Ceratopsidae
Die Ceratopsidae (auch Ceratopidae) sind eine Gruppe von Dinosauriern innerhalb der Ceratopsia.
| Ceratopsidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Lebendrekonstruktion von Triceratops | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Campanium bis Maastrichtium)[1] | ||||||||||||
| 83,6 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Ceratopsidae | ||||||||||||
| Marsh, 1890 |
Es waren große, quadrupede (sich auf allen vieren fortbewegende) Tiere, die durch ihre Hörner auf der Nase und über den Augen sowie durch ihren Nackenschild charakterisiert waren. Diese dienten nach heutigem Kenntnisstand vermutlich der Identifikation und Auseinandersetzung mit Artgenossen und weniger der Feindabwehr. Mit ihren Zahnbatterien (reihenförmig angeordnete Zähne, die bei Abnutzung durch den nachfolgenden Zahn ersetzt wurden) waren diese Dinosaurier gut an eine pflanzliche Ernährung angepasst. Die Ceratopsidae sind fast nur aus der späten Oberkreide (vor rund 83 bis 66 Millionen Jahren) aus dem westlichen Nordamerika bekannt, wo sie in erdgeschichtlich kurzer Zeit eine große Artenvielfalt erreichten. Die einzige bekannte Ausnahme stellt der erst 2010 beschriebene Sinoceratops dar, der im heutigen China gefunden wurde.[2]
Beim Massenaussterben am Ende der Kreidezeit sind sie, wie alle Nichtvogel-Dinosaurier, ausgestorben. Es werden zwei Unterfamilien unterschieden, die Centrosaurinae und die Chasmosaurinae.
Merkmale
Allgemeines
Die Ceratopsidae waren die größten Vertreter der Ceratopsia. Sie erreichten Längen von vier bis acht Metern und ein Gewicht von mehreren Tonnen. Es waren stämmig gebaute Tiere mit kräftigen Gliedmaßen, wobei die Hinterbeine deutlich länger als die Vorderbeine waren.
Zwar sind einige Gattungen nur durch Einzelfunde bekannt, von vielen Taxa gibt es jedoch zahlreiche Funde des Schädels und des Rumpfskeletts. Fossile Überreste des Rumpfskeletts sind seltener, von einigen Gattungen wie Arrhinoceratops und Diceratus ist nur der Schädel bekannt.
Schädel

Der Schädel der Ceratopsidae war wie bei allen Neoceratopsia sehr groß und wuchtig. Einschließlich des Nackenschildes erreichte er bei Pentaceratops und Torosaurus über zwei Meter, was den längsten bekannten Schädel aller landbewohnenden Tiere darstellt.[3] Von oben betrachtet war er annähernd dreieckig, wobei die zugespitzte Schnauze und die weit ausladende Wangenregion formgebend waren. Die Schnauzenspitze war wie bei allen Ceratopsia aus dem Rostralknochen (vor dem Oberkiefer) und dem Praedentale (vor dem Unterkiefer) gebildet. Die runzelige Oberfläche des Rostralknochens könnte darauf hinweisen, dass er von einer Keratinschicht umgeben gewesen sein könnte. Bei den Centrosaurinae war der Rostralknochen lateral (seitlich betrachtet) annähernd dreieckig, bei den Chasmosaurinae hingegen war er deutlich langgestreckter und nach unten gebogen. Das Praedentale war vorne nach oben gebogen und endete in einem spitzen Punkt, der am Rostralknochen anliegt. Die Schneidekante des Praedentales war bei den Chasmosaurinae flach und bei den Centrosaurinae seitlich gebogen.


Das hinter dem Rostralknochen gelegene Zwischenkieferbein (Praemaxillare) war sehr hoch und eines der prägendsten Elemente des Gesichtsschädels. Die Nasenlöcher waren deutlich vergrößert und annähernd rund. Bei den Chasmosaurinae befand sich im Zwischenkieferbein ein vermutlich luftgefüllter Hohlraum, der bei den Centrosaurinae fehlte. Das Tränenbein (Lacrimale) war bei den Ceratopsidae verkleinert, ebenso die Fenestra antorbitalis (das Schädelfenster vor der Augenhöhle). Das Nasenbein trug einen knöchernen Höcker, der oft zu einem Nasenhorn verlängert war und vermutlich mit Keratin überzogen war. Bei vielen Centrosaurinae war das Nasenhorn lang, es erreichte bis zu 50 Zentimeter bei Styracosaurus. Bei Einiosaurus war es auffällig nach vorne gebogen, bei manchen Centrosaurinae wie Achelousaurus und Pachyrhinosaurus war es zu einem verdickten Höcker umgebildet. Die Nasenhörner der Chasmosaurinae sind einheitlich kurz.
Die Überaugenhörner waren Auswüchse des Postorbitale, das auch den hinteren Rand der Augenhöhle und einen Teil der Wange bildete. Bei den Centrosaurinae waren die Überaugenhörner klein und erreichten maximal 15 Zentimeter Länge, manchmal waren sie zu Einbuchtungen oder kleinen Knochenhöckern umgebildet. Bei den Chasmosaurinae waren sie außer bei Chasmosaurus gut entwickelt und konnten bei den größeren Vertretern über einen Meter lang werden.
Die ausladende Wangenregion war stark modifiziert. Jugale, Quadratojugale und Quadratum sind ineinandergeschoben, die Fenestra infratemporalis (das untere Schädelfenster der Schläfenregion) war nach unten verschoben und stark verkleinert. Ein weiterer Knochen, das Epijugale, bildete bei manchen Arten wie Pentaceratops auffällige Wangenhörner, bei anderen ist es unauffällig. Die Fenestra supratemporalis (das obere Schädelfenster der Schläfengegend) wurde aus dem Postorbitale, dem Scheitelbein (Parietale) und dem Schuppenbein (Squamosum) gebildet. Ein nur bei den Ceratopsidae vorhandenes Merkmal ist ein hinter der Schädelhöhle gelegener Hohlraum, der sich durch eine Fontanelle auf den Hinterkopf öffnet. Dieser Hohlraum fehlt bei Jungtieren und ist bei ausgewachsenen Tieren sehr variabel. Die Funktion dieses Hohlraums ist bis heute ungeklärt.
Das neben den Hörnern auffälligste Merkmal der Ceratopsidae ist der Nackenschild, der aus dem Scheitel- und dem Schuppenbein gebildet wurde. Seine Länge betrug rund 60 bis 100 % der Länge des eigentlichen Schädels. Generell sind die Nackenschilde der Centrosaurinae kurz und die der Chasmosaurinae mit Ausnahme von Triceratops lang. Bei nahezu allen Arten fanden sich paarige Fenster im Schild, am größten waren sie bei Chasmosaurus und Pentaceratops, bei Triceratops waren sie hingegen geschlossen. Der äußere Rand des Schilds war mit wellenförmigen Verknöcherungen bedeckt, die Epoccipitalia genannt werden. Bei manchen Arten, insbesondere bei den Centrosaurinae, entwickelten sich die Epoccipitalia zu stachelähnlichen Strukturen, die auch nach vorne gerichtet sein konnten.
Zähne

Die Bezahnung der Ceratopsidae bestand aus Zahnbatterien, das sind reihenförmig angeordnete Zähne, die bei Abnutzung durch den nachfolgenden Zahn ersetzt wurden. Diese Form hat sich konvergent zu der der Hadrosauridae entwickelt. Die einzelnen Zähne standen in dicht gepackten Zahnreihen, jede Zahnposition verfügte über drei bis fünf Ersatzzähne. Die Anzahl der Zahnpositionen korreliert ungefähr mit der Schädelgröße: 28 bis 31 bei Centrosaurus und 36 bis 40 bei Triceratops. Die Okklusionsflächen des Gebisses standen annähernd senkrecht, was dafür spricht, dass die Zähne vorwiegend für eine schneidende Tätigkeit eingesetzt wurden.
Rumpfskelett
Die ersten Halswirbel der Ceratopsidae waren wie bei allen Coronosauria zum Syncervical verschmolzen, wobei umstritten ist, ob dieses aus den ersten drei oder vier Halswirbeln gebildet wurde. Die dahinterliegenden sechs Halswirbel waren kurz und breit. Danach befanden sich 12 freie Rückenwirbel, bis zu drei weitere Rückenwirbel waren mit den vier Kreuzbeinwirbeln und den ersten drei Schwanzwirbeln zum Kreuzbein (Sacrum) verwachsen. Der Schwanz der Ceratopsidae war lang, die genaue Anzahl der Schwanzwirbel ist aber nur in wenigen Fällen bekannt. So waren es bei Anchiceratops 38 oder 39, bei Styracosaurus hingegen 46, bei Pentaceratops sind 28 erhalten, wobei der letzte so klein ist, dass kaum mehr als fünf weitere vorhanden sein konnten. Der Schwanz wurde durch Chevronknochen (Y-förmige Fortsätze an der Unterseite der Schwanzwirbel) versteift.

Die Gliedmaßen der Ceratopsidae waren robust gebaut, die Vordergliedmaßen waren wie bei fast allen Dinosauriern kürzer als die Hintergliedmaßen und erreichten nur rund 70 % von deren Länge. Bei den größeren Arten wiesen die Röhrenknochen stark vergrößerte Enden mit rauer Oberfläche auf und ähnelten so denen der Sauropoden. Dies könnte ein Anzeichen für dickes Knorpelgewebe darstellen. Bei den kleineren Arten sind die Enden der Röhrenknochen weniger ausgeprägt und die Oberfläche ist glatt.[4]
Im Schultergürtel der Ceratopsidae ist das relativ große Rabenbein (Coracoid) auffällig, das bei ausgewachsenen Tieren mit dem Schulterblatt verschmolz. Schlüsselbeine wurden nie gefunden. Der Oberarmknochen war länger als die Speiche. Der Vorderfuß war immer kleiner als der Hinterfuß und kurz und breit gebaut. Er endete in fünf Zehen, von denen die ersten drei in stumpfen Hufen und die letzten beiden in knöchernen Noppen endeten.
Im Becken der Ceratopsidae war das Darmbein (Ilium) lang und tief, das Präpubis ragte ebenso weit nach vorne wie das Darmbein. Das Schambein (Pubis) war sehr kurz und das Sitzbein (Ischium) ist robust und gebogen, insbesondere bei den Chasmosaurinae. Der Oberschenkelknochen war stets länger als das Schienbein. Der Hinterfuß war kurz und kräftig und endete in vier Zehen mit stumpfen Hufen.
Integument (Haut)
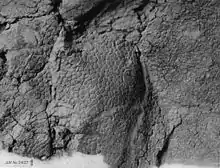
Bei mehreren Funden von Ceratopsidae lassen sich Reste des Integuments (Haut) erkennen. Bei Chasmosaurus ist ein Teil der Beckenregion bekannt, hier war die Haut mit großen, rundlichen Platten mit bis zu 55 Millimeter Durchmesser bedeckt. Die Platten verliefen in unregelmäßigen Reihen und wurden zum Bauch hin kleiner. Die großen Platten waren rund 50 bis 100 Millimeter voneinander getrennt, die Räume dazwischen waren mit kleinen, unregelmäßig geformten Plättchen bedeckt. Ein weiterer Fund der Haut am Oberschenkel von Centrosaurus zeigt ein ähnliches Muster, nur waren hier die großen Platten weiter voneinander entfernt.
Varianz und Geschlechtsdimorphismus
Da von vielen Gattungen zahlreiche fossile Überreste vorhanden sind, lässt sich die Varianz innerhalb eines Taxons besser vergleichen als bei anderen, weniger bekannten Dinosauriern. So gibt es von Triceratops rund 50 vollständig oder teilweise erhaltene Schädel, die sich zum Teil leicht voneinander unterscheiden. Diese Unterschiede haben dazu geführt, dass 16 Arten der Gattung Triceratops beschrieben wurden. Die heutige Sichtweise geht eher davon aus, dass es sich dabei um innerartliche Variationen handelt, und fasst alle Funde zu einer oder höchstens zwei Arten zusammen.
Damit zusammenhängend und auch verbunden mit der Suche nach der Funktion der Hörner und Nackenschilde ist die Frage eines Geschlechtsdimorphismus bei den Ceratopsidae, das heißt, ob es Unterschiede im Körperbau zwischen Männchen und Weibchen gab. Ein solcher Geschlechtsdimorphismus wurde bei mehreren Dinosauriern festgestellt, unter anderem auch bei Protoceratops, einem urtümlichen Vorfahren der Ceratopsidae. Auch bei den Ceratopsidae sind verschiedentlich geringe Geschlechtsdimorphismen postuliert worden, vorwiegend im Bereich der Hörner und des Nackenschildes. Auch wenn innerhalb eines Taxons zwei Morphen unterscheidbar sind, lässt sich nicht eindeutig sagen, ob dies auf Geschlechtsdimorphismus, auf mehrere Arten oder auf eine innerartliche Variation zurückzuführen ist. Dementsprechend gibt es keinen unzweifelhaften Beleg für einen Geschlechtsdimorphismus bei den Ceratopsidae – weder in der Körpergröße noch im Bau des Kopfschmuckes.[4]
Paläobiologie
Sozialverhalten und Lebensraum
Fossile Überreste von Ceratopsidae stammen meist von Einzeltieren, es gibt aber auch zahlreiche bone beds („Knochenlager“), bei denen die häufig zerstückelten Knochen von dutzenden, manchmal sogar tausenden Individuen gefunden wurden. Solche bone beds sind beispielsweise von Anchiceratops, Centrosaurus, Chasmosaurus, Einiosaurus, Pachyrhinosaurus und Styracosaurus bekannt, nicht aber von Triceratops, einem der häufigsten Ceratopsidae. Nahezu immer enthalten diese bone beds die Überreste einer einzigen Art und setzen sich aus Überresten von Jungtieren, subadulten (halbwüchsigen) und ausgewachsenen Tieren zusammen, während Einzelfunde meist von ausgewachsenen Tieren stammen.
Funde von Überresten mehrerer Tiere an einem Ort müssen nicht zwangsläufig auf ein Gruppenleben hindeuten, sondern können auch ablagerungstechnisch bedingt sein. Es ist auch denkbar, dass in Dürreperioden viele ansonsten einzelgängerische Tiere bei Wasserstellen zusammengekommen sind und dort aufgrund des Versiegens der Quelle den Tod fanden. Tatsächlich deuten Gesteinsuntersuchungen aus dem Zeitraum der Ceratopsidae auf ein trockenes, jahreszeitlich stark schwankendes Klima hin.
Aussagen über das Sozialverhalten der Ceratopsidae sind dementsprechend schwierig und es kann nicht davon ausgegangen werden, dass alle Arten in Gruppen lebten. Die Untersuchungen der Massenablagerungen lassen aber den Schluss zu, dass einige Arten zumindest einen Teil des Jahres Verbände mit Artgenossen bildeten.
Auch über den bevorzugten Lebensraum gibt es noch Unklarheiten. Generell dürften sie aber eher offene Habitate bewohnt haben. Einige Untersuchungen kommen zu dem Ergebnis, dass diese Tiere in Küstengebieten häufiger als im Landesinneren waren. D. Brinkmann et al.[5] halten ein Wanderverhalten für denkbar. Demzufolge könnten sie allein oder in kleinen Gruppen in Küstengebieten gelebt und sich dann zu großen Herden zusammengeschlossen haben, möglicherweise im Zusammenhang mit der Fortpflanzung.
Funktion der Hörner und Nackenschilde

Zur Funktion der Hörner und Nackenschilde sind mehrere Hypothesen aufgestellt worden. Die gängigste geht davon aus, dass sie der Verteidigung gegenüber Fressfeinden dienten. Die Hörner seien Stoßwaffen und die Schilde schützten den Nacken gegenüber Bissen. Hauptfressfeinde in diesem Szenario sind die Tyrannosauridae, insbesondere das „Late Cretaceous All-Star Game“[6] zwischen Tyrannosaurus und Triceratops ist ein beliebtes Motiv der populären Dinosaurierdarstellung. Diese Hypothese hat aber einige Schwachpunkte: Wenn Verteidigung das Hauptmotiv für die Entwicklung dieser Kopfauswüchse wäre, hätte nicht jede Ceratopsidae-Gattung ihre eigene Form der Hörner und Schilde entwickelt, von denen einige nutzlos oder sogar kontraproduktiv wären, wie die Rückbildung der Überaugenhörner bei vielen Centrosaurinae oder die hakenförmig nach vorn gebogenen Stacheln am Schildrand von Centrosaurus. Darüber hinaus waren die Schilde sehr dünn, sie maßen oft unter 2 Millimeter. Daneben waren sie mit Fenstern ausgestattet und mit zahlreichen Blutgefäßen versehen und daher kaum als Schutz vor Nackenbissen geeignet.
Eine andere Hypothese sieht den Nackenschild als Ansatzstelle für eine stark vergrößerte Kaumuskulatur. Morphologische Gründe sprechen dagegen: So ist die Fenestra supratemporalis (das obere Schädelfenster der Schläfengegend) relativ klein, und eine glatte Stelle daneben wurde bereits als Ansatzpunkt der Kaumuskulatur ausgemacht, außerdem spricht auch der Bau der Oberfläche der Schilde dagegen. Schließlich ist zu bezweifeln, ob ein über einen Meter langer Kaumuskel tatsächlich einen evolutiven Vorteil gebracht hätte, da eine Verlängerung eines Muskels nicht im gleichen Ausmaß eine Kräftigung mit sich bringt.
Eine dritte Hypothese, die erstmals von Wheeler 1978[7] formuliert wurde, sieht die Hörner und Schilde als Werkzeuge zur Thermoregulation. Diese Sichtweise stützt sich auf die Anzeichen von zahlreichen Blutgefäßen in diesen Strukturen verbunden mit dem Oberflächenzuwachs – der Nackenschild konnte mehr als einen Quadratmeter ausmachen. Zwar ist die Thermoregulationshypothese nicht unplausibel, sie erklärt aber nicht die starken morphologischen Unterschiede bei den einzelnen Gattungen.

Eine vierte, erstmals 1961 von L. Davitashvili[8] formulierte Hypothese besagt, dass die Hörner und Nackenschilde der Kommunikation und der Auseinandersetzung um Reviere oder Paarungspartner gedient hätten. Für diese Sichtweise spricht, dass sich die einzelnen Taxa oft nur im Bau der Hörner und Schilde unterschieden. Demzufolge könnten die Merkmale eine entscheidende Rolle dabei gespielt haben, wie sich die einzelnen Arten untereinander erkannt haben. Ein weiterer Punkt ist – wie Funde von Jungtieren zeigen –, dass diese Merkmale erst nach dem Erreichen der Erwachsenengröße voll ausgebildet waren, also erst zu dem Zeitpunkt, wo der Kampf um Reviere oder Paarungspartner einsetzt. Einige Stichwunden in der Wangenregion und den Schilden ließen sich auf Kämpfe mit Artgenossen zurückführen. In Zusammenhang mit der vermuteten zumindest zeitweiligen Bildung von größeren Herden lässt sich ein Szenario erahnen, in dem die Zurschaustellung der Kopfauswüchse, Drohgebärden oder Kämpfe zwischen Artgenossen eine Rolle spielten, bei denen es um Wettstreite um Territorien, das Paarungsvorrecht oder die Bildung von Rangordnungen ging.[9] Eine Untersuchung aus dem Jahr 2009 fand viele Verletzungen in den Schilden von Triceratops, verglichen mit wenigen Wunden bei Centrosaurus. Dieser Befund spricht dafür, dass Triceratops seine Hörner bei Kämpfen mit Artgenossen einsetzte, während bei Centrosaurus eher die Zurschaustellung die Hauptfunktion gewesen sein könnte (oder die Stöße gegen andere Körperteile gerichtet waren).[10]
Es ist durchaus denkbar, dass die Hörner und Nackenschilde mehreren Zwecken dienten, und außer der Ansatzstelle für eine überproportionale Kaumuskulatur könnten alle oben vorgestellten Hypothesen zutreffen. Auch in Analogie zu heute lebenden Tierarten gilt die Hypothese der Identifikation und Auseinandersetzung mit Artgenossen nach heutigem Kenntnisstand als am plausibelsten.[11]
Haltung und Fortbewegung

Die Frage nach der Körperhaltung und Fortbewegung der Ceratopsidae hat sich als nicht leicht zu beantworten herausgestellt. Viele alte Zeichnungen ebenso wie die ersten Skelett-Aufbauten (Triceratops 1904 und Chasmosaurus 1923) zeigten die Tiere mit aufgerichteten, senkrechten Hinterbeinen und weit auseinandergespreizten, abgeknickten Vorderbeinen, bei denen die Oberarmknochen nahezu waagrecht gehalten wurden. Die Gründe dafür lagen wohl im Bau des Kopfes des Oberarmknochens und des Rabenbeins. Spätere Untersuchungen, etwa von Robert Bakker,[12] sind hingegen zu dem Schluss gekommen, die Ceratopsidae hätten die Vorderbeine senkrecht und säulenartig gehalten. Anatomische Untersuchungen widersprachen dem und plädierten für eine halbaufgerichtete Haltung der Vorderbeine mit leicht nach außen gebogenen Ellbogen.[13] Neben anatomischen Untersuchungen wurden auch Ichnofossilien (versteinerte Fußabdrücke) herangezogen. So kamen Paul und Christiansen zu dem Ergebnis, diese Tiere hätten ihre Vorderbeine annähernd gerade, also parallel zur Körperlängsebene (parasagittal), und mit nur leicht abgewinkelten Ellbogen gehalten.[14]
Ebenso umstritten ist die Frage, auf welche Weise und mit welcher Geschwindigkeit sich diese Tiere fortbewegen konnten. Aufgrund ihres vermuteten hohen Gewichts und ihrer massiven, eher kurzen Gliedmaßen gelten sie eher als schwerfällige, langsame Tiere. Nach den Berechnungen von R. A. Thulborn[15] konnten sie eine Geschwindigkeit von 25 km/h erreichen. Paul und Christiansen[14] kommen zu der Vermutung, dass sie zu einer dem Galopp ähnlichen Fortbewegung fähig waren und eine Geschwindigkeit ähnlich der der Nashörner (über 40 km/h) erreichen konnten.
Nahrung

Der Bau der Kiefer der Ceratopsidae mit den Zahnbatterien mit senkrechter Okklusionsfläche ist einzigartig unter den Wirbeltieren. Diese Spezialisation ist bei allen Ceratopsidae gleich und hat sich in der Entwicklungsgeschichte dieser Dinosaurier nicht mehr verändert. Die Zähne waren für eine schneidende, nicht aber mahlende Bewegung ausgerichtet. Das Kiefergelenk saß tiefer als die Zahnreihe und der Muskelfortsatz (Processus coronoideus) am Unterkieferast war vergrößert und bot den Ansatzpunkt für eine vermutlich sehr kräftige Kaumuskulatur, all das deutet auf eine hohe Beißkraft dieser Tiere hin. Die schmale, zugespitzte Schnauze, die die Ceratopsidae mit allen Ceratopsia gemeinsam hatten, gilt gemeinhin als Anzeichen für eine selektive Nahrungsaufnahme. Sie dürfte eher zum Zupacken und Ausrupfen, nicht aber zum Abbeißen geeignet gewesen sein.
Möglicherweise hatten sie einen vergrößerten Verdauungstrakt mit symbiotischen Mikroorganismen, zur besseren Verwertung der schwer verdaulichen Pflanzennahrung. Denkbar sind beispielsweise ein mehrkammeriger Magen (wie bei den Wiederkäuern) oder ein vergrößerter Darmtrakt (wie bei den Unpaarhufern).[16]
Da der Kopf nahe beim Boden gehalten wurde, haben sie vermutlich vorwiegend krautige Pflanzen gefressen. Es ist auch denkbar, dass sie mit ihren Hörnern und Schnäbeln höhere Pflanzen heruntergebogen oder abgebrochen haben. Welche Pflanzen sie genau gefressen haben, ist allerdings nicht geklärt. Aufgrund ihrer großen Ausmaße, ihrer Zahnbatterien und ihrer möglichen Tendenz, größere Herden zu bilden, haben sie vermutlich minderwertige, stark faserhaltige Pflanzen verzehrt.[16] Häufig werden Palmfarne und Palmen als Hauptnahrung angenommen. Fossile Überreste dieser Pflanzen finden sich jedoch kaum in den Gesteinsschichten, aus denen die Ceratopsidae bekannt sind. Es ist darum denkbar, dass Farne eine wichtige Rolle bei der Ernährung der Ceratopsidae gespielt haben. Coe et al.[17] halten es für möglich, dass es regelrechte Farnprärien gab, die diese Tiere ernährten.
Fortpflanzung und Entwicklung

Über die Fortpflanzung und die Individualentwicklung der Ceratopsidae ist verglichen mit anderen Dinosauriern relativ wenig bekannt. Wie alle Dinosaurier haben sie Eier gelegt, eindeutig den Ceratopsidae zuordenbare Eierfunde gibt es jedoch bislang nicht. Ebenso wenig sind bislang Nester oder Schlüpflinge zutage gefördert worden.
Anhand der Funde in den Massenablagerungen konnten von Sampson et al. drei Altersabstufungen identifiziert werden: juvenil (jugendlich), subadult (halbwüchsig) und adult (ausgewachsen).[18] Dabei zeigt sich, dass die Unterscheidungsmerkmale im Bereich der Hörner und Nackenschilde sich erst bei den ausgewachsenen Tieren herausbildeten. Dementsprechend ist es sehr schwierig, die Jungtiere verschiedener Gattungen voneinander zu unterscheiden – zumindest der Centrosaurinae, der untersuchten Gruppe. Sampson et al. plädieren daher dafür, Taxa, die nur anhand von Fossilfunden von jugendlichen Tieren beschrieben wurden (Brachyceratops und Monoclonius), als Nomina dubia zu führen.
Systematik
Äußere Systematik und Entwicklungsgeschichte
Die Ceratopsidae werden innerhalb der Ceratopsia in die Neoceratopsia eingegliedert und hier wiederum in die Coronosauria. Ihre Schwestergruppe ist Zuniceratops, mit dem sie die Ceratopsoidea bilden, entfernter sind sie mit den Protoceratopsidae und Leptoceratopsidae verwandt. Das kommt in einem vereinfachten Kladogramm der Ceratopsia zum Ausdruck:[19]
| Ceratopsia |
| ||||||||||||||||||||||||||||||||||||
Ob die vorrangig aus Nordamerika bekannten Leptoceratopsidae oder die asiatischen Protoceratopsidae näher mit den Ceratopsoidea verwandt sind, ist umstritten. Im Gegensatz zum hier dargestellten Standpunkt geht eine Reihe von Untersuchungen[20] davon aus, dass die Protoceratopsidae die Schwestergruppe der Ceratopsoidea seien.
Die Ceratopsidae kamen fast ausschließlich im westlichen Nordamerika vor, Funde sind vom heutigen Alaska bis nach Mexiko bekannt. Da auch ein nächster Verwandter, Zuniceratops, dort lebte, dürfte die Gruppe sich auch auf diesem Kontinent entwickelt haben. Die einzige bekannte Ausnahme stellt der erst 2010 beschriebene Sinoceratops dar, der im heutigen China gefunden wurde.[2]
Die Ceratopsidae sind nur aus einem erdgeschichtlich relativ kurzen Zeitraum, vom Campanium bis zum Maastrichtium, aus der Oberkreide bekannt und damit rund 83 bis 66 Millionen Jahre alt.
Innere Systematik
Die Ceratopsidae lassen sich gut in zwei Unterfamilien einteilen, die Centrosaurinae und die Chasmosaurinae. Die Centrosaurinae besaßen in der Regel ein längeres Nasenhorn, kürzere oder fehlende Überaugenhörner und einen kürzeren Nackenschild. Bei den Chasmosaurinae war meistens das Nasenhorn kurz, die Überaugenhörner und der Nackenschild hingegen verlängert. (Näheres siehe unter Merkmale.)
Die nachfolgende Gattungsliste folgt P. Dodson et al. (2004)[4], schließt aber seither beschriebene Gattungen mit ein.
- Centrosaurinae
- Chasmosaurinae
- Agujaceratops
- Anchiceratops
- Arrhinoceratops
- Chasmosaurus
- Coahuilaceratops
- Diceratus (früher Diceratops)
- Eotriceratops
- Kosmoceratops[25]
- Medusaceratops
- Mercuriceratops[26]
- Mojoceratops
- Ojoceratops
- Pentaceratops
- Tatankaceratops
- Titanoceratops
- Torosaurus (wahrscheinlich synonym mit Triceratops[27])
- Triceratops
- Utahceratops
- Vagaceratops[25]
- Nomina dubia
Mehrere Untersuchungen kamen zu dem Ergebnis, dass sowohl die Ceratopsidae als auch die beiden Unterfamilien mit großer Wahrscheinlichkeit monophyletisch sind. Die größte Unsicherheit ergab sich bei der Stellung von Einiosaurus innerhalb der Centrosaurinae, der entweder die Schwestergruppe aller übrigen Centrosaurinae, der Achelousaurus-Pachyrhinosaurus-Klade oder der Styracosaurus-Centrosaurus-Klade ist. Die übrigen Abstammungslinien sind stabil und sind in nachfolgendem Kladogramm dargestellt.[4]
| Ceratopsidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Der 2007 erstbeschriebene Albertaceratops wird von seinen Erforschern innerhalb der Centrosaurinae als Schwestertaxon der übrigen Vertreter dieser Gruppe eingeordnet.[28]
Ceratopsidae und Menschen
Entdeckungs- und Forschungsgeschichte

Die ersten bekannten Ceratopsidae waren 1872 Agathaumas, 1874 Polyonax und 1876 Dysganus, alle beschrieben von Cope. Die rudimentären Funde ließen zunächst kaum Rückschlüsse auf die Tiere zu, was auch daran deutlich wird, dass Marsh 1887 einen Fund Bison alticornis nannte, ihn also für einen langhörnigen Bison hielt. Erst mit den Funden von Triceratops 1889 und Torosaurus 1891 (beide ebenfalls von Marsh beschrieben) ließ sich erstmals das tatsächliche Aussehen der Ceratopsidae erahnen. Den Namen Ceratopsidae prägte Marsh 1890, namensgebend war Ceratops, ein schlecht erhaltener Fund aus dem Jahr 1888. Obwohl die Formen „Ceratopia“ und „Ceratopidae“ (jeweils ohne s) grammatikalisch richtig wären, verwenden viele Fachwerke dennoch weiterhin die inkorrekten, von Marsh geprägten Bezeichnungen.
Anfang des 20. Jahrhunderts wurden mit Centrosaurus und Diceratus weitere Vertreter dieser Gruppe entdeckt, und 1904 wurde das erste montierte Skelett im National Museum of Natural History in Washington errichtet. 1907 erschien mit „The Ceratopsia“[29] die erste umfassende Monographie zu diesen Tieren, die von Lull basierend auf den Studien der kurz zuvor verstorbenen Marsh und Hatcher verfasst wurde. In den 1910er- und 1920er-Jahren war Kanada Fundstätte zahlreicher Gattungen wie Anchiceratops, Chasmosaurus und Styracosaurus. 1933 verfasste Lull seine revidierten Studien[30], in denen er eine Gruppe mit kurzem Nackenschild und eine Gruppe mit langem Nackenschild unterschied. Bis auf die Zugehörigkeit von Triceratops, der trotz eines kurzen Nackenschilds zu den langschildigen Chasmosaurinae gerechnet wird, entsprach diese Einteilung bereits den heute anerkannten Unterfamilien.
Nach dem Zweiten Weltkrieg wurde mit Pachyrhinosaurus (1950) zunächst nur eine einzige neue Gattung entdeckt, zunehmend rückten paläobiologische und -ökologische Studien in den Vordergrund. Über 30 Jahre nach Pachyrhinosaurus wurde 1986 mit Avaceratops der nächste Vertreter der Ceratopsidae beschrieben. In den 1990er-Jahren traten Achelousaurus und Einiosaurus ans Tageslicht. Seit dieser Zeit wird auch die Systematik dieser Familie mit kladistischen Methoden untersucht, unter anderem von Thomas Lehman und Peter Dodson.
Auch im 21. Jahrhundert wurden neue Gattungen beschrieben, Agujaceratops, Albertaceratops und Eotriceratops in den Jahren 2006 und 2007. Die offen gebliebenen Fragen lassen auch intensive zukünftige Forschungen erwarten.
In der Populärkultur
Einige Vertreter der Ceratopsidae, insbesondere Triceratops, zählen zu den bekanntesten Dinosauriern und haben einen Fixplatz in populären Werken über Dinosaurier. Schon im Film Die verlorene Welt aus dem Jahr 1925 waren Triceratops und Agathaumas zu sehen. In Jurassic Park erscheinen sie ebenso wie im Zeichentrickfilm In einem Land vor unserer Zeit. Auch Dokumentarfilme beschäftigen sich mit diesen Tieren, so beispielsweise Dinosaurier – Im Reich der Giganten (Walking with Dinosaurs) oder Kampf der Dinosaurier (The Truth About Killer Dinosaurs), wo der Kampf zwischen Triceratops und Tyrannosaurus thematisiert wird.
Literatur
- Peter Dodson, Catherine A. Forster, Scott D. Sampson: Ceratopsidae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Ausgabe. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 494–513.
- David E. Fastovsky, David B. Weishampel: The Evolution and Extinction of the Dinosaurs. 2. Ausgabe. Cambridge University Press, Cambridge u. a. 2005, ISBN 0-521-81172-4.
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, ISBN 978-0-691-13720-9, S. 257–272, Online (Memento des Originals vom 13. Juli 2015 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis..
- Xing Xu, KeBai Wang, XiJin Zhao, DunJing Li: First ceratopsid dinosaur from China and its biogeographical implications. In: Chinese Science Bulletin. Bd. 55, Nr. 16, 2010, ISSN 1001-6538, S. 1631–1635, doi:10.1007/s11434-009-3614-5.
- Dodson et al. (2004), S. 494.
- Dodson et al. (2004).
- Donald B. Brinkman, Michael J. Ryan, David A. Eberth: The paleogeographic and stratigraphic distribution of ceratopsids (Ornithischia) in the Upper Judith River Group of Western Canada. In: Palaios. Bd. 13, Nr. 2, 1998, ISSN 0883-1351, S. 160–169, doi:10.1043/0883-1351(1998)013<0160:TPASDO>2.0.CO;2.
- Fastovsky & Weishampel (2005), S. 171.
- P. E. Wheeler: Elaborate CNS cooling structures in large dinosaurs. In: Nature. Bd. 275, Nr. 5679, 1978, S. 441–443, doi:10.1038/275441a0.
- Лео Шйович Давиташвили: Теория полового отбора. Изд-во Академии наук СССР, Москва 1961.
- Fastovsky & Weishampel (2005), S. 174–175.
- Andrew A. Farke, Ewan D. S. Wolff, Darren H. Tanke: Evidence of Combat in Triceratops. In: PLoS ONE. Bd. 4, Nr. 1, 2009, e4252, doi:10.1371/journal.pone.0004252.
- Abschnitt Horns and Frills. In: Dodson et al. (2004), S. 512.
- Robert T. Bakker: The Return of the Dancing Dinosaurs. In: Sylvia J. Czerkas, Everett C. Olson (Hrsg.): Dinosaurs Past and Present. Band 1. Natural History Museum of Los Angeles County, Los Angeles CA 1987, ISBN 0-938644-24-6, S. 38–69.
- Rolf E. Johnson, John H. Ostrom: The forelimb of Torosaurus and an analysis of the posture and gait of ceratopsian dinosaurs. In: Jeff Thomason (Hrsg.): Functional Morphology in Vertebrate Paleontology. Cambridge University Press, Cambridge u. a. 1995, ISBN 0-521-44095-5, S. 205–218; oder Peter Dodson, James O. Farlow: The forelimb carriage of ceratopsid dinosaurs. In: Donald L. Wolberg, Edmund Stump, Gary Rosenberg (Hrsg.): Dinofest International. Proceedings of a symposium held at Arizona State University. Academy of Natural Sciences, Philadelphia PA 1997, ISBN 0-935868-94-1, S. 393–398.
- Gregory S. Paul, Per Christiansen: Forelimb Posture in Neoceratopsian Dinosaurs: Implications for Gait and Locomotion. In: Paleobiology. Bd. 26, Nr. 3, 2000, ISSN 0094-8373, S. 450–465, doi:10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2.
- Richard A. Thulborn: Speeds and gaits of dinosaurs. In: Palaeogeography, Palaeoclimatology, Palaeoecology. Bd. 38, Nr. 3/4, 1982, ISSN 0031-0182, S. 227–256, doi:10.1016/0031-0182(82)90005-0.
- Abschnitt Feeding, Diet and Respiration. In: Dodson et al. (2004), S. 511–512.
- M. J. Coe, David L. Dilcher, James O. Farlow, David M. Jarzen, Dale A. Russell: Dinosaurs and Land Plants. In: Else Marie Friis, William Gilbert Chaloner, Peter R. Crane (Hrsg.): The Origins of Angiosperms and their Biological Consequences. Cambridge University Press, Cambridge u. a. 1987, ISBN 0-521-32357-6, S. 225–259.
- Scott D. Sampson, Michael Ryan, Darren H. Tanke: Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): Taxonomic and behavioral implications. In: Zoological Journal of the Linnean Society. Bd. 121, Nr. 3, 1997, ISSN 0024-4082, S. 293–337, doi:10.1111/j.1096-3642.1997.tb00340.x.
- nach Xing Xu, Catherine A. Forster, James M. Clark, Jinyou Mo: A basal ceratopsian with transitional features from the Late Jurassic of northwestern China. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 273, Nr. 1598, 2006, ISSN 0950-1193, S. 2135–2140, doi:10.1098/rspb.2006.3566, Digitalisat (PDF; 916,04 kB); und Dodson et al., (2004).
- etwa Peter J. Makovicky: A Montanoceratops cerorhynchus (Dinosauria: Ceratopsia) braincase from the Horseshoe Canyon Formation of Alberta. In: Darren H. Tanke, Kenneth Carpenter (Hrsg.): Mesozoic Vertebrate Life. Indiana University Press u. a., Bloomington IN u. a. 2001, ISBN 0-253-33907-3, S. 243–262; oder Brenda Chinnery: Description of Prenoceratops pieganensis gen et sp. nov. (Dinosauria: Neoceratopsia) from the Two Medicine Formation of Montana. In: Journal of Vertebrate Paleontology. Bd. 24, Nr. 3, 2004, ISSN 0272-4634, S. 572–590, doi:10.1671/0272-4634(2004)024[0572:DOPPGE]2.0.CO;2.
- Scott D. Sampson, Eric K. Lund, Mark A. Loewen, Andrew A. Farke, Katherine E. Clayton: A remarkable short-snouted horned dinosaur from the Late Cretaceous (late Campanian) of southern Laramidia. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 280, Nr. 1766, 2013, ISSN 0950-1193, S. 1–7, doi:10.1098/rspb.2013.1186.
- Andrew T. McDonald, John R. Horner: New Material of „Styracosaurus“ ovatus from the Two Medicine Formation of Montana. In: Michael J. Ryan, Brenda J. Chinnery-Allgeier, David A. Eberth (Hrsg.): New Perspectives on Horned Dinosaurs. The Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press, Bloomington IN u. a. 2010, ISBN 978-0-253-35358-0, S. 156–168.
- Andrew A. Farke, Michael J. Ryan, Paul M. Barrett, Darren H. Tanke, Dennis R. Braman, Mark A. Loewen, Mark R. Graham: A new centrosaurine from the Late Cretaceous of Alberta, Canada, and the evolution of parietal ornamentation in horned dinosaurs. In: Acta Palaeontologica Polonica. Bd. 56, Nr. 4, 2011, ISSN 0567-7920, S. 691–702, doi:10.4202/app.2010.0121.
- David C. Evans, Michael J. Ryan. Cranial Anatomy of Wendiceratops pinhornensis gen. et sp. nov., a Centrosaurine Ceratopsid (Dinosauria: Ornithischia) from the Oldman Formation (Campanian), Alberta, Canada, and the Evolution of Ceratopsid Nasal Ornamentation. PLOS ONE, 2015; 10 (7): e0130007 DOI: 10.1371/journal.pone.0130007
- Scott D. Sampson, Mark A. Loewen, Andrew A. Farke, Eric M. Roberts, Catherine A. Forster, Joshua A. Smith, Alan L. Titus: New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endemism. In: PLoS ONE. Bd. 5, Nr. 9, 2010, e12292, doi:10.1371/journal.pone.0012292.
- Michael J. Ryan, David C. Evans, Philip J. Currie, Mark A. Loewen: A new chasmosaurine from northern Laramidia expands frill disparity in ceratopsid dinosaurs. In: Naturwissenschaften. Bd. 101, Nr. 6, 2014, S. 505–512, doi:10.1007/s00114-014-1183-1.
- John B. Scannella, John R. Horner: Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. In: Journal of Vertebrate Paleontology. Bd. 30, Nr. 4, 2010, S. 1157–1168, doi:10.1080/02724634.2010.483632.
- Michael J. Ryan: A new basal centrosaurine ceratopsid from the Oldman Formation, southeastern Alberta. In: Journal of Paleontology. Bd. 81, Nr. 2, 2007, ISSN 0022-3360, S. 376–396, doi:10.1666/0022-3360(2007)81[376:ANBCCF]2.0.CO;2.
- John B. Hatcher: The Ceratopsia (= Monographs of the United States Geological Survey. 49, ISSN 0886-7550). Based on preliminary Studies by Othniel C. Marsh. Edited and completed by Richard S. Lull. U.S. Government Printing Office, Washington DC 1907.
- Richard Swann Lull: A revision of the Ceratopsia or horned dinosaurs (= Memoirs of the Peabody Museum of Natural History. Bd. 3, Tl. 3, ZDB-ID 1002336-7). The Tuttle – Morehouse & Taylor, New Haven CT 1933, Digitalisat.