Kiemenlochtiere
Die Kiemenlochtiere (Hemichordata) sind ein Tierstamm, zu dem nur gut 100 bekannte rezente Arten gehören. Sie umfassen zwei Gruppen, die in der klassischen Systematik im Rang von Klassen geführt werden – die wurmartigen Eichelwürmer (Enteropneusta) und die koloniebildenden, festsitzenden (sessilen) Flügelkiemer (Pterobranchia).
| Kiemenlochtiere | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
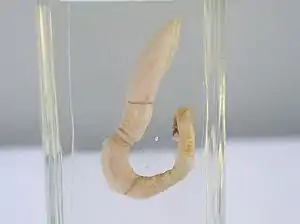
Eichelwurm (Saccoglossus sp.) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Hemichordata | ||||||||||||
| Bateson, 1885 | ||||||||||||
| Klassen | ||||||||||||
|
Die Kiemenlochtiere zählen zum Überstamm der Neumünder (Deuterostomia), zu denen auch die Chordatiere (Chordata) und mit diesen die Wirbeltiere (Vertebrata) gehören. Der wissenschaftliche Name Hemichordata („halbe Chordatiere“) spiegelt die traditionelle Ansicht wider, sie seien die Urform der Chordata. Tatsächlich stehen die Hemichordata mit den Stachelhäutern (Echinodermata) – Seesterne (Asteroidea), Seeigel (Echinoidea) und andere – in einem Schwestergruppenverhältnis und bilden mit ihnen ein Taxon, das Ambulacraria genannt wird.
Anatomie
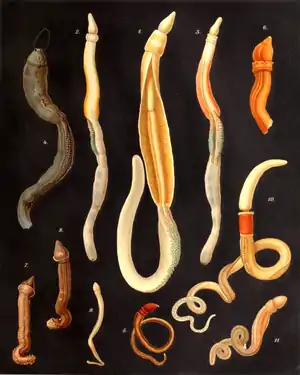
Kiemenlochtiere haben einen weichen, wurmähnlichen, aber (von einer groben Dreiteilung abgesehen) innerlich unsegmentierten Körper, der wie bei allen zweiseitig symmetrisch gebauten Tieren eine Spiegelsymmetrie aufweist und somit über drei klar definierte Körperachsen verfügt. Sie kommen in verschiedenen Farben von weiß bis dunkelviolett vor und können bis zu 2,50 m lang werden, andererseits finden sich aber auch nur millimetergroße Tiere. Charakteristisch ist die Gliederung des Körpers in drei Teile: Der vorderste Abschnitt ist der Kopflappen (Prosoma), der in der Klasse der Flügelkiemer (Pterobranchia) schildförmig geformt ist und daher auch als Kopf- oder Rostralschild bezeichnet wird. Darauf folgt ein kurzer Kragen (Mesosoma), in dem die Mundöffnung untergebracht ist und ein langer Rumpf (Metasoma). Diese grundlegende Dreiteilung des Körpers betrifft auch die Leibeshöhle, das Coelom, welche in Pro-, Meso- und Metacoel gegliedert ist, die räumlich in den entsprechenden Körperabschnitten liegen.
Insbesondere in der Klasse der Eichelwürmer (Enteropneusta) befinden sich im Rumpfabschnitt bis zu hundert für die Gruppe namensgebende Kiemenspalten, durch welche die Tiere atmen und an denen das durch den Mund eingesogene Wasser unter Rückhaltung der Nahrungspartikel wieder ausströmen kann. Sie verbinden den ersten Darmabschnitt, den Kiemendarm, mit der Außenwelt. Bei den Flügelkiemern dagegen sind entweder nur zwei paarig angelegte Kiemen vorhanden oder diese fehlen ganz.
Als Ausstülpung des Magens in den Mundlappen (Buccaltasche) existiert bei den Eichelwürmern eine weitere charakteristische Struktur, nämlich das Stomochord. Früher galt es als möglicher Vorläufer des Notochords, also der elastischen, stabförmigen Stützstruktur der Chordatiere. Da es mit diesem allerdings keine wesentlichen Gemeinsamkeiten aufweist und auch zu keinem Zeitpunkt als hydrostatisches Skelett dient, wird diese Ansicht heute wissenschaftlich kaum mehr vertreten.
Das offene Blutgefäßsystem der Kiemenlochtiere ist recht primitiv ausgebildet, nur im Kragen und Kopflappen befinden sich zwei echte Blutkanäle. Im Kopflappen liegt auch das sehr elementare „Herz“, das lediglich aus einem einzigen zusammenziehbaren (kontraktilen) Blutgefäß besteht, das rückseitig (dorsal) ankommendes Blut in die bauchseitige (ventrale) Ader pumpt. Der Blutfluss ist also gerichtet (unidirektional).
Die Ausscheidung flüssiger Abfallstoffe geschieht hauptsächlich durch die Haut, darüber hinaus existiert ein Glomerulus genanntes Membransystem, in dem aus dem Herz einströmendes Blut mehrfach gefiltert wird. Der entstehende Urin wird in die Leibeshöhle des Kopflappens entlassen und gelangt von dort durch eine Pore nach außen.
Das Nervensystem besteht im Wesentlichen aus je einem bauchseitigen und rückseitigen Nervenstrang, die im Kopflappen und um den Darm herum ringförmig verbunden sind und Nervenenden in die Außenhaut (Epidermis) entsenden. Bei den Eichelwürmern verläuft der rückseitige Nerv in einer speziellen Falte im Kragen. Dieser hohle Nervenstrang, das „Kragenmark“ der Eichelwürmer, wird vor allem wegen seiner embryonalen Entstehung manchmal als Homologon des Rückenmarks der Chordatiere angesehen, beide wären also stammesgeschichtlich aus derselben Vorgängerstruktur hervorgegangen.
Ernährung, Lebensraum und Verbreitung
Kiemenlochtiere können sich auf zwei verschiedenen Wegen ernähren: Entweder graben sie sich durch das Sediment des Meeresbodens, das heißt, sie nehmen Bodenschlamm auf und verdauen den darin enthaltenen organischen Inhalt, in etwa wie ein Regenwurm, oder sie filtrieren frei im Wasser schwebende Nahrungsteilchen wie zum Beispiel Algen. Sie leben daher meist in oder unterhalb der Gezeitenzone, auf oder im Grund des Meeres (benthisch), zum Teil bis in Tiefen von 5000 Metern, und bilden dort oft U-förmige Wohnhöhlen. Nur wenige Arten leben im offenen Meer (pelagisch). Kiemenlochtiere sind weltweit in allen Meeresgewässern von den Tropen bis in polare Zonen verbreitet.
Fortpflanzung
Kiemenlochtiere haben getrennte Geschlechter, die sich allerdings äußerlich kaum unterscheiden. Aus den befruchteten Eiern entwickeln sich meist erst bewimperte Larven („Tornaria-Larven“), die den Larven der Stachelhäuter ähneln und aus denen sich die erwachsenen Tiere entwickeln. Sie verbringen einen Teil ihres Lebenszyklus vor der Metamorphose im Plankton, wo sie sich von feinen Nahrungspartikeln ernähren, die in Wimpernbändern der Larve hängenbleiben und von dort zum Mund transportiert werden; man bezeichnet sie daher auch als planktotroph. Bei manchen Arten geschieht die Entwicklung allerdings auch direkt, das heißt ohne ein dazwischengeschaltetes Larvenstadium.
Neben der geschlechtlichen Fortpflanzung findet man auch die ungeschlechtliche: Dabei schnürt sich ein Jungtier in einem Knospung genannten Vorgang einfach vom genetisch identischen Elterntier ab. Daneben kann es auch vorkommen, dass ein Individuum einfach in zwei Teile zerfällt, die dann unabhängig voneinander fortbestehen.
Fossilien
Die ersten fossilen Funde von Kiemenlochtieren entstammen der chinesischen Chengjiang-Faunengemeinschaft und dem etwas jüngeren kanadischen Burgess-Schiefer, die im erdgeschichtlichen Zeitalter des Kambrium entstanden sind. Eine heute ausgestorbene Gruppe von Kiemenlochtieren, die Graptolithen (Graptolithina), bilden sogar wichtige Leitfossilien für Ordovizium und Silur.
Systematik
Man unterscheidet zwei Klassen noch heute lebender Kiemenlochtiere.
- Die Eichelwürmer (Enteropneusta) umfassen ungefähr 90 bis 100 Arten in vier Familien. Sie leben als Einzeltiere und sind im Gegensatz zu den Flügelkiemern nicht mit Tentakeln ausgestattet. Ihre Größe variiert zwischen 2,5 und 250 Zentimetern.
- Die etwa zwanzig Arten der Flügelkiemer (Pterobranchia) haben einen eher vasenförmigen Körper, verfügen über Tentakel und werden höchstens einen Zentimeter lang. Anders als die Eichelwürmer leben sie in Kolonien, in denen die Einzeltiere (Zooide) durch röhrenförmige Ausläufer, nämlich Stolons, miteinander verbunden sind. Eine solche Kolonie ist oft von einem aus Kollagen bestehenden Netzwerk von Höhlungen umgeben.
Unsicher und rätselhaft ist die Zuordnung der Art Planctosphaera pelagica, für die manchmal eine eigene Klasse Planctosphaeroidea unterschieden wird. Von dieser Art ist allerdings nur die charakteristische rundliche Tornaria-Larve bekannt, die im freien Ozean lebt und bis zu einem Zentimeter groß werden kann. Die meisten Forscher nehmen an, dass sie zu einer bisher unbekannten Eichelwurm-Art gehört, die vermutlich in der Tiefsee lebt.
Die Graptolithen (Graptolithina) bilden darüber hinaus eine wichtige Gruppe ausgestorbener Kiemenlochtiere; ihre Fossilien sehen wie spiralig aufgewundene kleine Sägeblätter aus. Es wird vermutet, dass dies eine Anpassung an das Schweben im freien Ozeanwasser (ihre pelagische Lebensweise) darstellt. Nach der Entdeckung der Flügelkiemer-Art Cephalodiscus graptolitoides vor der Küste Neukaledoniens, die in ihrem Körperbau verblüffende Übereinstimmung mit den fossilen Graptolithen zeigt, nehmen die meisten Forscher an, dass auch die Graptolithen zu den Flügelkiemern gehört haben.
Stammesgeschichte
Nach einigen Zweifeln, die Monophylie der Kiemenlochtiere und ihrer Teilgruppen betreffend, gilt heute die Monophylie aller drei Taxa nach morphologischen[1] und genetischen[2] Daten als wahrscheinlichste Hypothese, wenn auch die genetischen Daten gewisse Zweifel offenlassen, ob nicht die Flügelkiemer innerhalb der Eichelwürmer positioniert werden müssten[3] (was diese in diesem Fall paraphyletisch machen würde).
Das folgende Kladogramm gibt die nach heutigem Wissen wahrscheinlichsten Verwandtschaftsverhältnisse wieder:[4][5]
| Neumünder (Deuterostomia) |
| ||||||||||||||||||||||||||||||
Literatur
- D. T. Anderson: Invertebrate Zoology. Kap. 17. Oxford Univ. Press, Oxford 2001 (2. Aufl.), S. 418, ISBN 0-19-551368-1
- Richard S. K. Barnes, P. Calow, P. J. W. Olive, D. W. Golding, J. I. Spicer: The invertebrates - a synthesis. Kap. 7.2. 3. Aufl., Blackwell, Oxford 2001, S. 147, ISBN 0-632-04761-5
- Richard C. Brusca, G. J. Brusca: Invertebrates. Kap. 23. Sinauer, Sunderland Mass 2003 (2. Aufl.), S. 847, ISBN 0-87893-097-3
- A. Goldschmid: Hemichordata (Branchiotremata). in: Wilfried Westheide: Spezielle Zoologie. Teil 1. Einzeller und Wirbellose Tiere. Gustav Fischer, Stuttgart-Jena 1996, Akademie Verlag, Heidelberg 2004, ISBN 3-8274-1482-2
- E. E. Ruppert, R. S. Fox, R. P. Barnes: Invertebrate Zoology - A functional evolutionary approach. Kap. 27. Brooks/Cole 2004, S. 857, ISBN 0-03-025982-7
- J. van der Land: Hemichordata. in: M. J. Costello u. a. (Hrsg.): European register of marine species - a check-list of the marine species in Europe and a bibliography of guides to their identification. Collection Patrimoines Naturels. Bd 50. Publication scientifiques du M.N.H.N., Paris 2001, S. 336f, ISBN 2-85653-538-0
Einzelnachweise
- C.B. Cameron (2005): A phylogeny of the hemichordates based on morphological characters. Canadian Journal of Zoology 83: 196–215. doi:10.1139/Z04-190
- Oleg Simakov et al. (2015): Hemichordate genomes and deuterostome origins. Nature 527: 459-465. doi:10.1038/nature16150
- Johanna T. Cannon, Amanda L. Rychel, Heather Eccleston, Kenneth M. Halanych, Billie J. Swalla (2009): Molecular phylogeny of hemichordata, with updated status of deep-sea enteropneusts. Molecular Phylogenetics and Evolution 52: 17–24. doi:10.1016/j.ympev.2009.03.027
- Casey W. Dunn, Gonzalo Giribet, Gregory D. Edgecombe, Andreas Hejnol (2014): Animal Phylogeny and Its Evolutionary Implications. Annual Review of Ecology, Evolution, and Systematics 45: 371–395. doi:10.1146/annurev-ecolsys-120213-091627
- Maximilian J. Telford, Graham E. Budd, Hervé Philippe (2015): Phylogenomic Insights into Animal Evolution. Current Biology 25: R876–R887. doi:10.1016/j.cub.2015.07.060