Prosaurolophus
Prosaurolophus („vor dem Saurolophus“, im Vergleich zu dem später lebenden Dinosaurier mit einem ähnlichen Kopfkamm) war eine Gattung der Vogelbeckendinosaurier aus der Gruppe der Hadrosauridae. Die Gattung enthält derzeit zwei valide Arten, von denen Fossilien von mindestens 25 Individuen in Nordamerika gefunden wurden.[2] Die Fossilien stammen aus dem Mittleren bis Oberen Campanium (Oberkreide). Die Skelette wurden in der Dinosaur Park Formation in Alberta, Kanada, und der etwa gleich alten Two Medicine Formation in Montana, USA, gefunden.
| Prosaurolophus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Prosaurolophus maximus im Royal Tyrrell Museum | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Mittleres bis Oberes Campanium)[1] | ||||||||||||
| 80,6 bis 72 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Prosaurolophus | ||||||||||||
| Brown, 1916 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||

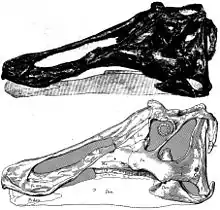
Merkmale
Prosaurolophus entsprach in seiner Anatomie den typischen Hadrosauriern. Er erreichte eine Körperlänge von acht bis neun Metern[3] und besaß einen sehr großen Kopf, der bei einem 8,5 Meter großen Exemplar 0,9 Meter lang war. Wie bei den meisten Hadrosauriern war der Vordere Bereich des Kopfes abgeflacht und schnabelartig verbreitert, wodurch er in der Lage war, Blätter von den Bäumen der nordamerikanischen Wälder abzubeißen. In der Mundhöhle besaß er tausende kleiner Zähne, durch die das Pflanzenmaterial vor dem Abschlucken zerkleinert wurde.
Als auffälligstes Merkmal besaß Prosaurolophus auf der Stirn vor den Augen einen vergleichsweise klein und dreieckig ausgebildeten Stirnkamm, der von den Nasenbeinen gebildet wurde und dessen Seiten durch ihre konkave Form Vertiefungen bildete. Die Vorderbeine waren im Vergleich zu anderen Hadrosauriern relativ kurz ausgebildet.
Die beiden Arten P. maximus und P. blackfeetensis unterschieden sich vor allem in den Proportionen des Schädels und der Größe des Stirnkamms. P. blackfeetensis besaß ein schmaleres und längeres Gesicht, zudem wanderte bei dieser Art der Stirnkamm während des Wachstums nach hinten in Richtung der Augen.
Fundstellen und Geschichte
Fossilien von Prosaurolophus sind aus Nordamerika von zwei Fundstellen bekannt, der Dinosaur Park Formation als oberster Schicht der Judith-River-Gruppe in Alberta, Kanada, und der etwa gleich alten Two Medicine Formation in Montana, USA. Von der Typusart P. maximus sind fossile Skelettüberreste von mindestens 25 Individuen sowie sieben vollständige Schädel bekannt, während von P. blackfeetensis ein teilweise vollständiges Skelett mit Schädel sowie mehrere Einzelknochen vorliegen.[3] Beide Fundgebiete zeichnen sich durch vielfältige Skelettfunde aus.
Die frühesten Fossilien der Typusart P. maximus wurden 1916 von dem amerikanischen Paläontologen Barnum Brown vom American Museum of Natural History beschrieben.[4] Er entdeckte 1915 einen Schädel in der Nähe des Red Deer River nahe der Stadt Steveville in Alberta, Kanada (Dinosaur Park Formation), der heute als AMNH 5836 im American Museum of Natural History liegt. Er beschrieb die Gattung 1916 als Prosaurolophus und nahm dabei Bezug auf den 1912 ebenfalls von ihm beschrieben Saurolophus.[5] Dieser besaß wie die neue Art einen auffälligen Stirnkamm, der allerdings länger und größer ausgebildet war. 1924 beschrieb William Parks ein fast vollständiges Skelett mit Schädel.[6] Bis heute sind aus Kanada und der Formation in Montana Fossilien von etwa 25 Individuen bekannt, teilweise in Form von fast vollständigen Skeletten und Schädeln.[2]
1992 beschrieb der Paläontologe Jack Horner vom Museum of the Rockies mit P. blackfeetensis eine zweite Art auf der Basis eines Skeletts, das in der Two Medicine Formation, genauer im Glacier County, entdeckt wurde.[7] Das Material wurde gemeinsam mit Überresten von drei oder vier weiteren Exemplaren als MOR 454 im Museum of the Rockies eingelagert. Die Fossilien wurden gemeinsam gefunden, sodass davon ausgegangen wird, dass sie gemeinsam gelebt haben und sich während einer Dürre nahe einer Wasserstelle sammelten.[8]
Systematik
|
Prosaurolophus bekam seinen Namen in Anlehnung an den Hadrosaurier Saurolophus, der einen ähnlich ausgebildeten Kamm besaß, in der Annahme, dass es sich um einen Vorfahren desselben handeln könnte (Vor-Saurolophus). Inwiefern tatsächlich eine engere Verwandtschaft zwischen den beiden Gattungen besteht, ist unklar; von einigen Autoren wird sie angenommen, andere ordnen beide Arten an unterschiedlichen Stellen der Systematik ein.
Nach der aktuellen Vorstellung wird Prosaurolophus in die Hadrosaurinae und damit in die nähere Verwandtschaft der Gattungen Brachylophosaurus, Edmontosaurus, Gryposaurus und Maiasaura gestellt.[9] Die Angehörigen dieser Gruppe zeichnen sich durch das Fehlen eines vollständigen Stirnkamms aus, der bei Saurolophus vorhanden ist.
Innerhalb der Gattung werden zwei valide Arten beschrieben, Nomina dubia sind nicht vorhanden. Die Typusart P. maxima wurde im Jahr 1916 erstmals von Barnum Brown aus der Two Medicines Formation beschrieben. 1992 beschrieb der Paläontologe Jack Horner vom Museum of the Rockies mit P. blackfeetensis eine zweite Art, die sich von P. maximus vor allem durch die Proportionen des Schädels sowie der Größe des Stirnkamms unterschied.[3]
Paläoökologie

Die Dinosaur Park Formation, in der Prosaurolophus maximus gefunden wurde, wird als ehemals flache Landschaft mit Flüssen und Überschwemmungsflächen betrachtet, die durch die Transgression des Western Interior Seaway nach Westen zunehmend sumpfiger wurde.[10] Das Klima war wärmer als im heutigen Alberta und ohne Frost, zugleich jedoch feuchter mit Trockenperioden. Die Vegetation bestand vor allem aus Nadelholzgewächsen mit einem Unterwuchs aus Farnen, Baumfarnen und Samenpflanzen.[11] Innerhalb dieser gut untersuchten Fossillagerstätte konnten Überreste von Prosaurolophus nur in der obersten Schicht gefunden werden, die mehr durch das Meer beeinflusst wurde als die unteren Schichten. Innerhalb dieser Schicht, deren Alter auf 76 bis 74 Millionen Jahre datiert wurde, stellt er den häufigsten Hadrosaurier dar.[12] Neben Prosaurolophus wurden zudem viele weitere Skelette von Dinosauriern gefunden, darunter die Ceratopsidae Centrosaurus, Styracosaurus und Chasmosaurus, der zu den Tyrannosauridae gehörende Gorgosaurus, die Ankylosauria Edmontonia und Euoplocephalus sowie mit Gryposaurus, Corythosaurus, Lambeosaurus und Parasaurolophus eine Reihe weiterer Hadrosaurier.[13]
Die zeitlich etwa gleich eingeordnete Two Medicine Formation, in der P. blackfeetensis entdeckt wurde, ist vor allem bekannt für die Fossilfunde von Dinosauriernestern, -eiern und Jungtieren der Hadrosaurier Hypacrosaurus stebingeri und Maiasaura sowie dem Troodontiden Troodon. Zudem wurden Skelettreste des Tyrannosauriden Daspletosaurus, dem Caenagnathiden Chirostenotes, den Dromaeosauriden Bambiraptor und Saurornitholestes, den Ankylosauriern Edmontonia und Euoplocephalus, dem Hypsilophodontiden Orodromeus sowie den Ceratopsiden Achelousaurus, Brachyceratops, Einiosaurus und Styracosaurus ovatus.[13] Die Fundstelle war weiter von der Western Interior Seaway entfernt, sie lag zudem höher und war trockener als die Dinosaur Park Formation.[8]
Paläobiologie

Ernährung
Prosaurolophus war, wie alle Hadrosaurier, ein großer Pflanzenfresser, der durch seinen schnabelähnlich ausgezogenen und komplexen Schädel in der Lage war, Pflanzenteile durch eine Bewegung ähnlich dem Kauen zu zerkleinern. Seine Zähne wurden kontinuierlich ersetzt und lagen in Zahnpaketen von mehreren hundert Zähnen im Kiefer, von denen nur ein Bruchteil gleichzeitig genutzt wurde. Das Pflanzenmaterial wurde mit seinen breiten Schnabel von Pflanzen vom Boden bis in etwa vier Meter Höhe abgerissen und verblieb durch wangenähnliche Bildungen im Mundraum. Wie andere Hadrosaurier bewegte sich auch diese Art wahrscheinlich vierfüßig (quadruped) und zweifüßig (biped) fort.[9]
Sozialverhalten
Dadurch, dass die Fossilfunde in Knochenlagern mit mehreren Skeletten gefunden wurden, wird abgeleitet, dass diese Tiere zumindest teilweise in Gruppen lebten.[8] Über das Schädelskelett gibt es weitere Annahmen für das Sozialverhalten: Der knöcherne Stirnkamm könnte eine Funktion als visuelles Kommunikationsmittel gehabt haben, zudem wird die Existenz von sackförmigen und häutigen Divertikeln an der Nasenhöhle angenommen, die neben der optischen Funktion auch akustisch der Kommunikation gedient haben könnten.Additionally, it had several potential methods for display in a social setting.[14]
Literatur
- David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 1st paperback edition. University of California Press, Berkeley CA u. a. 1992, ISBN 0-520-06726-6.
Weblinks
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, ISBN 978-0-691-13720-9, S. 299–300 princeton.edu.
- nach David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 1st paperback edition. University of California Press, Berkeley CA u. a. 1992, ISBN 0-520-06726-6, S. 557.
- Prosaurolophus (Memento vom 4. Oktober 2007 im Internet Archive), Datenblatt auf dinoruss.org.
- Veröffentlicht als: Barnum Brown: A new crested trachodont dinosaur Prosaurolophus maximus. In: Bulletin of the American Museum of Natural History. Bd. 35, Article 37, 1916, ISSN 0003-0090, S. 701–708, hdl:2246/2246/1328.
- Veröffentlicht als: Barnum Brown: A crested dinosaur from the Edmonton Cretaceous. In: Bulletin of the American Museum of Natural History. Bd. 31, Article 14, 1912, S. 131–136, hdl:2246/1401.
- Veröffentlicht als: William A. Parks: Dyoplosaurus acutosquameus. A new genus and species of armored dinosaur and notes on a skeleton of Prosaurolophus maximus (= University of Toronto Studies. Geological Series. Nr. 18, ISSN 0372-4913). University of Toronto – University Library, Toronto 1924.
- Veröffentlicht als: John R. Horner: Cranial Morphology of Prosaurolophus (Ornithischia: Hadrosauridae). With descriptions of two new hadrosaurid species and an evaluation of hadrosaurid phylogenetic relationships (= Museum of the Rockies Occasional Paper. Nr. 2). Montana State University – Museum of the Rockies u. a., Bozeman MT u. a. 1992, ISBN 1-56044-156-9.
- Raymond R. Rogers: Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation of northwestern Montana; evidence for drought-related mortality. In: Palaios. Bd. 5, Nr. 5, 1990, ISSN 0883-1351, S. 394–413, doi:10.2307/3514834.
- John R. Horner, David B. Weishampel, Catherine A. Forster: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 438–463.
- David A. Eberth: The Geology. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A spectacular ancient Ecosystem revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 54–82.
- Dennis R. Braman, Eva B. Koppelhus: Campanian palynomorphs. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A spectacular ancient Ecosystem revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 101–130.
- Michael J. Ryan, David C. Evans: Ornithischian Dinosaurs. In: Philip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, S. 312–348.
- David B. Weishampel, Paul M. Barrett, Rodolfo Coria, Jean Le Loeuff, Xing Xu, Xijin Zhao, Ashok Sahni, Elizabeth Gomani, Christopher R. Noto: Dinosaur distribution. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–683, hier S. 517–606.
- James A. Hopson: The evolution of cranial display structure in hadrosaurian dinosaurs. In: Paleobiology. Bd. 1, Nr. 1, 1975, ISSN 0094-8373, S. 21–43.