Gemeiner Vampir
Der Gemeine Vampir (Desmodus rotundus) ist eine auf dem amerikanischen Kontinent lebende Fledermausart. Diese Spezies ist die bekannteste – und auch am besten erforschte – der drei Arten der Vampirfledermäuse (Desmodontinae), der einzigen Säugetiergruppe, die sich ausschließlich vom Blut anderer Tiere ernährt. Gemeine Vampire sind als Überträger von Krankheiten wie der Tollwut auf Nutztiere und Menschen gefürchtet, gleichzeitig liefert ein gerinnungshemmendes Enzym in ihrem Speichel wichtige Impulse für die medizinische Forschung.
| Gemeiner Vampir | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Gemeiner Vampir (Desmodus rotundus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Desmodus | ||||||||||||
| Wied-Neuwied, 1826 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Desmodus rotundus | ||||||||||||
| (E. Geoffroy, 1810) |
Beschreibung
Körperbau und Abmessungen



Gemeine Vampire sind mittelgroße Fledermäuse: Sie erreichen eine Kopfrumpflänge von 70 bis 90 Millimetern und ein Gewicht von 15 bis 50 Gramm. Die Unterarmlänge, eine wichtige Größenangabe bei Fledertieren, beträgt zwischen 50 und 63 Millimetern und die Flügelspannweite zwischen 35 und 40 Zentimeter. Im Körperbau stimmt der Gemeine Vampir weitgehend mit den anderen Fledermäusen überein: die Flugmembran besteht aus zwei Hautschichten und erstreckt sich von den Handgelenken bis zu den Fußgelenken, gespannt wird sie von den stark verlängerten Fingern der Vordergliedmaßen. Weitere Flughäute erstrecken sich von den Schultern zum Handgelenk und zwischen den Beinen. Letztere wird Uropatagium genannt und dient bei vielen Fledermausarten zum Insektenfang. Da diese Funktion beim Gemeinen Vampir wegfällt, ist sie bei dieser Art klein und unbehaart. Ein Schwanz ist nicht vorhanden. Die Hinterbeine sind im Gegensatz zu vielen anderen Fledermausarten stark und muskulös und ermöglichen den Tieren unter anderem ein schnelles Vorwärtskrabbeln auf dem Boden. Vampirfledermäuse können darüber hinaus auch hüpfen (siehe Lokomotion). Das Fell dieser Tiere ist an der Oberseite graubraun gefärbt, die Unterseite ist etwas heller.
Von den anderen Vampirfledermaus-Arten unterscheidet sich der Gemeine Vampir durch die verlängerten Daumen, die fehlende Behaarung des Uropatagiums, die spitzeren Ohren sowie die Anzahl der Zähne.
Gesicht und Zähne

Die Schnauze des Gemeinen Vampirs ist kurz und kegelförmig. Sie hat im Gegensatz zu vielen anderen Blattnasen kein echtes Nasenblatt, sondern einen hufeisenförmigen Ballen über den Nasenlöchern. Die Ohren sind spitz und eher klein und enthalten einen kleinen lanzettförmigen Tragus (Ohrdeckel). Die Augen hingegen sind für Fledermausverhältnisse relativ groß.
Der Gemeine Vampir hat mit 20 die wenigsten Zähne der Vampirfledermäuse, seine Zahnformel lautet 1/2-1/1-2/3-0/0 x2. Am größten sind die Schneide- und Eckzähne, sie sind sichelförmig und an das Aufschneiden der Haut ihrer Opfer angepasst. Die Backenzähne sind rückgebildet und haben keinerlei zum Kauen geeignete Oberfläche mehr. An der Unterseite der Zunge finden sich zwei Rillen, die sich während der Mahlzeit zusammenziehen und ausdehnen. Weitere Anpassungen an die spezielle Ernährung sind die kurze Speiseröhre und der schlauchförmige Magen.
Lokomotion

Bestimmte Anpassungen im Körperbau und im Bewegungsverhalten des Gemeinen Vampirs werden von Fachleuten als „spektakulär“ bezeichnet[1]. So kann Desmodus rotundus zum Beispiel mit zusammengefalteten Flughäuten auf dem Erdboden stehen und darüber hinaus eine Art hüpfend-springende Fortbewegung ausführen. Dabei laufen die Tiere auf ihrem extrem verlängerten „Daumen“ (1. Finger). Diese Fledermäuse sind darüber hinaus in der Lage, aus dem Stand blitzschnell „in den Flug“ abzuspringen, selbst wenn sie eine Blutmahlzeit aufgenommen und dadurch ihr Körpergewicht erheblich erhöht haben. Sie vermögen schließlich aus dem Flug auf dem Erdboden zu landen und sofort in den Stillstand zu kommen. Der Gemeine Vampir besitzt artspezifische Besonderheiten in der Skelett- und Muskelanatomie, etwa besonders kräftige vordere Extremitäten (in Form der zusammengefalteten Flughäute), die solche Leistungen möglich machen[2][3] Diese Anpassungen befähigen diese Vampirfledermäuse, neben einem großen Beutetier (Rind, Pferd) auf dem Boden zu landen und sich diesem im Laufen zu nähern; während des Fressvorgangs kann Desmodus langsamen Bewegungen der Beute folgen oder aber blitzschnell auf Abwehrbewegungen reagieren.
Verbreitung und Lebensraum
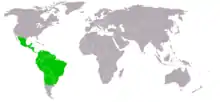
Gemeine Vampire leben auf dem amerikanischen Kontinent, ihr Verbreitungsgebiet reicht vom nördlichen Mexiko bis ins südliche Südamerika (Zentralchile, Argentinien und Uruguay). Sie finden sich auch auf einigen Südamerika vorgelagerten Inseln (wie Margarita und Trinidad), fehlen aber auf den anderen Westindischen Inseln. Sie bewohnen sowohl feuchte als auch trockene Gebiete der Tropen und Subtropen, darunter Wälder, offene Grasländer und Gebirgsregionen bis in 2400 m Höhe.
Der einzige Zoo in Deutschland, der Gemeine Vampire hält, ist der Zoologische Garten in Berlin.[4]
Lebensweise
Aktivitätszeiten und Gruppenleben

Gemeine Vampire sind ausschließlich nachtaktiv, als Schlafquartiere nutzen sie in erster Linie Höhlen, daneben findet man sie auch in hohlen Bäumen, Minen und Schächten sowie in verlassenen Gebäuden. Ruheplätze der Vampirfledermäuse riechen wegen des heraufgewürgten Blutes oft streng nach Ammoniak.
Diese Tiere leben in Gruppen von 20 bis 100 Tieren, gelegentlich bilden sie auch Kolonien von bis zu 2000 Tieren. Diese Gruppen zerfallen in Haremsgruppen, die aus 8 bis 20 Weibchen samt ihrem Nachwuchs sowie einem Männchen, das sich in deren Nähe aufhält, bestehen. Andere Männchen bilden Junggesellengruppen. Immer wieder versuchen männliche Tiere, dem Haremsführer seinen Rang streitig zu machen, in diesem Fall kann es zu wilden Kämpfen kommen. Die Kämpfe können auch durch aggressive Laute begleitet werden.
Sinnesfunktionen und Orientierung
Bei der Orientierung zeigen Vampirfledermäuse ähnliche Fähigkeiten und Leistungen wie verwandte Fledermausarten; allerdings hat die Forschung charakteristische Besonderheiten aufgedeckt, die als spezifische Anpassung an die blutfressende (sanguivore) Ernährungsweise interpretiert werden können.
Die Echoortung bei Desmodus ähnelt der anderer Arten aus der Familie der Blattnasenfledermäuse (Phyllostomatidae) und kann als eher unspezialisiert gelten. Die Ultraschall-Ortungslaute werden durch das Maul ausgestoßen; sie weisen zwei intensive Harmonische (Obertöne) auf, die insgesamt einen Frequenzbereich zwischen ca. 43 und 100 Kilohertz (kHz) abdecken. In Ruhe ist die Intensität der Laute nur gering (max. 84 dB SPL, 10 cm vor dem Maul der Fledermaus), im Flug steigt die Lautintensität auf bis zu 120 dB. Die Lautdauer variiert situationsabhängig zwischen 1 und 0,4 Millisekunden (ms). Im Flug werden die Laute während des Flügelaufschlags abgegeben[5][6][7][8].
Der Geruchssinn ist bei Desmodus gut entwickelt[9][10][11]. Die Riechleistung liegt für einige Duftstoffe im Bereich der olfaktorischen Sensitivität des Menschen[12]. Untersuchungen lassen darauf schließen, dass die Vampire ihren Geruchssinn insbesondere bei Kontakten in der sozialen Gruppe sowie möglicherweise auch beim Auffinden und Identifizieren einzelner Beutetiere nutzen[13].
Der Sehsinn ist bei Desmodus hervorragend ausgebildet, wie Untersuchungen zur Sehschärfe (abhängig von der Beleuchtungsstärke[14]), zur Größe des Gesichtsfeldes sowie zur Geschwindigkeits-Auflösung zeigen[15]. Desmodus ist darüber hinaus in der Lage, visuelle Muster zu unterscheiden[16]. Die Forschungsergebnisse lassen erwarten, dass Desmodus den visuellen Sinn beim Flug im Freiland sowohl über größere Distanz nutzt, um natürliche Landmarken (etwa Silhouetten von Bäumen oder Felsen) zu identifizieren, wie auch auf kürzere Distanz, zum Beispiel im Umfeld der Quartiere. Auf Distanzen unterhalb eines halben Meters scheinen sich diese Fledermäuse dann vor allem auf die Echoortung zu verlassen[17].
Spezifische Anpassungen an die blutfressende Ernährungsweise haben sich bei diesen Vampirfledermäusen insbesondere in zwei Sinnessystemen ausgebildet: zum einen beim Gehör (passive akustische Orientierung), zum anderen beim Tast- und Wärmesinn.
Das Gehör von Desmodus rotundus ist empfindlich vor allem im Ultraschallbereich zwischen 50 und 100 Kilohertz (dem Bereich ihrer eigenen Echoortung) sowie in einem Frequenzband zwischen 10 und 30 kHz. Wie elektrophysiologische Untersuchungen gezeigt haben, besitzt Desmodus im Vergleich zu vielen anderen Fledermäusen unterhalb von 25 kHz ungewöhnlich niedrige Hörschwellen, das heißt ein sehr gutes Hörvermögen[18]. Zudem fanden die Forscher im Gehirn der Vampirfledermaus Nervenzellen, die nur auf „Rauschen“ (breitbandige Schallfrequenzen) reagierten, sowie Neurone, die ausschließlich auf (menschliche) Atemgeräusche antworteten. Diese und weitere Forschungsergebnisse[19] lassen darauf schließen, dass Vampire das extrem leistungsfähige Gehör sehr gut nutzen können, um Beutetiere anhand der Geräusche des Atems oder kleinster Bewegungen aufzuspüren.

Bei der eigentlichen Auswahl der Bissstelle besitzt dagegen der Wärmesinn von Desmodus eine zentrale Bedeutung. Er ist im Vergleich zu anderen Fledermausarten außergewöhnlich leistungsfähig und ermöglicht den Tieren, die von durchblutetem Gewebe ausgehende Wärmestrahlung aus mehr als 10 Zentimeter Entfernung wahrzunehmen[20]. Weitergehende Untersuchungen[21] haben gezeigt, dass im zentralen Teil des Nasenaufsatzes (dem „Nasenblatt“) von Desmodus (siehe Abbildung) eine große Zahl an Kalt- und Warmrezeptoren (so genannte freie Nervenendigungen) konzentriert sind, insbesondere in dem dünnen Rand sowie dem mittleren Grat des Nasenblattes. Der Aktivitätsbereich der Warmrezeptoren ist im Vergleich mit anderen Säugetieren (z. B. Nagern) deutlich zu niedrigeren Temperaturen verschoben. Ähnliche Eigenschaften und Leistungen kannte man zuvor nur von bestimmten Schlangenarten[22]. Die zentrale Gesichtsregion von Desmodus bildet mithin ein regelrechtes Wärmesinnesorgan.
Desmodus trägt rund um dieses Nasenblatt zudem zahlreiche Sinneshaare, deren Mechanorezeptoren sehr empfindlich sowie richtungsabhängig auf winzigste Bewegungsreize antworten[23]. Der Tastsinn spielt offenbar eine wichtige Rolle während des eigentlichen Fressvorganges: Da die Fledermaus dabei direkten Körperkontakt mit der zumeist erheblich größeren Beute hat, ist sie durch mögliche Abwehrbewegungen gefährdet. Der sensible Tastsinn hilft den Tieren in solchen Situationen blitzschnell reagieren zu können.
Sozialverhalten
Gemeine Vampire besitzen ein hochentwickeltes Sozialverhalten. Dazu gehört die gegenseitige Fellpflege, ein unter Fledermäusen eher unübliches Verhalten; außerdem teilen sie sich mit ihren hungrigen Artgenossen häufig ihre Blutnahrung, indem sie diese hochwürgen und andere Tiere damit füttern[24].
Ein Gemeiner Vampir stirbt, wenn er in zwei oder drei aufeinanderfolgenden Nächten keine Nahrung zu sich nimmt. Zwischen 7 und 30 % der Tiere einer Gruppe scheitern jedoch in einer Nacht bei ihrer Nahrungssuche, sei es wegen Krankheit, Verletzung, Geburt oder simpler Erfolglosigkeit. Aufgrund des dringenden Blutbedarfes und der Schwierigkeiten, Beute zu finden, spielt das Heraufwürgen und Teilen der Nahrung eine wichtige Rolle für das Überleben der Gemeinen Vampire. Ohne Teilen der Nahrung läge die jährliche Todesrate der Tiere hochgerechnet bei 82 %, tatsächlich aber beträgt sie 24 %.
Wenn ein Tier eine nach menschlichen Maßstäben „selbstlose“ Handlung ausführt, geschieht dies entweder zum Nutzen seiner Verwandten, insbesondere der Nachkommen, oder zum Nutzen seiner selbst, damit ihm also sozusagen selbst ebenfalls entsprechend geholfen wird. Dies bezeichnet man als Reziproken Altruismus. Das Blutteilen der Gemeinen Vampire ist ein Beispiel dafür. Schätzungen zufolge gewinnt ein hungriges Tier durch eine Nahrungsspende 18 Stunden bis zum Hungertod, während der Spender nur rund 6 Stunden verliert. In Summe profitieren also beide Tiere von diesem Verhalten.
Abgesehen von der Versorgung des eigenen Nachwuchses, was im Tierreich absolut üblich ist, hängt die Frage, ob ein Vampir zum Teilen bereit ist, auch von der Wahrscheinlichkeit ab, diese Leistung auch einmal wieder zurückzubekommen, beziehungsweise von der Tatsache, ob das bittende Tier früher schon einmal mit dem jetzt Angebettelten geteilt hat. Voraussetzung dafür ist, dass die Tiere einander individuell erkennen. Die gegenseitige Fellpflege dürfte eine wichtige Rolle beim Identifizieren der einzelnen Artgenossen spielen. Wenn ein hungriges Tier einem anderen das Fell pflegt, so kann der angebettelte Artgenosse dieses identifizieren und erkennen, ob es selbst schon einmal geteilt hat. Mitentscheidend für den reziproken Altruismus scheint zu sein, dass Desmodus-Weibchen sehr stabile Gruppen bilden, in denen die einzelnen Individuen sehr lange, möglicherweise lebenslang zusammenbleiben[25].
Eine wichtige Rolle im Sozialleben der Vampire spielen Lautäußerungen. Bei entsprechenden Untersuchungen[26]. konnten insgesamt 16 deutlich unterscheidbare Laute identifiziert werden. Die meisten davon haben einen starken Frequenzanteil im Hörbereich des Menschen (also unterhalb von ca. 16 Kilohertz). Einzelne Lauttypen kann man bestimmten Situationen oder Funktionen im Rahmen des Sozialverhaltens der Tiere zuordnen: So gibt es Laute mit aggressivem Charakter, weiterhin Droh-, Kampf- und Abwehrlaute, außerdem Befreiungs-, Protest-, Kontakt- und Beschwichtigungslaute. Bestimmte Laute weisen strukturelle Ähnlichkeiten mit den so genannten Verlassenheitslauten von Jungtieren auf und haben sich möglicherweise direkt aus diesen entwickelt.
Beutetiere

Im Prinzip kann jedes Tier Opfer eines Vampirbisses werden. Der Gemeine Vampir nutzt als Wirte aber hauptsächlich Säugetiere, vornehmlich Rinder, Pferde und Esel sowie eine Reihe größerer Wildtiere wie etwa Tapire. Weniger häufig gehören auch Menschen zu ihren Blutquellen. Darüber hinaus werden auch größere Vögel wie Hühner und Truthühner gebissen. Hunde zählen kaum jemals zu den Opfern, vermutlich, weil sie die Ultraschalllaute, die die Tiere zur Orientierung aussenden, hören können.
Experimente mit gefangenen Tieren haben gezeigt, dass sie auch Schlangen, Echsen, Kröten, Krokodile und Schildkröten beißen. Ob diese Tiere auch in der Natur zu ihren Opfern zählen, ist nicht bekannt.
Annäherung an das Opfer
Die Tiere landen meistens nicht direkt auf ihren Opfern, sondern in der Nähe und laufen oder hüpfen dann in seine Richtung. Die kräftigen Hinterbeine und der lange Daumen stellen Anpassungen an diese Fortbewegungsweise dar.
Zunächst sucht sich die Fledermaus eine geeignete Bissstelle an ihrem Opfer, beispielsweise bei Rindern oder Pferden direkt oberhalb der Hufe. Die Bewegungen von Desmodus bei der Annäherung an eine Beute sind scheu und vorsichtig; die unter Fledermäusen seltene Fähigkeit zu hüpfen dient dem schnellen Ausweichen für den Fall, dass sie entdeckt und vom Opfer mit Tritten oder Schwanzschlägen verscheucht werden. Vampire kämpfen nicht mit ihren Beutetieren, ihr Biss erfolgt in der Regel unbemerkt und oft wacht das schlafende Tier nicht einmal auf. Als Bissstelle bevorzugen sie nicht von Haaren oder Federn bedeckte Körperteile wie die Anal- und die Vaginalregion oder die Ohren von Säugetieren beziehungsweise die Beine oder Kämme der Vögel.
Der Fressvorgang

Zunächst wird die Körperstelle mit der Zunge abgeleckt. Der Speichel der Tiere enthält ein Betäubungsmittel, mit dem die Bissstelle lokal unempfindlich wird. Anschließend werden eventuell vorhandene Haare oder Federn mit den Zähnen abrasiert. Mit den scharfen Schneidflächen der Eck- und Schneidezähne beißen sie ein Stück der Haut heraus. Die so entstehende Wunde ist rund drei bis zehn Millimeter breit und einen bis fünf Millimeter tief. Mit der Zunge lecken sie das ausfließende Blut auf und pumpen es durch die Rillen an der Unterseite der Zunge in den Mund. Ein Gerinnungshemmer sorgt dafür, dass das austretende Blut beim Trinkvorgang nicht gerinnt und somit für das Tier unbrauchbar wird. Das hierfür verantwortliche Enzym heißt Desmoteplase, auch Draculin oder Desmodus rotundus Salivary Plasminogen Activator (kurz DSPA) genannt; es ist ein Glykoprotein und bewirkt, dass sich das Fibrin (der feste Gerüststoff von gerinnendem Blut) auflöst. Nach einem Biss kann das Blut bis zu acht Stunden aus der Wunde fließen.
Der gesamte Vorgang kann bis zu zwei Stunden dauern, die eigentliche Nahrungsaufnahme bis zu 30 Minuten. Dabei nehmen die Tiere rund 20 bis 30 Milliliter Blut auf, eine Menge, die das Gewicht der Tiere nahezu verdoppelt. Dadurch fällt es ihnen häufig schwer, sich wieder in die Luft zu erheben. Nach einer Mahlzeit begeben sie sich an ihren Schlafplatz, um zu verdauen.
Gefahren des Bisses

Der Blutverlust des Opfers stellt ein geringes Problem dar. Viel größer sind die Gefahren durch Infektionskrankheiten, die von den Tieren übertragen werden, insbesondere die Tollwut und durch Trypanosomen verursachte Viehseuchen. Darüber hinaus kann es an der offenen Wunde zu Infektionen und zum Befall durch Larven der Neuwelt-Schraubenwurmfliege (Cochliomyia hominivorax) kommen. Tausende Tiere sterben so jährlich an den Folgen von Vampirbissen und auch Menschen kommen immer wieder zu Schaden.
Fortpflanzung
Das Reproduktionsverhalten von Desmodus unterscheidet sich in einigen Punkten von dem anderer, auch nah verwandter Fledermausarten[27].
Prinzipiell können sich Vampirfledermäuse das ganze Jahr über fortpflanzen. In den meisten Regionen ihres Verbreitungsgebietes treten aber ein- oder zweimal pro Jahr Geburtenspitzen auf. In Costa Rica fallen die meisten Geburten zum Beispiel in die Regenzeit (Oktober bis November), auf Trinidad und Tobago gibt es eine Spitze von April bis Mai, eine zweite von Oktober bis November.

Um die Paarung einzuleiten, klettert das Männchen auf den Rücken der Weibchen, umschließt ihre Flügel mit seinen Flugmembranen und beißt in ihren Nacken. Nach einer ca. siebenmonatigen Tragzeit[28] kommt in der Regel ein einzelnes Jungtier zur Welt. Neugeborene sind gut entwickelt und wiegen zwischen fünf und sieben Gramm. Im ersten Lebensmonat werden sie ausschließlich gesäugt, ab dem zweiten Lebensmonat füttert die Mutter sie zusätzlich mit hochgewürgtem Blut. Ab dem vierten Monat beginnt das Jungtier seine Mutter auf Beuteflügen zu begleiten; endgültig entwöhnt wird es aber erst mit neun oder zehn Monaten. Ungefähr im gleichen Alter tritt auch die Geschlechtsreife ein.
Werden die Jungtiere von ihrer Mutter getrennt, stoßen diese so genannte Verlassenheitslaute aus. Diese Laute bleiben erhalten, bis die Jungtiere nach ca. zehn Monaten endgültig unabhängig von der Mutter werden. Untersuchungen haben gezeigt, dass diese Laute individuell unterscheidbare Merkmale besitzen, die sich im Verlauf der Juvenilentwicklung nur unwesentlich verändern[29]. Dies ist eine entscheidende Voraussetzung dafür, dass die Muttertiere ihre eigenen Jungtiere anhand dieser Laute zweifelsfrei identifizieren können.
Bedingt durch die lange Tragzeit und die Abhängigkeit der Jungtiere von ihrer Mutter, können die Weibchen nur alle neun bis zehn Monate gebären, was den Fortpflanzungserfolg limitiert. Die Lebensdauer ist, wie bei vielen anderen Fledermäusen auch, recht lang. In freier Natur wird die Lebenserwartung auf neun bis zwölf Jahre geschätzt; das höchste bekannte Alter eines Tieres in menschlicher Obhut betrug 29 Jahre.
Natürliche Feinde
Zu den wenigen natürlichen Feinden der Gemeinen Vampire zählen Eulen, Greifvögel und Schlangen. Gegenüber letzteren nutzen sie ihre besonderen lokomotorischen Fähigkeiten, sich durch schnelle Sprünge in Sicherheit zu bringen.
Gemeine Vampire und Menschen
Die Sagengestalt Vampir
Legenden und Mythen von Vampiren, Sagengestalten, die sich von Blut ernähren und manchmal in Fledermaus- oder Eulengestalt auftreten, finden sich in vielen Kulturen rund um den Globus. Vielfach sind diese Sagen älter und können auch aufgrund der geografischen Distanz nicht von den Vampirfledermäusen beeinflusst sein, die nur aus Amerika belegt sind. Die Tiere haben ihren Namen von der Sagengestalt und nicht umgekehrt, wie auch die frühere Verwendung des Wortes „Vampir“ für die Gestalt als für die Tiere zeigt. Näheres dazu siehe unter dem Lemma Vampirfledermäuse.
Unabhängig davon gab es in der Mythologie der Maya ein Camazotz genanntes Ungeheuer in Fledermausgestalt, das Menschen und Tiere anfiel und ihr Blut trank. Inwieweit diese Vorstellungen vom Gemeinen Vampir oder von Desmodus draculae, einer ausgestorbenen, noch größeren Art der Vampirfledermäuse (siehe unter Systematik), beeinflusst sind, ist unklar.
Schäden durch Vampirbisse
Die Schäden, die durch den Gemeinen Vampir verursacht werden, sind enorm. Jährlich werden zahllose Nutz- und Haustiere das Opfer von Krankheiten, die durch Vampirbisse übertragen werden. Insbesondere Hausrinder sind gefährdet, da sie in großer Zahl vorhanden und auf den Weiden leicht zugänglich sind. Schätzungen zufolge sind diese Fledermäuse jedes Jahr für bis zu 100.000 Todesfälle bei Rindern durch Tollwut und durch Trypanosomen verursachte Krankheiten wie Surra verantwortlich. Eine Studie aus dem Jahr 1988 bezifferte den jährlichen wirtschaftlichen Schaden, der daraus entsteht, auf 40 Millionen US-Dollar.
Obwohl Menschen nicht zu den bevorzugten Opfern des Gemeinen Vampirs zählen, kommt es auch bei ihnen immer wieder zu Todesfällen durch Krankheiten, die durch die Bisse dieser Tiere hervorgerufen werden. So starben im Mai 2004 in Brasilien bis zu 22 Menschen an der Tollwut, die durch diese Tiere übertragen wurde.
Bedrohung durch den Menschen
Mit der großflächigen Einführung von Rinderfarmen und Viehweiden in Mittel- und Südamerika haben sich die Bestandszahlen des Gemeinen Vampirs vergrößert. Aufgrund der Gefahren durch Krankheitsübertragungen wird jedoch vielerorts versucht, die Tiere auszurotten. Höhlen wurden gesprengt oder ausgeräuchert. Da Gemeine Vampire optisch schlecht von anderen Fledermausarten zu unterscheiden sind, waren auch viele harmlose frucht- oder insektenfressende Tiere betroffen, oft im stärkeren Ausmaß als Gemeine Vampire selbst. Andere Methoden, die gegen diese Fledermäuse eingesetzt werden, sind langsam wirkende Gifte, die auf das Fell gefangener Tiere gestrichen werden und die durch die gegenseitige Fellpflege auch die anderen Bewohner des Quartiers töten, schwache Gifte, die in die Rinder injiziert werden, Fangnetze vor Ställen und andere. Trotz alledem ist die Art häufig und zählt nicht zu den bedrohten Arten.
Bedeutung für die Forschung
Es war schon länger bekannt, dass ein Bestandteil des Speichels dieser Tiere die Blutgerinnung hemmt. Seit den 1980er Jahren gab es Bestrebungen, dieses Enzym zu isolieren, was einem deutsch-mexikanischen Forscherteam gelang. Seit Anfang der 1990er Jahre kann dieses Enzym (Desmodus rotundus Salivary Plasminogen Activator (DSPA) oder Desmoteplase genannt) biotechnologisch hergestellt werden. Es sollte vor allem als Medikament gegen ischämische Schlaganfälle eingesetzt werden, da es auch bereits bestehende Blutverklumpungen im Gefäßsystem auflösen kann. Zwischen 1990 und 1999 wurde schon eine Studie mit Herzinfarktpatienten erfolgreich abgeschlossen. Synthetisierte Präparate, die das Enzym enthalten, unterdrücken die Blutgerinnung bis zu 20-mal länger als herkömmliche Mittel und sie sollen nach ersten Studien auch eine bis zu 150-mal stärkere Wirkung aufweisen. Man versprach sich mit diesem Mittel eine Erweiterung des Zeitfensters für die Behandlung, das gegenwärtig beim Ischämischen Hirninfarkt bei drei Stunden liegt. In der Phase-III-Studie konnte beim Ischämischen Hirninfarkt jedoch keine Verbesserung des klinischen Outcomes nachgewiesen werden.[30]
Systematik
Stellung innerhalb der Fledermaussystematik
Der Gemeine Vampir bildet zusammen mit dem Kammzahnvampir (Diphylla ecaudata) und dem Weißflügelvampir (Diaemus youngi) die Gruppe der Vampirfledermäuse (Desmodontinae), wobei der Weißflügelvampir der nächste Verwandte des Gemeinen Vampirs ist. Vampirfledermäuse werden als Unterfamilie der Blattnasen (Phyllostomidae) eingeordnet, einer formenreichen, auf den amerikanischen Kontinent beschränkten Fledermausgruppe. Phylogenetisch bilden sie das Schwestertaxon aller übrigen Blattnasenarten.
Von der Gattung Desmodus sind außerdem zwei ausgestorbene Arten bekannt:
- Desmodus stocki war etwas schwerer als der Gemeine Vampir und lebte in den südlichen Vereinigten Staaten und in Mexiko. Die Art ist erst vor rund 3000 Jahren ausgestorben.
- Desmodus draculae, auch „Riesenvampir“ genannt, war größer als der Gemeine Vampir, seine Flügelspannweite wird auf 60 bis 75 Zentimeter geschätzt. Fossile Überreste dieser Art aus dem Pleistozän wurden in Mexiko, Venezuela, Argentinien und Brasilien gefunden.[31] Wegen des (zumindest behaupteten) jungen Alters der Funde und Legenden der Maya über Camazotz, eines Wesens in Gestalt eines Riesenvampirs, mutmaßen Kryptozoologen, dass die Art sogar noch leben könnte oder erst in jüngster Zeit ausgestorben ist. Beweise dafür gibt es allerdings keine.
Stammesgeschichte
Abgesehen von den zwei oben erwähnten Arten sind keine fossilen Vorfahren der Vampirfledermäuse bekannt. Die Frage, wie es zur Entwicklung dieser spezialisierten Ernährungsweise kam, kann derzeit nur auf spekulativem Weg beantwortet werden. Zwei Theorien wurden hierfür vorgeschlagen:
- Nach einer Theorie haben sich Vampirfledermäuse aus fruchtfressenden Vorfahren entwickelt, die besonders geformten Schneide- und Eckzähne seien zunächst eine Anpassung an das Aufbeißen hartschaliger Früchte gewesen.
- Nach einer anderen Theorie haben sich Vampirfledermäuse aus insektenfressenden Vorfahren entwickelt, die auf ektoparasitische (auf der Haut lebende) Tiere spezialisiert waren. Möglicherweise lockten die Wunden, die die Insekten ihren Wirten zufügten, diese Vorfahren an.
Innerhalb der Blattnasen gibt es sowohl frucht- als auch insektenfressende Arten, so dass beide Wege denkbar sind.
Literatur
- Klaus Richarz: Fledermäuse beobachten, erkennen und schützen. Kosmos, Stuttgart 2004, ISBN 3-440-09691-2.
- Ronald M. Nowak: Walker's Mammals of the World. Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9.
- Uwe Schmidt: Vampirfledermäuse. Spektrum Verlag, Heidelberg 1995. ISBN 978-3-89432-176-5.
- Dennis Turner: The Vampire Bat. A Field Study in Behavior and Ecology. Johns Hopkins University Press, Baltimore 1975, ISBN 0-8018-1680-7.
- Arthur M. Greenhall, Gerhard Joermann, Uwe Schmidt: Desmodus rotundus (PDF; 753 kB). In: Mammalian Species. 202, 1983.
- Arthur M. Greenhall, Uwe Schmidt: Natural History of Vampire Bats. CRC Press, Boca Raton 1988, ISBN 0-8493-6750-6.
- Gerald S. Wilkinson: Food Sharing in Vampire Bats. (PDF; 919 kB) In: Scientific American. 262, 1990, S. 76–82.
Einzelnachweise
- J. S. Altenbach: Locomotion. In: Natural History of Vampire Bats. (Editors: A. M. Greenhall, U. Schmidt). CRC Press, Boca Raton 1988, S. 71.
- J. S. Altenbach: The locomotiv morphology of the vampire bat, Desmodus rotundus. In: Spec. Publ. Am. Soc. Mamm. 6, 1979, S. 1.
- D. K. Riskin, J. W. Hermanson: Biomechanics: Independent evolution of running in vampire bats. In: Nature, Band 434, 2005, S. 292.
- "Gemeiner Vampir" zootierliste.de
- U. Schmidt, C. Schmidt: Echolocation performance of the vampire bat (Desmodus rotundus). In: Zeitschrift fu?r Tierpsychologie. Band 45, Nummer 4, Dezember 1977, S. 349–358, PMID 610226.
- U. Schmidt, G. Joermann: Echoortung bei der Vampirfledermaus Desmodus rotundus. In: Z. f. Säugetierkunde. Band 46, 1981, S. 129.
- G. Joermann, U. Schmidt: Echoortung bei der Vampirfledermaus Desmodus rotundus. II. Lautaussendung im Flug und Korrelation zum Flügelschlag. In: Z. f. Säugetierkunde. Band 46, 1981, S. 136.
- G. Joermann: Echoortung bei Vampirfledermäusen (Desmodus rotundus) im Freiland. In: Z. f. Säugetierkunde. 49, 1984, S. 221.
- H. Stephan, P. Pirlot: Volumetric comparison of brain structures in bats. In: Bijdragen tot de Dierkunde. 40, 1970, S. 95.
- K. P. Bhatnagar, F. C. Kallen: Cribriform plate of ethmoid, olfactory bulb and olfactory acuity in forty species of bats. In: J. Morphol. 142, 1974, S. 71.
- R. Kämper, U. Schmidt: Die Morphologie der Nasenhöhle bei einigen neotropischen Chiropteren. In: Zoomorphol. Band 87, 1977, S. 3.
- U. Schmidt: Vergleichende Riechschwellenbestimmungen bei neotropischen Chiropteren. In: Z. f. Säugetierkunde. Band 40, 1975, S. 269.
- U. Schmidt: Orientation and Sensory Functions in Desmodus rotundus. In: Natural History of Vampire Bats (Editors: A. M. Greenhall, U. Schmidt). CRC Press, Boca Raton 1988, S. 143.
- U. Manske, U. Schmidt: Visual acuity of the vampire bat, Desmodus rotundus, and its dependence upon light intensity. In: Zeitschrift fu?r Tierpsychologie. Band 42, Nummer 2, Oktober 1976, S. 215–221, PMID 1007656.
- K. Hessel: Vergleichende optokinetische Untersuchungen zur Leistungsfähigkeit des visuellen Systems bei neotropischen Fledermäusen (Fam. Phyllostomidae) und der Hausmaus. Diss., Bonn 1998.
- U. Manske, U. Schmidt: Untersuchungen zur optischen Musterunterscheidung bei der Vampirfledermaus, Desmodus rotundus. In: Z. Tierpsychol. Band 49, 1979, S. 120.
- G. Joermann, U. Schmidt, C. Schmidt: The Mode of Orientation during Flight and Approach to Landing in two Phyllostomid Bats. In: Ethology. Band 78, 1988, S. 332.
- U. Schmidt, P. Schlegel, H. Schweizer, G. Neuweiler: Audition in vampire bats, Desmodus rotundus (PDF; 733 kB). In: J. Comp. Physiol. A. Band 168, 1991, S. 45.
- U. Gröger, L. Wiegrebe: Classification of human breathing sounds by the common vampire bat, Desmodus rotundus. In: BMC Biology. 4, 2006, S. 18.
- L. Kürten, U. Schmidt: Thermoperception in the common vampire bat Desmodus rotundus. In: J. Comp. Physiol. Band 146, 1982, S. 223.
- L. Kürten, U. Schmidt, K. Schäfer: Warm and Cold Receptors in the Nose of the Vampire Bat, Desmodus rotundus. In: Naturwissenschaften. Band 71, 1984, S. 327.
- A. L. Campbell, R. R. Naik, L. Sowards, M. O. Stone: Biological infrared imaging and sensing (Memento des Originals vom 15. Juni 2003 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis. (PDF; 628 kB). In: Micron. Band 33, 2002, S. 211.
- L. Kürten: Mechanorezeptoren im Nasenaufsatz der Vampirfledermaus Desmodus rotundus. In: Z. f. Säugetierkunde. Band 50, 1985, S. 26.
- G. S. Wilkinson: Reciprocal food sharing in the vampire bat. In: Nature. Band 308, 1984, S. 181.
- G. S. Wilkinson: Social Organization and Behavior. In: Natural History of Vampire Bats (Editors: A. M. Greenhall, U. Schmidt). CRC Press, Boca Raton 1988, S. 85.
- S.-R. Park: Sozialverhalten und akustische Kommunikation bei der Vampirfledermaus, Desmodus rotundus. Diss. Bonn, 1986.
- C. Schmidt: Reproduction. In: Natural History of Vampire Bats (Editors: A. M. Greenhall, U. Schmidt). CRC Press, Boca Raton 1988, S. 99.
- U. Schmidt: Die Tragzeit der Vampirfledermäuse (Desmodus rotundus). In: Z. f. Säugetierkunde. Band 39, 1974, S. 129.
- U. Schmidt, G. Joermann, C. Schmidt: Struktur und Variabilität der Verlassenheitslaute juveniler Vampirfledermäuse (Desmodus rotundus). In: Z. f. Säugetierkunde. Band 47, 1982, S. 143.
- T. Steiner, E. Jüttler, P. Ringleb: Akuttherapie des Schlaganfalls. In: Der Nervenarzt. Band 78, Nr. 10, 2007, S. 1147–1154.
- Santiago Brizuela & Daniel A. Tassara. 2021. New Record of the Vampire Desmodus draculae (Chiroptera) from the Late Pleistocene of Argentina. Ameghiniana 58 (2): 169-176; doi: 10.5710/AMGH.30.12.2020.3379
Weblinks
- Desmodus rotundus in der Roten Liste gefährdeter Arten der IUCN 2009. Eingestellt von: R. Barquez, S. Perez, B. Miller, M. Diaz, 2008. Abgerufen am 18. Oktober 2009.
- Bildergalerie (englisch)
- Informationen des Pharmakonzerns Paion zu Desmoteplase