen.[8]
Bei dem Holotypus-Skelett von Parasaurolophus walkeri wurde eine Einkerbung in den Wirbelfortsätzen gefunden, die an der Stelle liegt, wo der Knochenzapfen den Rücken berührt hätte. Es gab Vermutungen, nachdem das Tier beim Laufen seinen Knochenzapfen in diese Kerbe gelegt hätte.[9] Wahrscheinlich ist dieses Merkmal jedoch pathologisch bedingt und eine Besonderheit dieses Individuums.[5] William Parks, der Erstbeschreiber des Tieres, postulierte Ligamente oder Muskeln, die vom Ende des Knochenzapfens bis zu den Wirbeln im Bereich der Einkerbung verlaufen sollten, um den Kopf zu unterstützen.[7] Dies erscheint jedoch unwahrscheinlich[4] – ein Überbleibsel dieser Idee ist jedoch ein Hautlappen, der in einigen Rekonstruktionen zwischen Knochenzapfen und Rücken aufgespannt ist. Nach der Vorstellung mancher Illustratoren war dieser hypothetische Hautlappen sowie der erhöhte Rücken für die Arterkennung oder für die Brautwerbung bunt gefärbt.[10]
Systematik und Arten

Innerhalb der Hadrosauridae wird Parasaurolophus zu den Lambeosaurinen gezählt, obwohl er anfangs als ein naher Verwandter des Hadrosaurinen Saurolophus galt.[11] Der nächste Verwandte von Parasaurolophus war möglicherweise Charonosaurus aus der Amur-Region Nordostchinas. Charonosaurus besitzt einen ähnlichen Schädel wie Parasaurolophus, der Knochenzapfen ist jedoch nur unvollständig überliefert. Die nahe Verwandtschaft dieser Gattungen wird an gemeinsamen Merkmalen (Synapomorphien) an den Schädel- und Beinknochen festgemacht; so bildete beispielsweise das paarige Stirnbein eine Plattform zur Unterstützung des Knochenzapfens, die sich bis über Teile des Supratemporalfensters erstreckte. Im Unterschied zu Charonosaurus zeigte Parasaurolophus aber beispielsweise einen sagittalen Kamm am Scheitelbein (Parietale).[12][13] Parasaurolophus und Charonosaurus werden innerhalb der Lambeosaurinae als Parasaurolophini zusammengefasst und der Schwestergruppe Lambeosaurini, die alle anderen Lambeosaurinen umfasst, gegenübergestellt.[14][2][15]
Bisher wurden drei Arten der Gattung Parasaurolophus beschrieben:
Von der Typusart Parasaurolophus walkeri (Parks, 1922) aus Alberta ist ein einziges Skelett mit einem kompletten Schädel bekannt.[2] Diese Art unterschied sich von Parasaurolophus tubicen durch eine einfachere Röhrenstruktur des Knochenzapfens; im Gegensatz zu Parasaurolophus cyrtocristatus wies sie einen langen, nicht nach unten gebogenen Knochenzapfen auf. Im Unterschied zu allen anderen Lambeosaurinen, abgesehen von Parasaurolophus cyrtocristatus, zeigte Parasaurolophus walkeri außerdem einen tieferen Hals des Präpubis und längere Oberarmknochen (Humerus) als Unterarme.[15]
Parasaurolophus tubicen (Wiman, 1931) aus der Kirtland-Formation in New Mexico ist durch Funde von mindestens drei Individuen bekannt.[2] Sie war die größte Art, zeigte komplexere Röhren im Knochenzapfen als Parasaurolophus walkeri und besaß einen längeren und geraderen Knochenzapfen als Parasaurolophus cyrtocristatus.[8]
Parasaurolophus cyrtocristatus (Ostrom, 1961) wurde ebenfalls in New Mexico (Fruitland-Formation) und der Kaiparowits-Formation in Utah gefunden. Drei mögliche Funde sind bekannt. Sie war die kleinste Art und zeichnete sich durch einen kurzen und abgerundeten Knochenzapfen aus. Dies führte zu der Hypothese, dass es sich um ein Weibchen oder Jungtier von Parasaurolophus tubicen handeln könnte.[16] Als letztes wurde diese Idee von Thomas Williamson vertreten, welcher anmerkt, dass Parasaurolophus cyrtocristatus ungefähr 72 % der Größe von Parasaurolophus tubicen aufweist, dem etwa gleichen Größenverhältnis, welches auch bei anderen Lambeosaurinae zwischen erwachsenen Tieren und Jungtieren, die anfangen, deutlichen Geschlechtsdimorphismus auszubilden, vermutet wird.[16] In jüngeren Abhandlungen wurde diese Ansicht jedoch zurückgewiesen.[2][15] Alternativ könnten die anderen beiden Arten näher miteinander verwandt gewesen sein als mit Parasaurolophus cyrtocristatus, was die Größenabweichung dieser Art bedingt.[15]
Ein Hirnschädel eines Jungtiers aus dem Dinosaur Provincial Park von Alberta konnte keiner bestehenden Art zugeordnet werden.[13] Drei weitere Funde aus Alberta können eventuell ebenfalls Parasaurolophus zugeschrieben werden.[17]
Paläobiologie
Habitat
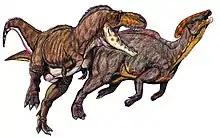
Parasaurolophus walkeri aus der Dinosaur-Park-Formation von Alberta war Teil einer reichhaltigen und gut dokumentierten Dinosaurierfauna, die Ceratopsier (Horndinosaurier) wie Centrosaurus, Styracosaurus und Chasmosaurus, weitere Hadrosaurier wie Prosaurolophus, Gryposaurus, Corythosaurus und Lambeosaurus, Ankylosaurier (gepanzerte Dinosaurier) wie Edmontonia und Euoplocephalus sowie den Tyrannosauriden Gorgosaurus umfasste.[18] Er war ein seltener Bestandteil dieser Fauna.[19] Die Landschaft war vermutlich von Flüssen und versumpften Auen durchzogen und das Klima war durch den Western Interior Seaway marin beeinflusst.[17] Das Klima war wärmer als im heutigen Alberta, ohne Frost im Winter, jedoch mit nassen und trockenen Jahreszeiten. Die Flora bestand aus Koniferen, die das Blätterdach bildeten, sowie aus Farnen, Baumfarnen und Bedecktsamern.[20]
Die Arten aus der Kirtland-Formation in New Mexico hingegen lebten mit dem großen Sauropoden Alamosaurus, dem Hadrosaurier Kritosaurus, dem Ceratopsier Pentaceratops, dem Ankylosaurier Nodocephalosaurus, dem kleinen Theropoden Saurornitholestes sowie bisher nicht näher bestimmten Tyrannosauriden zusammen.[18] Die Sedimente der Kirtland-Formation repräsentieren wahrscheinlich eine Überflutungsebene, die nach dem Rückzug des Western Interior Seaways entstand. Koniferen dominierten die Flora, und chasmosaurine Ceratopier (eine Gruppe gehörnter Dinosaurier) kamen anscheinend häufiger vor als Hadrosauriden.[21]
Ernährungsweise
Als Mitglied der Hadrosauridae war Parasaurolophus ein großer, zum vierbeinigen als auch zum zweibeinigen Laufen fähiger Herbivore, dessen fortgeschrittene Kiefer mahlende Kaubewegungen erlaubten. Die Kauleisten bildeten sogenannte Zahnbatterien, in denen mehrere hundert Zähne angeordnet waren, wobei immer nur eine Handvoll gleichzeitig benutzt wurde. Abgenutzte Zähne wurden ständig ersetzt. Pflanzenmaterial wurde erst mit dem Schnabel abgerupft und dann in wangenähnlichen Mundpartien gehalten. Sie konnten wahrscheinlich vom Grund bis in vier Meter Höhe weiden.[2] Der Paläontologe Bob Bakker sagte, Lambeosaurinae hätten schmalere Schnauzen als die Hadrosaurinae, der anderen Unterfamilie der Hadrosauridae. Dies führte zu der Theorie, dass Lambeosaurinae wählerischere Fresser waren als ihre breitschnabligen, meist kammlosen Gegenstücke.[22]
Knochenzapfen
Es wurden zahlreiche Hypothesen aufgestellt, welche die Funktionen des Knochenzapfens zu erklären versuchten – die meisten wurden jedoch verworfen.[23][4] Heute vermutet man mehrere Funktionen: Visuelle Funktionen (Identifikation von Arten und Geschlecht), Wirkung als Resonanzkörper für Rufe zur Kommunikation, und Temperaturregulierung. Es ist jedoch nicht klar, welche Funktion zu welcher Zeit in der Evolution die Bedeutendste war.[24]
Unterschiede zwischen Arten und Altersstufen
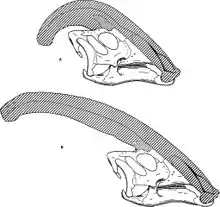
Wie bei anderen Lambeosaurinen geht man auch bei Parasaurolophus davon aus, dass sich die Kopfornamente mit dem Alter verändern und bei Männchen und Weibchen unterschiedlich sind (Geschlechtsdimorphismus).
James Hopson, einer der ersten Forscher, der die Ornamente der Lambeosaurinen in Bezug auf diese Unterschiede beschrieb, vermutete, dass Parasaurolophus cyrtocristatus, die kleinere Art mit dem kurzen, abgerundeten Knochenzapfen, ein Weibchen war.[10] Thomas Williamson meinte indes, es handele sich um ein Jungtier von Parasaurolophus tubicen.[16] Andere Forscher sehen Parasaurolophus cyrtocristatus hingegen als eigenständige Art. Keine dieser Ideen wurde weitestgehend akzeptiert – da insgesamt nur sechs Schädel und ein Hirnschädel eines Jungtieres bekannt sind, werden wohl nur weitere Funde Klarheit bringen können.
Williamson vermutet, dass die Jungtiere kleine Ornamente hatten, deren Wachstum sich in der Zeit direkt vor der Geschlechtsreife beschleunigte.[16] Eine neue Studie über den Hirnschädel eines Jungtiers, der zuvor Lambeosaurus zugeschrieben wurde, jetzt aber Parasaurolophus zugeordnet wird, gibt Hinweise auf einen kleinen Kamm bei juvenilen Tieren. Der Fund zeigt einen kleinen, vom paarigen Stirnbein (Frontale) gebildeten Wulst. Eine ähnliche Struktur ist auch von den Fossilien adulter Tiere bekannt, dort nur wesentlich größer – hier bilden die Frontalknochen eine Plattform, welche die Basis des Knochenzapfens unterstützt.[13]
Widerlegte Hypothesen über die Funktion

In der Vergangenheit gab es zahlreiche Hypothesen, welche die Funktion des Knochenzapfens zu erklären versuchten.[23][4] So gingen früher viele Forscher, in Bezug auf die bis in die 1960er verbreitete Hypothese, Hadrosaurier seien amphibisch, davon aus, es handle sich um eine Anpassung an eine aquatische Lebensweise. Beispielsweise schlug Alfred Sherwood Romer (1933) eine schnorchelähnliche Funktion des Knochenzapfens vor[25], während Martin Wilfarth (1947) eine Funktion als Halterung für einen Rüssel in Betracht nahm, welcher zum Atmen und zum Nahrungssammeln gedient haben sollte.[26] Charles M. Sternberg (1935) wiederum nahm an, es handle sich um eine Einrichtung, um mit Luft Wasser aus den Lungen zu halten[27], und Ned Colbert (1945) vermutete, der hohle Knochenzapfen diene als Luftreservoir um längere Tauchgänge zu ermöglichen.[28]
Andere Vermutungen waren mehr physischer Natur. Wie bereits oben erwähnt, glaubte der Erstbeschreiber William Parks an ein Ligament, welches das Knochenzapfenende mit den Rückenwirbeln verband – zur Stabilisierung des Kopfes.[7] Othenio Abel hielt den Knochenzapfen für eine Waffe, die bei Kämpfen zwischen zwei rivalisierenden Parasaurolophus zum Einsatz kam[29], und Angela Milner meinte, er könnte dafür verwendet worden sein, im Dickicht die Vegetation beiseite zu drücken, wie es der heutige Kasuar mit seinem Helm tut.[23] Weitere Deutungen sind z. B. die von Teresa Maryańska und Halszka Osmólska, die meinten, der Knochenzapfen beherberge Salzdrüsen[30], oder von John Ostrom, der, in Anbetracht der fehlenden Verteidigungsmöglichkeiten des Tieres, Riechgewebe vermutete, das den Tieren einen besseren Geruchssinn geben sollte.[31]
Die meisten dieser Hypothesen sind mit der Zeit in Verruf geraten.[4] So gab es z. B. weder ein Loch, das für eine Schnorchelfunktion notwendig war, noch Ansätze für die Muskeln eines Rüssels, der ohnehin bei einem mit einem Schnabel ausgestatteten Tier wenig Sinn hätte. Als Luftreservoir war der dazu zur Verfügung stehende Platz viel zu unbedeutend, und andere Hadrosaurier hatten ebenfalls große Köpfe, ohne dass sie diese durch Ligamente zu stützen brauchten. Auch lagen die Geruchsnerven und Geruchszellen außerhalb des Knochenzapfens, was zeigt, dass letzterer nichts mit dem Geruchssinn zu tun haben kann.[24] Auf der anderen Seite erklären diese Hypothesen nicht, warum der Knochenzapfen genau diese Form hatte oder warum andere Lambeosaurinen ganz andere Konstruktionen auf dem Kopf trugen. Stattdessen rücken visuelle und auditive Funktionen in den Fokus der Forscher.
Visuelle Funktionen
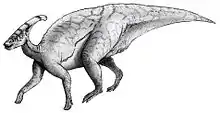
Als großes Objekt konnte der Knochenzapfen gut als visuelles Signal genutzt werden. Die Augen der Tiere waren recht groß und wiesen Skleralringe auf, was darauf hindeutet, dass die Tiere tagaktiv waren und der Sehsinn sehr wichtig war. Wahrscheinlich waren, wie auch bei anderen Lambeosaurinen vermutet wird, zwei visuelle Funktionen des Knochenzapfes gegeben: Auf der einen Seite die Funktion als Erkennungsmerkmal, damit die Tiere Artgenossen von anderen Hadrosaurierarten (wie z. B. Lambeosaurus oder Corythosaurus) unterscheiden konnten, und auf der anderen Seite die zur sexuellen Identifikation – eventuell zur Brautwerbung und zur Beeindruckung von Rivalen – ähnlich wie bei den rezenten Hirschen. Allerdings erklären diese Funktionen nicht die Komplexität der Röhrengänge im Inneren des Knochenzapfens, weshalb es noch eine weitere Funktion gegeben haben muss.[24]
Funktion als Resonanzkörper
Carl Wiman war der erste, der eine Lautsignal-Funktion für die Röhrengänge vorschlug (1931) – und verglich den Knochenzapfen mit einem Krummhorn.[32] Hopson und David B. Weishampel griffen diese Idee in den 1970ern und 1980ern wieder auf.[10][33][34] Hopson fand Hinweise auf ein empfindliches Gehör bei Hadrosauriden: So wurde bei dem verwandten Corythosaurus ein Steigbügel (ein Gehörknöchelchen) entdeckt, der, in Kombination mit einem großen Raum für das Trommelfell, ein empfindliches Mittelohr anzeigt. Des Weiteren ist die Lagena wie bei Krokodilen verlängert, was auf ein gut entwickeltes Innenohr hinweist.[10] Weishampel sagte voraus, Parasaurolophus walkeri wäre fähig gewesen, mit Hilfe des Knochenzapfens Frequenzen von 48 bis 240 Hz zu produzieren; für Parasaurolophus cyrtocristatus gab er 75 bis 375 Hz an. Weiter meinte er – basierend auf Ähnlichkeiten des Innenohrs mit dem von Krokodilen – erwachsene Hadrosauriden seien empfindlich gegenüber hohen Frequenzen, wie sie Jungtiere produziert haben könnten.[33]
Ein gut erhaltener Schädel von Parasaurolophus tubicen wurde am Computer modelliert, und, basierend auf dem 3D-Modell, der eventuelle Laut des Tieres, ein nebelhornartiges Geräusch, rekonstruiert.[35] Diese Art weist eine noch höhere Komplexität im Aufbau des Knochenzapfens auf als Parasaurolophus walkeri, der Hauptteil des Knochenzapfens erzeugte Töne von ca. 30 Hz.[36]
Funktion zur Kühlung
Eine andere Theorie besagt, es handele sich um eine Einrichtung zur Temperaturregulierung. Diese Idee, erstmals im Jahr 1978 vorgeschlagen, wird seit den 1990ern wieder stark diskutiert. P. E. Wheeler meinte zuerst, der Knochenzapfen sei ein Mittel zur Kühlung des Gehirns.[37] Etwa zur gleichen Zeit tauchte die Vorstellung einer Temperaturregulation durch den Knochenzapfen auch in einer Abhandlung von Maryańska und Osmólska auf. Weitere Beachtung fand die Theorie in einer Arbeit von Sullivan und Willamson über diese Gattung.[38] Als Indizien werden die große Oberfläche und Blutgefäße im Knochenzapfen genannt. In einer Diskussion von David Evans (2006) über die Funktionen der Lambeosaurinae-Kämme favorisierte dieser die Idee.[24]
Entdeckungsgeschichte
Der erste Fund wurde im Jahr 1920 im Rahmen einer Expedition der University of Toronto im Sand Creek am Red Deer River in Alberta (Kanada) gemacht.[7] Die Gesteinsschicht ist heute als Dinosaurier-Park-Formation bekannt und beinhaltet Fossilien aus dem Campanium. Das Exemplar (Holotyp; Exemplarnummer ROM 768) besteht aus einem Teilskelett mit Schädel, wobei die Beinknochen unterhalb der Kniegelenke und der Großteil des Schwanzes fehlen.

Im Jahr 1922 beschrieb William Parks das neue Tier als Parasaurolophus walkeri. Parks nannte die Gattung Parasaurolophus, was so viel wie „fast eine Kammechse“ (gr. sauros – Echse, lophos – Kamm) bedeutet, da er Saurolophus für einen nahen Verwandten hielt. Das Artepitheth walkeri ehrt Sir Byron Edmund Walker, einen Vorsitzenden des Kuratoriums des Royal Ontario Museums.[7] Parasaurolophus-Funde aus Alberta blieben jedoch selten[19] – so sind lediglich drei weitere Funde ohne Schädel aus der Dinosaurier-Park-Formation bekannt, die vielleicht zu Parasaurolophus gehören.[39] 2007 wurde der Hirnschädel eines Jungtiers (Exemplarnummer CMN 8502) neu untersucht und dieser Gattung zugeschrieben. Dieser wahrscheinlich aus der Dinosaurier-Park-Formation stammende Fund wurde bereits 1917 von Charles Mortram Sternberg gesammelt und 1924 von Gilmore als Lambeosaurus sp. beschrieben.[13]
Charles Hazelius Sternberg entdeckte im Jahr 1921 in der etwas jüngeren Kirtland-Formation in San Juan Country (New Mexico) einen fragmentarischen Schädel (Exemplarnummer PMU.R1250), der zur weiteren Untersuchung nach Uppsala (Schweden) gesandt wurde. 1931 beschrieb ihn Carl Wiman als eine zweite Art, Parasaurolophus tubicen.[32] Das Artepitheth tubicen ist aus dem lateinischen tǔbǐcěn abgeleitet und bedeutet „Trompeter“.[40] Ein zweiter, fast kompletter Schädel von Parasaurolophus tubicen (Exemplarnummer NMMNH P-25100) wurde 1995 in New Mexico entdeckt. Dieser Schädel wurde 1999 mittels Computertomografie untersucht und bot unter anderem Einblicke in die Funktionsweise des Knochenzapfens.[8]
John Ostrom beschrieb einen weiteren guten Fund (Exemplarnummer FMNH P27393) aus New Mexico im Jahr 1961 als Parasaurolophus cyrtocristatus. Das sich heute im Field Museum of Natural History befindende Exemplar besteht aus einem unvollständigen Schädel mit einem kurzen, abgerundeten Knochenzapfen sowie aus dem Großteil des postkranialen Skeletts (das Restskelett, das sich dem Schädel anschließt). Es fehlen die Füße, der Hals sowie Teile des Schwanzes.[41] Der Artname leitet sich von den lateinischen Wörtern curtus – „verkürzt“ und cristatus – „kammtragend“ ab.[40] Das Skelett wurde entweder im Top der Fruitland-Formation gefunden oder, was wahrscheinlicher wäre, in der darüberliegenden Kirtland-Formation.[8] 1979 wurde die Art auch in Utah nachgewiesen: David B. Weishampel und James A. Jensen beschrieben einen unvollständigen Schädel (Exemplarnummer BYU 2467) mit einem ähnlich kurzen Knochenzapfen aus den Schichten der Kaiparowits-Formation, Garfield County.[42] 1999 wurde ein weiterer Schädel aus Utah mit kurzem und abgerundeten Knochenzapfen beschrieben.[8] Ein 2013 beschriebenes Skelett eines juvenilen, noch nicht einmal ein Jahr alten Jungtiers zeigt nur einen kleinen Knochenkamm.[43]
In der Populärkultur
Mit seinen spektakulären Hornzapfen war Parasaurolophus Darsteller in vielen Filmen und Fernsehshows. Ein Beispiel ist die Kinderfilm-Serie „In einem Land vor unserer Zeit“ (The Land Before Time), in dem die Figur „Ducky“ offiziell als ein Parasaurolophus gilt, obwohl sie ein Saurolophus ist.[44] Weiter taucht dieser Dinosaurier im Hintergrund vieler Filme auf, wie im Abschnitt „Le sacre du Printemps“ aus Disneys „Fantasia“,[45] allen drei „Jurassic Park“-Filmen[46] und Disneys „Dinosaurier“[47]. Des Weiteren findet sich Parasaurolophus seit Jahrzehnten häufig als Dinosaurier-Spielzeug, z. B. von „Louis Marx and Company“, „MPC“, „Battat“, "Hancock" oder der Carnegie-Kollektion.
Weblinks
- John H Ostrom: Parasaurolophus cyrtocristatus, a crested hadrosaurian dinosaur from New Mexico (Volume Fieldiana, Geology, Vol. 14, No. 8) E-Text
- Skelettrekonstruktionen von Parasaurolophus walkeri und Parasaurolophus cyrtocristatus (Memento vom 19. Februar 2012 im Internet Archive), im gleichen Maßstab, auf „Skeletal Drawing.com“.
{kind=link}
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, ISBN 978-0-691-13720-9, S. 308, Online.
- John R. Horner, David B. Weishampel, Catherine A. Forster: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 438–463.
- Richard S. Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America (= Geological Society of America. Special Papers. Bd. 40, ISSN 0072-1077). Geological Society of America, New York NY 1942, S. 229.
- Donald F. Glut: Parasaurolophus. In: Donald F. Glut: Dinosaurs. The Encyclopedia. McFarland & Co, Jefferson NC u. a. 1997, ISBN 0-89950-917-7, S. 678–684.
- Richard S. Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America (= Geological Society of America. Special Papers. Bd. 40, ISSN 0072-1077). Geological Society of America, New York NY 1942, S. 209–213.
- Michael K. Brett-Surman, Jonathan R. Wagner: Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids. In: Kenneth Carpenter (Hrsg.): Horns and Beaks. Ceratopsian and ornithopod Dinosaurs. Indiana University Press, Bloomington IN 2007, ISBN 978-0-253-34817-3, S. 135–169.
- William A. Parks: Parasaurolophus Walkeri. A new genus and species of crested Trachodont dinosaur (= University of Toronto Studies. Geology Series. Bd. 13, ISSN 0372-4913). University of Toronto – University Library, Toronto 1922, Digitalisat.
- Robert M. Sullivan, Thomas E. Williamson: A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus (= New Mexico Museum of Natural History and Science. Bulletin. 15, ISSN 1524-4156). New Mexico Museum of Natural History and Science, Albuquerque NM 1999, Digitalisat.
- Douglas Palmer (Hrsg.): Dinosaurier & prähistorische Tiere. Eine illustrierte Enzyklopädie. Könemann, Köln 2000, ISBN 3-8290-6113-7, S. 153.
- James A. Hopson: The evolution of cranial display structure in hadrosaurian dinosaurs. In: Paleobiology. Bd. 1, Nr. 1, 1975, ISSN 0094-8373, S. 21–43.
- Charles W. Gilmore: On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks. In: Canada, Department of Mines, Geological Survey. Bulletin. Bd. 38, Nr. 43, 1924, ZDB-ID 429582-1, S. 29–48.
- Pascal Godefroit, Shuqin Zan, Liyong Jin: Charonosaurus jiayinensis n.g., n.sp.,a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China. In: Comptes Rendus de l'Académie des Sciences. Série 2, Fascicule A: Sciences de la Terre et des Planètes. Bd. 330, Nr. 12, 2000, ISSN 0764-4450, S. 875–882, doi:10.1016/S1251-8050(00)00214-7.
- David C. Evans, Robert R. Reisz, Kevin Dupuis: A juvenile Parasaurolophus (Ornithischia: Hadrosauridae) Braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. In: Journal of Vertebrate Paleontology. Bd. 27, Nr. 3, 2007, ISSN 0272-4634, S. 642–650, doi:10.1671/0272-4634(2007)27[642:AJPOHB]2.0.CO;2.
- David B. Weishampel, Jack R. Horner: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, Berkeley CA u. a. 1990, ISBN 0-520-06726-6, S. 534–561.
- David C. Evans, Robert R. Reisz: Anatomy and Relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta. In: Journal of Vertebrate Paleontology. Bd. 27, Nr. 2, 2007, S. 373–393, doi:10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2.
- Thomas E. Williamson: Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico. In: Spencer G. Lucas, Andrew B. Heckert (Hrsg.): Dinosaurs of New Mexico (= New Mexico Museum of Natural History and Science. Bulletin. 17). New Mexico Museum of Natural History and Science, Albuquerque NM 2000, S. 191–213, online.
- David A. Eberth: The Geology. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A spectacular ancient Ecosystem revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 54–82.
- David B. Weishampel, Paul M. Barrett, Rodolfo Coria, Jean Le Loeuff, Xing Xu, Xijin Zhao, Ashok Sahni, Elizabeth Gomani, Christopher R. Noto: Dinosaur distribution. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–683, hier S. 517–606.
- Michael J. Ryan, David C. Evans: Ornithischian Dinosaurs. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 312–348.
- Dennis R. Braman, Eva B. Koppelhus (Hrsg.): Campanian palynomorphs. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A spectacular ancient Ecosystem revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 101–130.
- Dale A. Russell: An odyssey in time. The dinosaurs of North America. NorthWord Press u. a., Minocqua WI u. a. 1989, ISBN 1-55971-038-1, S. 160–164.
- Robert T. Bakker: The Dinosaur Heresies. New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. William Morrow, New York NY 1986, ISBN 0-688-04287-2, S. 194.
- David B. Norman: Hadrosaurids II. In: David B. Norman: The Illustrated Encyclopedia of Dinosaurs. An Original and Compelling Insight into Life in the Dinosaur Kingdom. Crescent Books, New York NY 1985, ISBN 0-517-468905, S. 122–127.
- David C. Evans: Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs. In: Paleobiology. Bd. 32, Nr. 1, 2006, S. 109–125, doi:10.1666/0094-8373(2006)032[0109:NCHACC]2.0.CO;2.
- Alfred Sherwood Romer: Vertebrate Paleontology. University of Chicago Press, Chicago IL 1933, S. 491.
- Martin Wilfarth: Rüsseltragende Dinosaurier. In: Orion. Illustrierte Zeitschrift für Natur und Technik. Bd. 2, 1947, ZDB-ID 207436-9, S. 525–532.
- Charles M. Sternberg: Hooded Hadrosaurs of the Belly River series of the Upper Cretaceous (= National Museum of Canada. Geological Series. Bulletin. 52, ZDB-ID 429579-1 = National Museum of Canada. Bulletin. 77). Patenaude, Ottawa 1935.
- Edwin H. Colbert: The Dinosaur Book. The ruling Reptiles and their Relatives (= Man and Nature Publications. Handbook. Nr. 14). The American Museum of Natural History, New York NY 1945, Digitalisat.
- Othenio Abel: Die neuen Dinosaurierfunde in der Oberkreide Canadas. In: Die Naturwissenschaften. Bd. 12, Nr. 36, 1924, S. 709–716, doi:10.1007/BF01504818.
- Teresa Maryańska, Halszka Osmólska: Aspects of hadrosaurian cranial anatomy. In: Lethaia. Bd. 12, Nr. 3, 1979, ISSN 0024-1164, S. 265–273, doi:10.1111/j.1502-3931.1979.tb01006.x.
- John H. Ostrom: The cranial crests of hadrosaurian dinosaurs. In: Postilla. Bd. 62, 1962, ISSN 0079-4295, S. 1–29.
- Carl Wiman: Parasaurolophus tubicen n. sp. aus der Kreide in New Mexico (= Nova Acta Regiae Societatis Scientiarum Upsaliensis. Serie 4, Bd. 7, Nr. 5, ZDB-ID 210356-4). Almqvist & Wiksell, Uppsala 1931.
- David B. Weishampel: Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). In: Paleobiology. Bd. 7, Nr. 2, 1981, S. 252–261.
- David B. Weishampel: The nasal cavity of lambeosaurine hadrosaurids (Reptilia; Ornithischia); comparative anatomy and homologies. In: Journal of Paleontology. Bd. 55, Nr. 5, 1981, S. 1046–1057.
- Sandia National Laboratories: Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur. 5. Dezember 1997, archiviert vom Original am 17. August 2014; abgerufen am 24. Oktober 2014.
- Carl F. Diegert, Thomas E. Williamson: A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen. In: Journal of Vertebrate Paleontology. Bd. 18, Supplement zu Nr. 3 = Abstracts of Papers Fifty-Eighth Annual Meeting, Society of Vertebrate Paleontology, Snowbird Ski and Summer Resort Snowbird, Utah, September 30 – October 3, 1998, 1998, S. 38A.
- P. E. Wheeler: Elaborate CNS cooling structures in large dinosaurs. In: Nature. Bd. 275, Nr. 5679, 1978, S. 441–443, doi:10.1038/275441a0.
- Robert M. Sullivan, Thomas E. Williamson: A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications. In: Journal of Vertebrate Paleontology. Bd. 16, Supplement zu Nr. 3 = Abstracts of Papers Fifty-Sixth Annual Meeting, Society of Vertebrate Paleontology, American Museum of Natural History, New York, New York, October 16–19, 1996, 1996, S. 68A.
- Philip J. Currie, Dale A. Russell: Geographic and stratigraphic distribution of dinosaur remains. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 553.
- David P. Simpson: Cassell's Latin – English, English – Latin Dictionary. 5th edition. Cassell u. a., London u. a. 1979, S. 883.
- John Harold Ostrom: A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico. In: Journal of Paleontology. Bd. 35, Nr. 3, 1961, S. 575–577.
- David B. Weishampel, James A. Jensen: Parasaurolophus (Reptilia; Hadrosauridae) from Utah. In: Journal of Paleontology. Bd. 53, Nr. 6, 1979, S. 1422–1427.
- Andrew A. Farke, Derek J. Chok, Annisa Herrero, Brandon Scolieri, Sarah Werning: Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids. In: PeerJ., 22. Oktober 2013, 1:e182 doi:10.7717/peerj.182.
- Die „In einem Land vor unserer Zeit“-DVD (Memento vom 17. Februar 2009 im Internet Archive)
- Fantasia (1940). In: Movie Mistakes. Abgerufen am 24. Oktober 2014.
- Parasaurolophus. In: Park Pedia. Abgerufen am 24. Oktober 2014.
- Disney Dinosaur Interviews David Krentz. In: Dave Hone’s Archosaur Musings. Abgerufen am 24. Oktober 2014.