Hypacrosaurus
Hypacrosaurus ist eine Gattung ornithopoder Dinosaurier aus der Gruppe der Hadrosaurier. Sie ähnelte dem verwandten Corythosaurus – beide Gattungen zeigten einen großen, abgerundeten hohlen Kamm auf dem Kopf, wobei der Kamm von Hypacrosaurus nicht ganz so groß und gerade war. Es wurden zwei Arten benannt, die während der Oberkreide vor 75 bis 67 Millionen Jahren in Nordamerika lebten. Fossilien wurden in Alberta (Kanada) und in Montana (USA) gefunden.
| Hypacrosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Hypacrosaurus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Campanium bis Maastrichtium)[1] | ||||||||||||
| 75 bis 67 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Hypacrosaurus | ||||||||||||
| Brown, 1913 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Hypacrosaurus ist der letzte Vertreter der Lambeosaurinae (Hadrosaurier mit hohlem Knochenkamm) in Nordamerika, der durch vollständige Fossilfunde bekannt ist. Die Typusart Hypacrosaurus altispinus wurde erstmals 1913 von Barnum Brown wissenschaftlich beschrieben.
Beschreibung
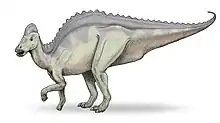
Hypacrosaurus lässt sich von anderen Lambeosaurinae durch die Form des Kammes sowie durch die hohen Dornfortsätze der Wirbel unterscheiden. Die Dornfortsätze der Rückenwirbel sind 5 bis 7 Mal so hoch wie die jeweiligen Wirbelkörper[2], was zu einem hohen Rücken führte. Der hohle Kopfkamm ähnelt dem des Corythosaurus, läuft zur Oberseite jedoch spitzer zu, ist nicht so hoch, breiter und besitzt einen kleinen knöchernen Wulst an der Hinterseite.[2] Im Gegensatz zu anderen Lambeosaurinae verliefen die Atemwege innerhalb des Kammes nicht in Form einer S-Kurve.[3] Das Tier erreichte wahrscheinlich eine Körperlänge von 9,1 m[2], und wurde etwa 4 Tonnen schwer.[4] Das Postkranialskelett unterscheidet sich nur geringfügig von dem anderer Hadrosaurier, obwohl einige Merkmale am Beckengürtel bei Hypacrosaurus einzigartig sind.[5] Wie alle Hadrosaurier war Hypacrosaurus herbivor und lief sowohl auf zwei als auch auf vier Beinen. Die beiden benannten Arten, Hypacrosaurus altispinus und Hypacrosaurus stebingeri, lassen sich von anderen Gattungen nicht durch einzelne, einzigartige Merkmale (Autapomorphien) unterscheiden, da Hypacrosaurus stebingeri als eine Übergangsform zwischen dem früheren Lambeosaurus und dem späteren Hypacrosaurus betrachtet wird.[6]
Systematik
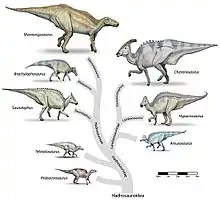
Innerhalb der Hadrosauridae wird Hypacrosaurus zu den Lambeosaurinae gezählt.[7] Die nächsten Verwandten schließen Lambeosaurus und Corythosaurus mit ein.[4] Jack Horner und Philip Currie (1994) vermuten, das Hypacrosaurus stebingeri eine Übergangsform zwischen Lambeosaurus und Hypacrosaurus altispinus darstellt.[6] Eine andere Studie von Michael K. Brett-Surman (1989) hält Hypacrosaurus für identisch mit Corythosaurus.[8] Suzuki und Kollegen (2004) schlagen in ihrer Neubeschreibung von Nipponosaurus eine enge Verwandtschaft zwischen Nipponosaurus und Hypacrosaurus stebingeri vor und halten Hypacrosaurus für paraphyletisch.[9] Eine jüngere Studie von Evans und Reisz (2007) widerspricht dieser Hypothese jedoch und sieht Hypacrosaurus innerhalb einer Klade, die Corythosaurus und Olorotitan als engste Verwandte umfasst, nicht aber Nipponosaurus.[10]
Das hier dargestellte Kladogramm folgt Evans und Reisz (2007) (vereinfacht):[10]
| Hadrosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Forschungsgeschichte und Namensgebung

Das Holotyp-Exemplar wurde bei einer Grabung unter der Leitung des berühmten Paläontologen Barnum Brown im Jahr 1910 für das American Museum of Natural History geborgen.[5] Der Fund, ein fragmentarisches postkraniales Skelett, das aus mehreren Wirbeln und einem teilweisen Becken besteht (Exemplarnummer AMNH 5204), wurde am Red Deer River nahe Tolman Ferry in Alberta (Kanada) entdeckt. Der geologische Formation, aus der der Fund stammt, ist heute als Horseshoe-Canyon-Formation bekannt und wird auf das frühe Maastrichtium datiert. Brown beschrieb diese Überreste 1913 zusammen mit weiteren postkranialen Knochen als neue Gattung, die er sich als ähnlich mit Saurolophus vorstellte.[11] Zur Zeit der Erstbeschreibung war kein Schädel bekannt; zwei Schädel wurden jedoch bald entdeckt und beschrieben.[7]
Der Name Hypacrosaurus (gr. υπο-, hypo- = „kleiner“, „geringer“; ακρος, akros = „hoch“ + σαυρος, sauros = „Echse“) bedeutet so viel wie „beinahe die höchste Echse“ und soll auf ein Tier hinweisen, das fast, aber nicht ganz so groß wie Tyrannosaurus war.[11][12]
In der Folgezeit wurden mehrere Taxa besonders kleiner Lambeosaurinen beschrieben, die heute jedoch als Jungtiere verschiedener anderer Lambeosaurinen betrachtet werden. So wurde die Gattung Cheneosaurus 1917 von Lawrence Lambe aufgestellt, basierend auf einem teilweisen Skelett samt Schädel, Beinknochen, Wirbeln und Beckenknochen aus der Horseshoe-Canyon-Formation.[13] Drei Jahre nach der Beschreibung von Cheneosaurus identifizierten Richard Swann Lull und Nelda Wright ein Skelett aus dem American Museum of Natural History (AMNH, Exemplarnummer 5461) aus der Two-Medicine-Formation von Montana als ein Exemplar von Procheneosaurus.[14] Diese und andere Taxa wurden bis in die 1970er als gültig akzeptiert, als Peter Dodson zeigte, dass es sich um Jungtiere anderer etablierter Lambeosaurinen handelte. Dodson vermutete, dass die "Cheneosaurus"-Exemplare wahrscheinlich als Jungtiere des zeitgenössischen Hypacrosaurus altispinus zu betrachten sind.[15] Spätere Veröffentlichungen folgten dieser Hypothese.[4] Das Procheneosaurus-Exemplar aus der Two-Medicine-Formation gilt heute als Vertreter der zweiten, 1994 beschriebenen Hypacrosaurus-Art, Hypacrosaurus stebingeri.[6]
Arten

Hypacrosaurus altispinus, die Typusart, ist von 5 bis 10 im anatomischen Verbund entdeckten Schädeln und assoziierten Teilen des Postkraniums bekannt. Hypacrosaurus stebingeri ist von Überresten bekannt, die zu zahlreichen verschiedenen Individuen gehören und alle Altersstufen umfassen, von Embryonen bis erwachsenen Tieren.[4]
Paläoökologie
Hypacrosaurus altispinus stammt aus der Horseshoe-Canyon-Formation und teilte sich seinen Lebensraum mit den Hadrosauriern Edmontosaurus und Saurolophus, dem kleinen ursprünglichen Ornithopoden Parksosaurus, dem Ankylosauriden Euoplocephalus, dem Nodosauriden Edmontonia, den gehörnten Dinosauriern Montanoceratops, Anchiceratops, Arrhinoceratops und Pachyrhinosaurus, dem Pachycephalosaurier Stegoceras, den straußenartigen Ornithomimosauriern Ornithomimus und Struthiomimus, verschiedenen wenig bekannten, kleinen Theropoden inklusive Troodontiden und Dromaeosauriden, sowie mit den Tyrannosauriden Albertosaurus und Daspletosaurus.[16] Die Horseshoe-Canyon-Formation ist wegen des immer wieder vorgedrungenen Western Interior Seaway, einem Meeresarm, der während der Kreide das mittlere Nordamerika bedeckte, deutlich marin geprägt.[17] Hypacrosaurus altispinus lebte vermutlich mehr im Landesinneren.[4]
In der etwas älteren Two-Medicine-Formation, aus der die Fossilien von Hypacrosaurus stebingeri stammen, waren Hadrosaurier auch durch die Gattungen Maiasaura und Prosaurolophus vertreten. Troodontide Theropoden sind durch Troodon repräsentiert. Weitere Theropoden waren der Tyrannosauride Daspletosaurus, der Oviraptoride Chirostenotes sowie die Dromaeosauriden Bambiraptor und Saurornitholestes. Herbivoren waren mit den Ankylosauriern Edmontonia und Euoplocephalus, dem ursprünglichen Ornithopoden Orodromeus und den Ceratopsier Achelousaurus, Brachyceratops, Einiosaurus und Styracosaurus vertreten.[16] Der Ablagerungsraum dieser Formation war weiter vom Western Interior Seaway entfernt, lag höher und trockener und war stärker terrestrisch geprägt.[18]
Paläobiologie
Nester und Entwicklungsbiologie

Von der Art Hypacrosaurus stebingeri konnte eine Vielzahl von fossilen Eiern, Nestern und Jungtieren beschrieben werden. Diese Eier waren annähernd rund und 20 mal 18,5 Zentimeter groß und enthielten Embryonen mit einer Körperlänge von etwa 60 Zentimetern. Frisch geschlüpfte Tiere waren ca. 1,7 Meter lang. Jungtiere wiesen tiefe Schädel auf, deren Oberseite nur eine leichte Erhöhung derjenigen Knochen zeigte, die beim erwachsenen Tier den Schädelkamm formten.[6] Anhand von Ringen in Querschnitten der Langknochen, die von Zeiten verminderten Wachstums herrühren, und der Dicke dieser Knochen ließ sich das Wachstumsmuster dieses Dinosauriers rekonstruieren. So war das Wachstum in den ersten Jahren sehr schnell und mit dem eines Laufvogels vergleichbar. Nach dieser starken Wachstumsphase war das Wachstum nicht abgeschlossen, sondern setzte sich deutlich langsamer fort.[19] Untersuchungen von Lisa Cooper und Kollegen an Hypacrosaurus stebingeri zeigen, dass diese Tiere die Geschlechtsreife schon mit einem Alter von 2 bis 3 Jahren und die volle Größe mit etwa 10 bis 12 Jahren erreichten. Der Umfang des Oberschenkelknochen betrug zum vermuteten Zeitpunkt der Geschlechtsreife etwa 40 % des endgültigen Umfangs bei einem vollständig ausgewachsenen Tier. Die vermutete Wachstumsrate von Hypacrosaurus stebingeri übertraf die von Tyrannosauriden wie Albertosaurus und Tyrannosaurus, mutmaßlichen Prädatoren von Hypacrosaurus, und könnte sich aufgrund des Prädatorendrucks herausgebildet haben, um möglichst schnell eine zur Verteidigung nützliche Körpergröße zu erreichen. Auch das frühe Erreichen der Geschlechtsreife wäre für ein Beutetier von Vorteil gewesen.[20]
Funktion des Kopfkamms

Der hohle Kopfkamm von Hypacrosaurus hatte, wie bei anderen Lambeosaurinen, vermutlich soziale Funktionen. Er könnte als visuelles Erkennungsmerkmal gedient haben, das Individuen erlaubte, Vertreter eines bestimmten Geschlechts oder der eigenen Spezies zu erkennen. Zudem diente er vermutlich als Resonanzkörper zur Erzeugung von Lautsignalen.[4]
Forscher um John Ruben (1996) ziehen den Kopfkamm von Hypacrosaurus als ein Argument gegen die verbreitete Forschungsmeinung heran, Dinosaurier seien endotherm (warmblütig) gewesen. Wie diese Forscher anhand von Computertomographie-Scans feststellen, fehlte Hypacrosaurus und zwei anderen von diesen Forschern untersuchten Dinosaurier-Spezies wahrscheinlich ein bestimmter Typ von Nasenmuscheln, der die Funktion hat, Wasserverlust beim Atmen zu verhindern. Dieser Typ von Nasenmuscheln ist heute nur bei endothermen Tieren – Säugetieren und Vögel – zu finden, nicht aber bei ektothermen Tieren wie Reptilien. Endotherme Tiere würden ohne derartige Nasenmuscheln zu viel Wasser verlieren, da sie wegen ihres höheren Stoffumsatzes mehr Atemzüge tätigen müssen.[21][22]
Einzelnachweise
- Victoria M. Arbour, Michael E. Burns, Robin L. Sissons: A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus. In: Journal of Vertebrate Paleontology. Bd. 29, Nr. 4, 2009, ISSN 0272-4634, S. 1117–1135, doi:10.1671/039.029.0405.
- Richard S. Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America (= Geological Society of America. Special Paper. Bd. 40, ISSN 0072-1077). Geological Society of America, New York NY 1942, S. 206–208.
- David B. Weishampel: The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies. In: Journal of Paleontology. Bd. 55, Nr. 5, 1981, ISSN 0022-3360, S. 1046–1057.
- John R. Horner, David B. Weishampel, Catherine A. Forster: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 438–463.
- Donald F. Glut: Dinosaurs. The Encyclopedia. McFarland, Jefferson NC u. a. 1997, ISBN 0-89950-917-7, S. 478–482.
- John R. Horner, Philip J. Currie: Embryonic and neonatal morphology and ontogeny of a new species of Hypacrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta. In: Kenneth Carpenter, Karl F. Hirsch, John R. Horner (Hrsg.): Dinosaur Eggs and Babies. Cambridge University Press, Cambridge 1994, ISBN 0-521-44342-3, S. 312–336.
- Charles W. Gilmore: On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks. In: Canada, Department of Mines, Geological Survey. Bulletin. Bd. 38, Nr. 43, 1924, ZDB-ID 429582-1, S. 29–48.
- Michael Keith Brett-Surman: A Revision of the Hadrosauridae (Reptilia: Ornithischia) and their Evolution during the Campanian and Maastrichtian. The Faculty of The Graduate School of Arts and Sciences of The George Washington University, Washington DC 1989 (Dissertation).
- Daisuke Suzuki, David B. Weishampel, Nachio Minoura: Nipponosaurus sachaliensis (Dinosauria; Ornithopoda): anatomy and systematic position within Hadrosauridae. In: Journal of Vertebrate Paleontology. Bd. 24, Nr. 1, 2004, S. 145–164, doi:10.1671/A1034-11.
- David C. Evans, Robert R. Reisz: Anatomy and Relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta. In: Journal of Vertebrate Paleontology. Bd. 27, Nr. 2, 2007, S. 373–393, doi:10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2.
- Barnum Brown: A new trachodont dinosaur, Hypacrosaurus, from the Edmonton Cretaceous of Alberta. In: Bulletin of the American Museum of Natural History. Bd. 32, Article 20, 1913, ISSN 0003-0090, S. 395–406, online.
- Benjamin S. Creisler: Deciphering duckbills. In: Kenneth Carpenter (Hrsg.): Horns and Beaks. Ceratopsian and ornithopod Dinosaurs. Indiana University Press, Bloomington IN 2007, ISBN 978-0-253-34817-3, S. 185–210.
- Lawrence M. Lambe: On Cheneosaurus tolmanensis, a new genus and species of trachodont dinosaur from the Edmonton Cretaceous of Alberta. In: The Ottawa Naturalist. Bd. 30, Nr. 10, 1917, ISSN 0316-4411, S. 117–123, Digitalisat.
- William D. Matthew: Canadian dinosaurs. In: Natural History. Bd. 20, Nr. 5, 1920, ISSN 0028-0712, S. 536–544, Digitalisat.
- Peter Dodson: Taxonomic Implications of Relative growth in Lambeosaurine Hadrosaurs. In: Systematic Zoology. Bd. 24, Nr. 1, 1975, ISSN 0039-7989, S. 37–54, doi:10.1093/sysbio/24.1.37.
- David B. Weishampel, Paul M. Barrett, Rodolfo Coria, Jean Le Loeuff, Xing Xu, Xijin Zhao, Ashok Sahni, Elizabeth Gomani, Christopher R. Noto: Dinosaur distribution (Late Triassic, Africa). In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–683, hier S. 517–606.
- Peter Dodson: The Horned Dinosaurs. A Natural History. Princeton University Press, Princeton NJ 1996, ISBN 0-691-02882-6, S. 14–15.
- Raymond R. Rogers: Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation of northwestern Montana; evidence for drought-related mortality. In: PALAIOS. Bd. 5, Nr. 5, 1990, ISSN 0883-1351, S. 394–413, doi:10.2307/3514834.
- Lisa N. Cooper, John R. Horner: Growth rate of Hypacrosaurus stebingeri as hypothesized from lines of arrested growth and whole femur circumference. In: Journal of Vertebrate Paleontology. Bd. 19, Supplement zu Nr. 3 = Abstracts of Papers Fifty-Ninth Annual Meeting Society of Vertebrate Paleontology, Adams Mark Hotel, Denver, Colorado October 20–23, 1999, 1999, S. 35A.
- Lisa Noelle Cooper, Andrew H. Lee, Mark L. Taper, John R. Horner: Relative growth rates of predator and prey dinosaurs reflect effects of predation. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 275, Nr. 1651, 2008, ISSN 0080-4649, S. 2609–2615, doi:10.1098/rspb.2008.0912, PMID 18682367, PMC 2605812 (freier Volltext).
- John A. Ruben, Willem J. Hillenius, Nicholas R. Geist, Andrew Leitch, Terry D. Jones, Philip J. Currie, John R. Horner, George Espe III: The metabolic status of some Late Cretaceous dinosaurs. In: Science. Bd. 273, Nr. 5279, 1996, S. 1204–1207, doi:10.1126/science.273.5279.1204.
- Anusuya Chinsamy, Willem J. Hillenius: Physiology of nonavian dinosaurs. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 643–659.