Buschschliefer
Der Buschschliefer (Heterohyrax brucei), teilweise auch Steppenschliefer, ist die einzige Art der gleichnamigen Gattung innerhalb der Schliefer. Sein Verbreitungsgebiet reicht vom nordöstlichen über das östliche bis zum südöstlichen Afrika, zusätzlich kommt die Art auch im zentralen und südwestlichen Afrika vor. Der Lebensraum der Tiere umfasst teils offene Landschaften durchsetzt mit Kopjes oder Steilklippen, sie treten zusätzlich auch in Waldgebieten auf. Im äußeren Erscheinungsbild ähnelt der Buschschliefer wie andere Schliefer auch einem Meerschweinchen, die Gliedmaßen sind kurz, der Schwanz fehlt. Charakteristisch ist ein gelber Fleck auf dem Rücken, der sich deutlich von der grauen bis braunen Fellfärbung der Oberseite abhebt. Die Unterseite zeigt sich auffallend heller. Zudem sind verschiedene helle Gesichtsflecken und lange Tasthaare ausgebildet.
| Buschschliefer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Buschschliefer (Heterohyrax brucei) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Heterohyrax | ||||||||||||
| J. E. Gray, 1868 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Heterohyrax brucei | ||||||||||||
| (J. E. Gray, 1868) |
Die Lebensweise des Buschschliefers gleicht in vielen Aspekten der des Klippschliefers. Teilweise kommen beide Arten sympatrisch vor und bewohnen die gleiche Felsformation. Die Tiere sind gesellig und bilden Kolonien mit einem dominanten Männchen, mehreren Weibchen und den Jungtieren. Sie nutzen Felsspalten und -höhlen als Unterschlupf, können aber auch in Bäumen klettern. Das Männchen verteidigt sein Revier und stößt markante, weit tragende Territorialrufe aus. Die Hauptnahrung besteht aus weicher Pflanzenkost mit einem geringen Anteil an Gräsern. In der Regel fressen die Tiere gemeinsam am frühen Morgen und am Abend. Die Fortpflanzung findet weitgehend innerhalb der Kolonie statt. Weibchen bringen einmal jährlich einen Wurf mit bis zu zwei Jungen zur Welt. Als einer der bedeutendsten Fressfeinde tritt der Kaffernadler in Erscheinung, der sich fast ausschließlich von Schliefern ernährt.
Die Erstbeschreibung des Buschschliefers als Art und Gattung erfolgte im Jahr 1868, allerdings nicht gemeinsam. Berichte über die Tiere reichen bis in die zweite Hälfte des 18. Jahrhunderts zurück. Die mehrfache Erwähnung der Tiere in dieser forschungsgeschichtlich frühen Zeit führte zu einer Kontroverse über den tatsächlichen wissenschaftlichen Artnamen, die im Übergang vom 19. zum 20. Jahrhundert bestand. Fossil trat der Buschschliefer erstmals zum Ende des Miozäns in Erscheinung, einzelne Funde stammen aus dem südwestlichen Afrika. Die heutigen Bestände gelten als ungefährdet.
Merkmale
Habitus

Der Buschschliefer ist ein kleines bis mittelgroßes Tier. Die Gesamtlänge von mehr als 90 untersuchten Individuen aus der Serengeti variierte von 32,5 bis 47,2 cm, das Körpergewicht betrug 1,3 bis 2,4 kg. Dagegen waren 17 vermessene Tiere aus Simbabwe mit einer Körperlänge von 46,5 bis 53,0 cm und einem Körpergewicht von 2,3 bis 3,6 kg etwas größer. Weibchen weisen durchschnittlich größere Körpermaße als Männchen auf. Allgemein hat der Buschschliefer wie andere Schliefer auch ein meerschweinchen- bis kaninchenartiges Äußeres. Die Beine sind kurz, der Schwanz ist nur rudimentär ausgebildet. Das Körperfell besitzt am Rücken und an den Seiten eine graue bis dunkel rötlichbraune Färbung, einige Individuen können aber auch weißlich gefleckt sein. Die Leithaare sind weicher als beim Klippschliefer (Procavia capensis), sie haben schwarze Spitzen und werden bis zu 30 mm lang. Die Unterwolle ist dicht, die einzelnen Haare zeichnen sich durch dunkelgraue Basen und gelblichbraune Spitzen aus. Auf dem Rücken hebt sich markant ein cremefarbener bis gelber Fleck hervor, der eine Drüse von rund 15 mm Länge umgibt. In einigen Populationen fehlt der Fleck. Die Haare des Flecks können aufgerichtet werden. Die Bauchseite ist zumeist heller getönt und steht so im Kontrast zur Färbung von Rücken und Kopf. Mitunter treten auch albinotische Tiere auf.[1] Am Kopf fallen die weißen Augenbrauen auf, die deutlich hervortreten und heller sind als beim Klippschliefer. Die Ohren sind mit 29 bis 34 mm länger als bei anderen Schliefern, ihre Form ist gerundet. Im Gesicht treten über den Augen und am Kinn bis zu 90 mm lange Vibrissen auf. Weitere Tasthaare finden sich verteilt über den gesamten Rücken, den Bauch und die Beine. Die Hände weisen vier, die Füße drei Strahlen auf. Alle sind mit hufartig platten Nägeln ausgestattet, eine Ausnahme bildet der jeweils innere Zeh der Hinterfüße, der eine gebogene Kralle trägt. Die Sohlen haben dicke Hautpolster von zumeist dunkler Farbgebung. Die Hinterfußlänge variiert von 65 bis 73 mm.[2][3][4][5]
Beim Männchen befindet sich der Penis 65 bis 82 mm vor dem Anus, die Distanz ist zwei- bis dreimal größer als bei anderen Schliefern. Der Penis, der im erigierten Zustand 60 mm Länge erreicht, weist außerdem einen komplexeren Bau auf. Hervorgerufen wird dies durch einen Fortsatz an der Spitze, der die Harnröhre umschließt. Der Querschnitt des Penis ist rundlich.[6] Weibchen besitzen zumeist ein Zitzenpaar in der Brust- und zwei in der Leistengegend. Manchmal fehlen aber die Brustzitzen.[2][3][4][5]
Schädel- und Gebissmerkmale
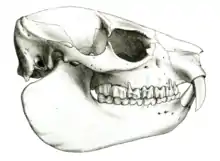
Der Schädel wird 66,8 bis 87,4 mm lang.[7] Das Scheitelbein verschmilzt mit dem Os interparietale (ein Schädelknochen zwischen dem Hinterhauptsbein und den Scheitelbeinen). Der Postorbitalbogen ist zumeist offen. Es treten Temporalleisten auf, die sich selten zu einem Scheitelkamm vereinigen, in der Regel aber enger liegen als bei den Baumschliefern (Dendrohyrax). Der aufsteigende Ast des Unterkiefers ist relativ schmal, der Kronenfortsatz erhebt sich deutlich über den Gelenkfortsatz. Der horizontale Knochenkörper zeichnet sich durch seinen sehr niedrigen Bau aus, die Unterseite der Symphyse ist zumeist glatt. Unterhalb des ersten Prämolaren ist ein Foramen mentale ausgebildet.[7][8] Das Gebiss besteht aus 34 Zähnen mit folgender Zahnformel: . Die oberen Schneidezähne ähneln den Eckzähnen (caniniform), sie stehen aber weiter auseinander als bei den anderen Schliefern. Bei Männchen zeigen sie sich dreieckig im Querschnitt mit der Spitze nach vorn zeigend, bei den Weibchen sind sie rund im Querschnitt. Zum hinteren Gebiss besteht ein Diastema von 10 bis 16 mm Länge. Die inneren Schneidezähne des Unterkiefers weisen drei kleine Spitzen auf (tricuspid oder trifid),[9] die äußeren schließen mit den oberen. Sowohl die Prämolaren als auch die Molaren haben niedrige Zahnkronen (brachyodont). Im Oberkiefer wird die Reihe der Vormahlzähne etwa genauso lang wie die der Mahlzähne. Der zweite Mahlzahn ist hier der größte Zahn in der gesamten Reihe.[2][3][4][5]

Verbreitung
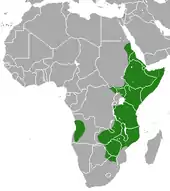
Der Buschschliefer kommt endemisch in Afrika vor. Sein Verbreitungsgebiet umfasst den östlichen Teil des Kontinents, etwa vom nordöstlichen Sudan im Norden über das Horn von Afrika bis zu den südafrikanischen Provinzen Mpumalanga und Limpopo im Süden. Zwei isolierte Populationen bestehen im Südwesten der Demokratischen Republik Kongo im zentralen Afrika und in Angola im südwestlichen Afrika. Vermeintliche Nachweise des Buschschliefers von der Halbinsel Sinai in Ägypten oder vom Ahaggar-Plateau in Algerien, beides nördliches Afrika, sind Verwechslungen mit dem Klippschliefer oder Falschzuweisungen.[10][11] Die Tiere leben in Landschaften mit Kopjes, Steilklippen oder Geröllfeldern. In einigen Regionen bewohnt der Buschschliefer auch Galeriewälder. Die Höhenverbreitung reicht in Ostafrika bis auf etwa 3800 m.[2][3][4][5]
Häufig tritt der Buschschliefer sympatrisch mit dem Klippschliefer auf, teilweise bewohnen beide Arten die gleichen Kopjes. Dieses gemischtartliche Vorkommen variiert über die Jahreszeiten, ist aber vor allem während der Fortpflanzungsphase beständig. In der Serengeti wurde eine Populationsdichte von bis zu 75 Individuen je Hektar eines Kopjes festgestellt, wenn die Felsen vom Buschschliefer ausschließlich bewohnt werden. Beim Vorkommen beider Arten an einem Kopje geht die Individuendichte auf etwa 28 Individuen je Hektar Kopje zurück. Während eines 17-jährigen Untersuchungszeitraumes konnten an einem 3600 m² großen Kopje beträchtliche Schwankungen an Buschschliefern festgestellt werden. So variierte die tatsächliche Individuenanzahl im Zeitraum von 1971 und 1976 zwischen 19 und 20 Tiere mit einem kurzen Einbruch 1975. Bis 1982 fiel sie auf 6 Individuen ab und stieg dann bis 1988 wieder auf 8 Tiere an. Die Häufigkeit des Buschschliefers in der Savannenregion korreliert dabei mit der Anzahl der Felsen, weniger mit der Dichte der Untergrundvegetation.[12][13] Im Matobo-Nationalpark in Simbabwe gab es in einem 13-jährigen Untersuchungszeitraum ähnliche Fluktuationen. Die durchschnittliche Individuenanzahl je Hektar stieg zunächst von 0,5 bis 1,1 Tieren im Zeitraum 1993/1995 auf bis zu 1,9 im Jahr 1998 an, die geschätzte Gesamtpopulation nahm dabei von 21.800 auf 81.400 Buschschliefer zu.[14][15] In der Folgezeit variierte die Populationsdichte stark. Im Jahr 2002 lag sie bei rund 1,2 Tieren je Hektar, erholte sich dann im Folgejahr auf rund 1,4 und brach wiederum ein Jahr später auf 0,8 Tiere ein. Die tatsächliche Individuenanzahl an 28 gezählten Fundstellen verringerte sich dadurch von 700 Tieren im Jahr 2003 auf 400 Tiere im Jahr 2004, was einen Rückgang von 43 % darstellt.[1][16] Der jeweilige Anstieg der Population im Untersuchungszeitraum stimmte dabei jeweils mit regenreichen Perioden und einer erhöhten Anzahl an Nachwuchs überein. Demnach kann der Niederschlag als beeinflussender Faktor angesehen werden.[17][4][5]
Lebensweise
Territorialverhalten

Der Buschschliefer ist überwiegend tagaktiv und kann agil klettern, an den Sohlen seiner Füße befinden sich zahlreiche Drüsen für Sekrete, die den Griff festigen. Er bewohnt hauptsächlich Landschaften, die von Kopjes durchsetzt sind. Kopjes bilden einzelnstehende Felsformationen („Inselberge“) mit mehreren tausend Quadratmeter Grundfläche und bis zu 40 m Höhe.[18] Auf diesen Kopjes lebt der Buschschliefer gesellig in Kolonien von bis zu 37 Individuen. Die Kolonien setzen sich aus einem dominanten Männchen, mehreren geschlechtsreifen Weibchen und den Jungtieren zusammen. Je nach Größe der Kopjes variieren auch die Gruppengrößen. Zwei in den 1970er Jahren untersuchte Kopjes in der Serengeti von 3600 und 2800 m² Fläche beherbergten über sechs Jahre der Dokumentation Gruppen von durchschnittlich 16,3 und 8,6 Individuen. In der Regel nutzt die Kolonie die gesamte Felsformation als Heimatgebiet. Inwiefern unter den Weibchen eine Rangfolge besteht ist unklar, häufig führen aber ältere Individuen eine Gruppe an, etwa auf dem Weg zu den Fressplätzen. Auf größeren Kopjes können auch mehrere Kolonien vorkommen, wie dies eine ebenfalls in den 1970er Jahren analysierte Felsformation von 14.750 m² Fläche zeigt, auf der 112 Buschschliefer lebten. Das soziale Gefüge ist dann komplexer als in den einzeln lebenden Kolonien. Die Weibchen einer Gruppen nutzen Aktionsräume, die bei den einzelnen Individuen zwischen 1080 und 4050 m² groß sind (durchschnittlich 2100 m²), der gemeinsam genutzte Raum bildet die Fläche der Kolonie. Manche Weibchen können aber auch in allen Kolonien eines großen Kopjes auftreten. Die dominanten Männchen beanspruchen ein eigenes Territorium, das etwa 60 % des gemeinsamen Aktionsraums der Weibchen umfasst, und verteidigen ihr Revier gegen Geschlechtsgenossen. Außerdem verpaaren sie sich häufiger mit älteren Weibchen über 28 Monaten. Abweichend von den kleineren Kopjes mit nur einer Kolonie treten an größeren Kopjes in den Randgebieten der Gruppen ebenfalls Männchen auf. Diese als „Randmännchen“ bezeichneten untergeordneten Individuen haben untereinander eine Rangfolge, gelegentlich übernimmt das ranghöchste beim Verschwinden des territorialen Männchens die Kolonie. Die „Randmännchen“ paaren sich überwiegend mit jüngeren Weibchen. Als dritte Gruppe kommen junge und späte Auswanderer vor. Zumeist handelt es sich um junge, geschlechtsreife Männchen, die ihre Heimatgruppe verlassen und umherwandern beziehungsweise auf großen Kopjes zu „Randmännchen“ werden. Die Besiedlungsweise der einzelnen Kopjes ist dynamisch und variiert über die Zeit, die Abwanderung der jungen Männchen wird als wichtiger Regulationsmechanismus aufgefasst, jedoch ist auch die Wanderung von Weibchen belegt.[19][12][13] Möglicherweise hat die Wanderungsbewegung beider Geschlechter einen Einfluss auf den Genfluss zwischen und eine Verminderung von Inzest auf einzelnen Kopjes. Nach genetischen Analysen ist die Variabilität innerhalb der Kolonie gering. Die meisten Tiere sind zwar untereinander verwandt, jedoch kommen bei den reproduktionsfähigen Weibchen auch familienfremde Individuen vor. Neubesiedlungen von Kopjes gehen möglicherweise von jungen Männchen aus, die dann Weibchen anlocken und so neue Kolonien gründen.[20][2][4][5]
Auf den Kopjes nutzen die Tiere Felshohlräume als Verstecke, selten bestehen sie auch aus Erdlöchern. Damit eine Kolonie entsteht sind Kopjes mit einem komplexeren Hohlraumsystem erforderlich, singuläre Unterschlüpfe werden nur selten aufgesucht. Diese Hohlräume sollten wenigstens eine Grundfläche von 1 m² aufweisen, um mehreren Individuen Platz zu bieten. Damit ein Tier kriechend hinein und heraus gelangen kann, müssen die Zugänge mindestens 11 cm hoch sein. Hohlräume mit zu hohen Eingängen meidet der Buschschliefer aber meist aus Schutz vor Fressfeinden. Die Eingänge getrennter Hohlräumen sind mit oberirdischen Pfaden verbunden.[21] Die Unterschlüpfe bieten eine mehr oder weniger konstante Temperatur von 17 bis 25 °C und Luftfeuchtigkeit von 32 bis 40 %, zusätzlich schützen sie vor Feuer.[18] Schlaf nimmt etwa 5,7 Stunden eines 24-Stunden-Tages ein.[22] Die Ruheperioden verbringen die Tiere innerhalb oder außerhalb der Verstecke. Sie liegen dann entweder dicht aneinander gekauert oder teilweise übereinander gehäuft, wobei sich dann die Jungtiere oben befinden. Diese Formierungen können teilweise am frühen Morgen beobachtet werden, wenn die Tiere ihre Verstecke verlassen und das erste Sonnenlicht über die Felsen streift. Sobald der gesamte Kopje beleuchtet ist, bricht die enge Formierung in kleinere Gruppen auf. Tagsüber badet der Baumschliefer auch in der Sonne, die einzelnen Individuen berühren sich dann nicht, sondern liegen nebeneinander entweder parallel gegenständig oder gleichgerichtet in einer leicht radialen Position, bei der die Köpfe dann etwas weiter voneinander entfernt sind als der Hinterleib. Die Hinterbeine liegen abgewinkelt nach hinten gestreckt mit den Sohlen nach oben. Beide Verhaltensweisen, das gehäufte Liegen und das Sonnenbaden, beeinflussen die Körpertemperatur.[23] Diese ist beim Buschschliefer instabil. In Ruheperioden beträgt sie durchschnittlich 36,4 °C. Sie ist aber teilweise abhängig von der Außentemperatur und kann um 7 bis 8 °C variieren. Die thermoneutrale Zone liegt bei 24 bis 35 °C. Überhitzung bei hohen Außentemperaturen begegnet der Buschschliefer mit einem hohen Wasserverlust, der vor allem über die Füße und die Nase erfolgt. Verbunden ist die schwankende Körpertemperatur mit einer niedrigen Stoffwechselrate und der Befähigung Urin zu konzentrieren.[24][25][2][4][5]
Innerhalb der Kolonie dient das gehäufte Liegen mit irregulären Positionen der einzelnen Individuen zueinander und das gegenständige oder gleichgerichtete Nebeneinanderruhen einer Aggressionsvermeidung.[23] Der Buschschliefer verteidigt sein Territorium aktiv gegen das Eindringen von Artgenossen. Drohgebärden und aggressives Verhalten zeichnen sich vor allem bei Männchen durch das Heben von Kopf und Schulter, eine Öffnung des Mauls und Präsentation der eckzahnartigen Schneidezähne sowie ein Aufrichten der Haare des hellen Rückenflecks aus. Letzteres fungiert auch als Warnsignal gegenüber anderen Tierarten oder erfolgt bei Erregung. Bei Attacken schnappen und beißen die Tiere einander. Darüber hinaus hat die Drüse des Rückens auch eine weitere kommunikative Funktion, indem sie Duftstoffe absondert, die etwa bei der Paarung eine Rolle spielen.[26] Territoriale Männchen geben laute Dominanzrufe ab. Diese zeigen nur wenige jahreszeitliche Schwankungen, werden aber zur Fortpflanzungsphase intensiver. Die Rufe sind schrill und lang, sie dauern zusammenhängend etwa 5 Minuten, ein einzelner Ruf hält rund 1,5 Sekunden an. In der Regel stimuliert ein rufendes Männchen andere Männchen in der Umgebung.[6][19] Daneben gibt es noch ein weites Repertoire an weiteren Rufen, die bei Gefahr oder bei Kontakten ausgestoßen werden und teilweise auch von Klippschliefern erkannt werden. Zum Komfortverhalten zählt das Kratzen mit der Kralle des Hinterfußes, mit der die Tiere nahezu alle Körperteile erreichen. Beim Putzen werden zudem die unteren, teils kammartigen Schneidezähne eingesetzt. Der Buschschliefer badet auch in Sand, häufig mit abgespreiztem Fell, was wohl bei der Entfernung von Ektoparasiten behilflich ist.[2][4][5]
Der Buschschliefer nutzt regelmäßig Latrinen zur Defäkation. Das konzentrierte Urin enthält hauptsächlich Salz, in Verbindung mit dem Kot entsteht eine dunkle kristalline und äußerlich pechartige Substanz, die mit der Zeit aufhellt. Diese bedeckt flächig die Latrinen. Sie wird als „Hyraceum“ bezeichnet und findet Einsatz in der lokalen Medizin.[18][2][4][5]
Ernährung
.jpg.webp)
Der Buschschliefer ernährt sich hauptsächlich von weicher Pflanzenkost (browsing). Er verbringt mehr als 80 % seiner Zeit der Nahrungsaufnahme mit dem Fressen von Zweigen, Rinde, Blättern, Knospen, Blüten und Früchten. Ihre Nahrung beziehen die Tiere überwiegend von Bäumen und Büschen, seltener von der Bodenvegetation. Während der Trockenzeit stammt rund 81 % der Nahrung von Gehölzpflanzen, in der Regenzeit etwa 92 %. Nur äußerst selten konsumiert der Buschschliefer auch Gräser, was dann auch meist während der Regenzeit geschieht, wenn die Pflanzen frisch sind. Aus der Serengeti wurden 64 verschiedene Pflanzenarten dokumentiert, die der Buschschliefer verzehrt, davon bilden jedoch zwei bis elf Pflanzen rund 90 % der Hauptnahrung eines Tieres. Bedeutende Nahrungspflanzen stellen Vertreter von Vachellia, Allophylus, Cordia, Grewia, Iboza und Maerua sowie Hibiskus und Feigen dar. Die Zusammensetzung der Nahrung variiert hier zwischen den einzelnen Familiengruppen und ist abhängig vom jeweiligen Standort und der Bevorzugung.[18][27] Im Matobo-Nationalpark ernähren sich die Tiere häufig von Arten von Flueggia, Strynchia, Kirkia, Croton und Mundulea, während Jungtiere auch Teile von Rhus und Commiphora nicht verschmähen. Der Flüssigkeitsbedarf wird überwiegend mit der Nahrung gedeckt, gelegentlich trinkt der Buschschliefer aber auch frisches Wasser.[2][3][4][5]
In der Regel frisst der Buschschliefer morgens zwischen 8.00 und 11.00 Uhr sowie abends zwischen 15.00 und 19.00 Uhr, gelegentlich auch nachts. Die Gruppe entfernt sich dabei bis zu 50 m vom Zentrum der Kolonie. Seltener gehen einzelne Individuen fressen, die sich dann häufig in einer Distanz nicht weiter als 20 m aufhalten. Die einzelnen Fressphasen dauern zwischen 20 und 35 Minuten. Der Buschschliefer klettert behände im Geäst und balanciert auf dünnen Zweigen, um auch die entferntesten Blätter zu erreichen.[28][18] Häufig beobachtet ein einzelnes Individuum als „Wächter“ die Gruppe und gibt Alarmsignale bei aufziehender Gefahr von sich. Vor allem in der Trockenzeit fressen sowohl der Buschschliefer als auch der Klippschliefer die gleiche Nahrungsressource. In Gebieten mit sympatrischem Vorkommen nutzen beide Arten dann gleichzeitig dieselben Bäume und Büsche.[18][27][2][4][5]
Fortpflanzung

Die Paarungszeit ist möglicherweise in äquatornahen Gebieten zweigeteilt, ansonsten stärker jahreszeitlich abhängig, in höheren Regionen kann die Sonneneinstrahlung eine Rolle spielen. Der Östrus der Weibchen tritt wohl zweimal jährlich auf und hält etwa 1 bis 3 Tage an, möglicherweise wird er mehrfach innerhalb von vier Wochen wiederholt.[19] Bei Männchen schwellen die Hoden in dieser Phase extrem an.[29] Es drückt seinen Paarungswillen mit einem schrillen Schrei aus, das Weibchen stellt die Haare des Rückenflecks auf. Danach vollführen beide einen Tanz, bei dem das Männchen an den weiblichen Geschlechtsorganen schnüffelt und sein Kinn auf den Rumpf legt, bevor es die Partnerin besteigt. Der Geschlechtsakt dauert etwa drei bis fünf Minuten und wird nach bis zu drei Stunden wiederholt.[2][4][5]
Die Tragzeit währt etwa 26 bis 30 Wochen oder rund 7,5 Monate, was ausgesprochen lang ist für ein Tier geringer Größe.[30] In der Serengeti wurden zwei Wurfzeiten pro Jahr registriert, eine zwischen Mai und Juli, die zweite zwischen Dezember und Januar. In beiden Fällen liegt der Zeitraum kurz nach dem Ende der Regenzeit. Die Weibchen jeder Kolonie bekommen einmal im Jahr Junge, so dass die Geburt des Nachwuchses in einen der beiden Zeiträume fällt. Die durchschnittliche Anzahl der Jungen pro Wurf liegt bei 1,6.[12] Im Matobo-Nationalpark wurden Geburten vor allem im März beobachtet, etwa zwei Monate nach dem Höhepunkt der Regenzeit. Hier kamen durchschnittlich 2,1 Jungen pro Wurf zur Welt. Insgesamt ist die Geburt des Nachwuchses stark synchronisiert, so dass etwa die Hälfte aller Weibchen einer Population beteiligt sind.[31] Das Geburtsgewicht der Neugeborenen variiert von 220 bis 230 g.[3] Die Jungen sind weit entwickelt und vollständig behaart. Häufig finden gemeinsame Aktivitäten der Jungtiere statt, diese umfassen kneifen oder beißen, klettern, verfolgen, kämpfen oder drücken. An Kopjes, wo Busch- und Klippschliefer zusammen auftreten, sind auch gemischte Jungengruppen zu beobachten.[32] Die Muttertiere säugen ausschließlich ihren eigenen Nachwuchs, die Jungen suchen dafür eine bevorzugte Zitze auf. Der Entwöhnung setzt im fünften Lebensmonat ein. Die sexuelle Reife wird mit 16 bis 17 Monaten erreicht. Junge männliche Tiere verlassen dann in der Regel ihre Heimatkolonie, junge weibliche werden in die Familiengruppe integriert. An zwei Kopjes der Serengeti betrug die durchschnittliche Lebensdauer der Weibchen 50,6 beziehungsweise 35 Monate, die der Männchen 19,1 beziehungsweise 22,6 Monate.[12] Ein einzelnes Weibchen wurde dabei über 11 Jahre alt,[13] was in etwa auch dem bekannten Höchstalter in menschlicher Gefangenschaft entspricht.[2][4][5]
Fressfeinde und Parasiten
Einen sehr starken Einfluss auf die örtlichen Bestände an Schliefern hat der Klippenadler. Bei Analysen an Gewöllresten aus der Serengeti entfielen von 224 Beuteresten des Greifvogels 123 auf den Buschschliefer.[12] Ähnlich verhält es sich im Matobo-Nationalpark, hier bezieht der Kaffernadler mehr als 90 % seiner Beute vom Busch- und vom Klippschliefer. Dabei stammen nach Analysen von 184 beobachteten Nestern mit rund 1550 Beuteresten mehr als 830 vom Buschschliefer. Insgesamt 86 % der erlegten Buschschliefer repräsentieren ausgewachsene Tiere.[33][34][35] Dadurch ist der Buschschliefer im Matobo-Nationalpark einem hohen Jagddruck ausgesetzt; es wird geschätzt, dass unter den Jungtieren 52 bis 61 % das erste Lebensjahr nicht überstehen.[14] Die offensichtliche Abhängigkeit des Kaffernadlers von den Schliefern zeigt sich auch darin, dass sich beispielsweise die Brutperiode bei den Greifvögeln verschiebt, wenn ihre Beutetiere in trockenen Jahren nur eine geringe Nachwuchsrate aufweisen.[16] Unter den Säugetieren stellt der Leopard einen der bedeutendsten Fressfeinde des Buschschliefers dar. Ebenfalls im Matobo-Nationalpark bildet er gemeinsam mit dem Klippschliefer 32 bis 50 % der Nahrung der Raubkatze.[36][37] Daneben gibt es noch zahlreiche weitere Beutegreifer, so verschiedene Raubtiere, Schlangen und Echsen. Der Buschschliefer reagiert meist mit Flucht in die Verstecke auf aufziehende Gefahr.[18][12][2][4][5]
Zu den äußeren Parasiten gehören Zecken wie Rhipicephalus und Haemaphysalis, Flöhe wie Procaviopsylla und Läuse wie Prolignognathus. Des Weiteren sind als innere Parasiten vor allem Fadenwürmer belegt, etwa Crossophorus, der fast ausschließlich im vorderen Blinddarm nistet. Der Buschschliefer ist außerdem Träger von Leishmanien und kann an Lungenentzündung und Tuberkulose erkranken.[12][2][4][5]
Systematik
Innere Systematik der rezenten Schliefer nach Maswanganye et al. 2017[38]
|
Der Buschschliefer ist eine Art aus der Gattung Heterohyrax und deren einziger Vertreter. Art und Gattung gehören zur Familie der Schliefer (Procaviidae) und zu der im Deutschen gleichnamigen Ordnung der Schliefer (Hyracoidea). Die Ordnung besteht heute aus zwei weiteren Gattungen. In ihrer stammesgeschichtlichen Vergangenheit, vor allem im Paläogen und im frühen Neogen, stellte sie aber eine sehr formen- und variantenreiche Gruppe dar. Sie schloss damals zahlreiche Angehörige ein, darunter sowohl kleine als auch riesige Tiere. Diese zeigten die unterschiedlichsten ökologischen Anpassungen, die Verbreitung der damaligen Schliefer reichte über weite Teile Eurasiens und Afrikas. Die heutigen Schliefer sind auf meerschweinchenartige Formen beschränkt und kommen mit einer Ausnahme nur auf dem afrikanischen Kontinent vor. Der Buschschliefer ist ausschließlich in Afrikas verbreitet, in seiner sozialen Lebensweise und seiner Tagesaktivität ähnelt er dem Klippschliefer. Er weicht von diesem aber in einzelnen anatomischen Merkmalen ab, auch gibt es bestimmte serologische Unterschiede wie in der Mobilität der Amylasen.[39] Zu den Baumschliefern bestehen größere Unterschiede im Skelettbau wie auch in der Lebensweise.[40][41]
Gegenwärtig werden bis zu 24 Unterarten des Buschschliefers unterschieden:[4][5]
- H. b. albipes Hollister, 1922; Kenia
- H. b. bakeri (J. E. Gray, 1874); Uganda
- H. b. bocagei (J. E. Gray, 1869); Angola
- H. b. brucei (J. E. Gray, 1868); Äthiopien
- H. b. chapini (Hatt, 1933); Demokratische Republik Kongo
- H. b. dieseneri Brauer, 1917; Tansania
- H. b. frommi (Brauer, 1913); Tansania
- H. b. granti (Wroughton, 1910); Südafrika
- H. b. hindei (Wroughton, 1910); Kenia
- H. b. hoogstraali Setzer, 1956; Sudan
- H. b. kempi (Thomas, 1910); Kenia
- H. b. lademanni Brauer, 1917; Tansania
- H. b. manningi (Wroughton, 1910); Malawi
- H. b. mossambicus (Peters, 1870); Mosambik
- H. b. muenzneri (Brauer, 1913); Tansania
- H. b. princeps (Thomas, 1910); Äthiopien
- H. b. prittwitzi Brauer, 1917; Tansania
- H. b. pumilus (Thomas, 1910); Somaliland
- H. b. ruddi (Wroughton, 1910); Mosambik
- H. b. rudolfi (Thomas, 1910); Äthiopien
- H. b. somalicus (Thomas, 1892); Somaliland
- H. b. ssongeae Brauer, 1917; Tansania
- H. b. thomasi (Neumann, 1901); Sudan
- H. b. victorianjansae Brauer, 1917; Tansania
Eine weitere Unterart wurde mit H. b. antineae in Algerien angenommen.[42][2] Beschrieben wurde diese 1932 von Henri Heim de Balsac und Max Bégouen, sie steht aber neueren Untersuchungen zufolge dem Klippschliefer näher.[10] Es ist zurzeit unklar, ob die hohe Anzahl an Unterarten tatsächlich Bestand hat, eine Revision der Gattung wird als notwendig erachtet. Anhand der Fellfärbung lässt sich jedoch eine starke Variabilität über das gesamte Verbreitungsgebiet belegen.[9] Auch bestehen weitere Unterschiede. So weist die Form H. b. chapini als einziger Vertreter der Buschschliefer nur zwei Zitzenpaare auf, die sich in der Leistengegend befinden.[43][44] Andererseits ist für H. b. princeps und H. b. thomasi aus dem nordöstlichen, für H. b. lademanni aus dem östlichen sowie für H. b. mossambicus aus dem südlichen Abschnitt des Verbreitungsgebietes eine stärker baumbewohnende Lebensweise dokumentiert.[7][45]
Einzelne Formen wurden in der zweiten Hälfte des 20. Jahrhunderts als eigenständige Arten anerkannt, hierzu gehören unter anderem Heterohyrax chapini und zusätzlich auch Heterohyrax antineae, allerdings sehen die meisten modernen Systematiken den Buschschliefer als einzelne Art und die Gattung somit als monotypisch an.[2][46][4][5] Vorläufige molekulargenetische Studien sprechen demgegenüber dafür, dass sich die Gattung wohl aus mehreren kryptischen Arten zusammensetzt. So sind die beiden Formen H. b. ruddi und H. b. granti aus dem südlichen Afrika nicht näher miteinander verwandt, sondern erstere steht H. b. hindei aus dem östlichen Afrika näher.[47] Es sind bisher noch keine umfassenden genetischen Analysen vorgelegt worden. Allgemein lautet die Chromosomennummer 2n=54. Der Chromosomensatz besteht aus 20 acrozentrischen, 2 telozentrischen, 2 submetazentrischen und 2 metazentrischen Autosomenpaaren. Das X-Chromosom repräsentiert das größte submetazentrische Chromosom und enthält 5,2 % des weiblichen Genoms, das Y-Chromosom ist klein und acrozentrisch mit 1,4 % des Genoms.[48][4][5]
Neben dem heutigen Buschschliefer wird eine fossile Art unterschieden:[49]
- Heterohyrax auricampensis Rasmussen, Pickford, Mein, Senut & Conroy, 1996
Forschungsgeschichte

Die Systematikgeschichte des Buschschliefers ist komplex. Die heute gebräuchliche wissenschaftliche Bezeichnung Heterohyrax brucei geht auf John Edward Gray aus dem Jahr 1868 zurück. Gray führte aber Gattungs- und Artnahmen unabhängig voneinander ein. Er kreierte die Gattung Heterohyrax aufgrund vergleichbarer Schädelmerkmale als Untergattung von Dendrohyrax, setzte beide jedoch durch die Ausprägung des Postorbitalbogens voneinander ab, der bei ersterer unvollständig, bei letzterer vollständig ist. Als Typusart bestimmte er vorläufig Heterohyrax blainvillii.[50] Der Name bezieht sich auf Henri Marie Ducrotay de Blainville, der zuvor im Jahr 1863 im dritten Band seines Werkes Ostéographie ou description iconographique comparée du squelette et du système dentaire des mammifères récents et fossiles die Schliefer umfänglich abgehandelt und die Schädel- und Skelettanatomie beschrieben hatte.[51] Der Auffassung, Heterohyrax sei eine Unterart von Dendrohyrax, wurde teilweise bis weit in das 20. Jahrhundert gefolgt. Als vereinendes Charakteristikum galten die niederkronigen Backenzähne.[18][52] In der Regel ist aber Heterohyrax heute als eigenständige Gattung innerhalb der Schliefer anerkannt, da nicht nur bedeutende Unterschiede in der Anatomie, sondern auch Abweichungen im Verhalten und in der Lebensweise der Tiere vorliegen.[4][40]
Im gleichen Aufsatz stellte Gray die Art Hyrax brucei vor, die er in die Nähe des Klippschliefers verwies, sie aber von diesem durch ein weicheres Fell unterschied. Als Typusregion gab Gray Abessinien an, also das heutige Äthiopien und Eritrea.[50] Der Name bezieht sich auf James Bruce, einem schottischen Naturforscher, der im Zeitraum von 1770 bis 1772 in Abessinien verweilte und dabei auch die Quellen des Nils suchte. In seinen Reiseerzählungen Travels to discover the sources of the Nile aus dem Jahr 1790 beschäftigte er sich eingehend mit einem Tier, für das er die lokale Bezeichnung ashkoko nutzte. Seine Beschreibung des ashkoko bezieht sich mit dem dunklen Rücken und dem hellen Bauch, dem gelben Rückenfleck, dem fehlenden Schwanz und der geselligen Lebensweise eindeutig auf den Buschschliefer.[53] Nur zwei Jahre später widmete sich Johann Christian von Schreber den Schliefern ausführlich. Er verwies dabei auf Bruces umfassende Beschreibungen des Buschschliefers und benannte die Art Hyrax syriacus, allerdings unter Einbeziehung weiterer Berichte, die Tiere aus Vorderasien behandelten. Er trennte seine neue Form von den damals bekannten südafrikanischen Klippschliefern ab, die er unter der Bezeichnung Hyrax capensis führte.[54] In seiner Erstbeschreibung zu Hyrax brucei 1868 wies Gray bereits auf den Umstand hin, ebenso wie auf die Tatsache, dass die abessinische Form nicht in Vorderasien auftritt. Er beschrieb diese daher als Hyrax brucei neu. Die Tiere aus Vorderasien führte er dagegen unter Hyrax sinaiticus.[50] Im Jahr 1892 unterstützte Oldfield Thomas diese Trennung, er wies Procavia brucei als Typusform von Heterohyrax aus und stellte die vorderasiatischen Schliefer zu Procavia syriaca. Als Begründung für letzteren Schritt gab Thomas an, das Exemplar, das Bruce seiner Beschreibung zu Grunde legte, stamme tatsächlich aus dem Libanon.[55]
Vor allem in der Wende vom 19. zum 20. Jahrhundert wurden zahlreiche neue Schlieferformen beschrieben. Die Verwendung der Gattung Heterohyrax war dabei sehr variabel, vielmehr wurden die Tiere in einer „Heterohyrax-Gruppe“ zusammengefasst. Allein Oldfield Thomas führte 1892 und 1910 fünf Vertreter des Buschschliefers ein,[55][56] ebenfalls im Jahr 1910 etablierte Robert Charles Wroughton vier Formen[57] und 1913 sowie 1917 benannte August Brauer ein Dutzend weitere neue Angehörige.[58][59] In einem unveröffentlichten Manuskript erweiterte Brauer dann die Gattung Heterohyrax auf 11 Arten mit bis zu 28 Unterarten. Im Jahr 1934 überarbeitete Herbert Hahn Brauers Entwurf und legte eine umfassende Revision der Schliefer vor. Darin fasste er alle Buschschliefer zu einer Art zusammen und bezeichnete diese mit Heterohyrax syriacus. Er widersprach damit der Auffassung von Thomas. Hahn argumentierte, dass Schreber in seiner Beschreibung von Hyrax syriacus unter Berufung auf Bruce ausdrücklich die Tiere aus Abessinien meinte. Diese stimmten äußerlich mit der scharf gegen den dunklen Rücken abgesetzten hellen Bauchfärbung nicht mit dem vorderasiatischen Klippschliefer überein. Als älterer Name für den Buschschliefer wäre demnach Schrebers Artbezeichnung die richtige.[7] Die Zusammenfassung zu einer Art wurde mit einzelnen Ausnahmen weitgehend akzeptiert, auch übernahmen einzelne Wissenschaftler die Artbezeichnung Heretohyrax syriacus.[60] Entgegen Hahn meinten aber John Ellerman und Kollegen Anfang der 1950er Jahre, Heterohyrax syriacus sei nicht der korrekte Artname. Schreber basierte seine Art zwar auf Bruces Beschreibung des Buschschliefers, fasste in diese aber auch den vorderasiatischen Klippschliefer mit ein. Schrebers Hyrax syriacus kann daher als Hybrid aus zwei Arten angesehen werden. Unter Berücksichtigung der Prioritätsregel ist Heterohyrax brucei somit der älteste verfügbare Name für den Buschschliefer.[61][62] Er findet seitdem mehrheitlich Verwendung.[2][3][4][5]
Stammesgeschichte
Die ältesten Nachweise procaviider Schliefer stammen aus dem Miozän. Funde der Gattung Heterohyrax sind aus den Otavibergen im nördlichen Namibia belegt. Es handelt sich um Unterkiefer- und Schädelfunde, die in einer Höhlenbrekzie am Berg Aukas lagerten und ein ausgewachsenes und zwei jüngere Tiere repräsentieren. Sie werden zur Art Heterohyrax auricampensis gestellt, welche 1996 von D. Tab Rasmussen unter Beteiligung von Martin Pickford, Brigitte Senut und weiteren Forschern wissenschaftlich eingeführt worden war. Die Tiere übertrafen den heutigen Buschschliefer in ihren Körperausmaßen und entsprachen in diesen etwa dem Klippschliefer. Die Zähne sind stark niederkronig und ähneln denen der Baumschliefer, der Radius ist aber deutlich abgeplattet und eignete sich daher nicht für starke Rotationsbewegungen, die für das Klettern benötigt werden. Die Funde sind etwa 10 Millionen Jahre alt, was dem beginnenden Oberen Miozän entspricht. Weitere Funde kamen in Omo in Äthiopien zum Vorschein. Sie gehören dem Pliozän an, sind aber bisher unbeschrieben.[49][63]
Bedrohung und Schutz
Der Buschschliefer wird von der IUCN in der Kategorie „nicht gefährdet“ (least concern) gelistet. Größere Bedrohungen sind nicht bekannt. Lokal werden die Tiere gejagt und die Felle zu Decken verarbeitet, was zu einem erheblichen Ausdünnen der örtlichen Bestände führen kann. Insgesamt ist die Art aber weit verbreitet, ein drastischer Rückgang der Gesamtpopulation wird nicht erwartet. Sie kommt in zahlreichen Schutzgebieten vor.[64]
Literatur
- Ronald E. Barry und Hendrik N. Hoeck: Heterohyrax brucei Bush Hyrax (Yellow-spotted Hyrax). In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 161–165.
- Ronald E. Barry und Jeheskel Shoshani: Heterohyrax brucei. Mammalian Species 645, 2000, S. 1–7.
- Angela Gaylard: Heterohyrax brucei Gray, 1868 – Bush hyrax. In: John D. Skinner und Christian T. Chimimba (Hrsg.): The Mammals of the Southern African Subregion. Cambridge University Press, 2005, S. 46–48.
- Hendrik Hoeck: Family Procaviidae (Hyraxes). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 2: Hooved Mammals. Lynx Edicions, Barcelona 2011, ISBN 978-84-96553-77-4, S. 28–47 (S. 45–46).
- Ronald M. Nowak: Walker’s Mammals of the World. Johns Hopkins University Press, 1999, ISBN 0-8018-5789-9.
Videos
- Hendrik N. Hoeck: Heterohyrax brucei (Procaviidae) – Fortbewegung im Geäst und Nahrungsaufnahme (Freilandaufnahmen). Film E-2266 des Instituts für den Wissenschaftlichen Film, Publikation von H. N. Hoeck, Publikationen zu Wissenschaftlichen Filmen, Sektion Biologie, Serie 10, Nr. 60/E 2266, 1977, Begleitheft 11 Seiten.
- Hendrik N. Hoeck: Heterohyrax brucei (Procaviidae) – Spiel der Jungen miteinander und mit Jungen von Procavia johnstoni (Freilandaufnahmen). Film E-2473 des Instituts für den Wissenschaftlichen Film, Publikation von H. N. Hoeck, Publikationen zu Wissenschaftlichen Filmen, Sektion Biologie, Serie 11, Nr. 29/E 2473, 1978, Begleitheft 8 Seiten.
- Hendrik N. Hoeck: Heterohyrax brucei (Procaviidae) – Paarungsverhalten (Freilandaufnahmen). Film E-2474 des Instituts für den Wissenschaftlichen Film, Publikation von H. N. Hoeck, Publikationen zu Wissenschaftlichen Filmen, Sektion Biologie, Serie 11, Nr. 30/E 2474, 1978, Begleitheft 10 Seiten.
- Hendrik N. Hoeck: Ethologie von Busch- und Klippschliefer. Film D-1338 des Instituts für den Wissenschaftlichen Film, Publikation von H. N. Hoeck, Publikationen zu Wissenschaftlichen Filmen, Sektion Biologie, Serie 15, Nr. 31/D 1338, 1982, Begleitheft 24 Seiten.
- Hendrik N. Hoeck: Nahrungsökologie bei Busch- und Klippschliefer – Sympatrische Lebensweise. Film D-1371 des Instituts für den Wissenschaftlichen Film, Publikation von H. N. Hoeck, Publikationen zu Wissenschaftlichen Filmen, Sektion Biologie, Serie 15, Nr. 32/D 1371, 1982.
Einzelnachweise
- Ngoni Chiweshe: Dassie census in the Matobo Hills, Zimbabwe. Afrotherian Conservation 1, 2002, S. 6–7.
- Ronald E. Barry und Jeheskel Shoshani: Heterohyrax brucei. Mammalian Species 645, 2000, S. 1–7.
- Angela Gaylard: Heterohyrax brucei Gray, 1868 – Bush hyrax. In: John D. Skinner und Christian T. Chimimba (Hrsg.): The Mammals of the Southern African Subregion. Cambridge University Press, 2005, S. 46–48.
- Hendrik Hoeck: Family Procaviidae (Hyraxes). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 2: Hooved Mammals. Lynx Edicions, Barcelona 2011, ISBN 978-84-96553-77-4, S. 28–47 (S. 45–46).
- Ronald E. Barry und Hendrik N. Hoeck: Heterohyrax brucei Bush Hyrax (Yellow-spotted Hyrax). In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 161–165.
- Hendrik Hoeck: Systematics of the Hyracoidea: toward a clarification. In: Duane A. Schlitter (Hrsg.): Ecology and taxonomy of African small mammals. Bulletin of the Carnegie Museum of Natural History 6, 1978, S. 146–151.
- Herbert Hahn: Die Familie der Procaviidae. Zeitschrift für Säugetierkunde 9, 1934, S. 207–358 (biodiversitylibrary.org).
- Martin Pickford: Fossil hyraxes (Hyracoidea: Mammalia) from the Late Miocene and Plio-Pleistocene of Africa, and the phylogeny of the Procaviidae. Palaeontologia africana 41, 2005, S. 141–161.
- J. du P. Bothma: Recent Hyracoidea (Mammalia) of Southern Africa. Annals of the Transvaal Museum 25, 1967, S. 117–152.
- Michael Hoffmann, Hendrik N. Hoeck und Koenraad De Smet: Does the bush hyrax Heterohyrax brucei occur in North Africa? Afrotherian Conservation 6, 2008, S. 6–8.
- Peter Coals und Francis Gilbert: Notes on hyrax in South Sinai’s high mountains. Afrotherian Conservation 11, 2015, S. 5–8.
- H. N. Hoeck: Population dynamics, dispersal and genetic isolation in two species of hyrax (Heterohyrax brucei and Procavia johnstoni) on habitat islands in the Serengeti. Zeitschrift für Tierpsychologie 59, 1982, S. 177–210.
- H. N. Hoeck: Demography and competition in hyrax: a 17 years study. Oecologia 79, 1989, S. 353–360.
- Ronald E. Barry und Peter J. Mundy: Population dynamics of two species of hyraxes in the Matobo National Park, Zimbabwe. African Journal of Ecology 36, 1998, S. 221–236.
- R. E. Barry, N. Chiweshe und P. J. Mundy: Ten years of population data on two species of hyrax in the Matobo National Park, Zimbabwe. Afrotherian Conservation 2, 2003, S. 8–9.
- Ngoni Chiweshe: An update on the annual dassie census in Matobo Hills, Zimbabwe. Afrotherian Conservation 3, 2005, S. 7–8.
- Ronald E. Barry, Ngoni Chiweshe und Peter J. Mundy: Fluctuations in bush and rock Hyrax (Hyracoidea: Procaviidae) Abundances Over a 13-Year Period in the Matopos, Zimbabwe. African Journal of Wildlife Research 45 (1), 2015, S. 17–27.
- M. I. M. Turner und I. M. Watson: An introductory study on the ecology of hyrax (Dendrohyrax brucei and Procavia johnstoni). African Journal of Ecology 3 (1), 1965, S. 49–60.
- H. N. Hoeck, H. Klein und P. Hoeck: Flexible social organization in hyrax. Zeitschrift für Tierpsychologie 59, 1982, S. 265–298.
- G. Gerlach und H. N. Hoeck: Islands on the plains: metapopulation dynamics and female biased dispersal in hyraxes (Hyracoidea) in the Serengeti National Park. Molecular Ecology 10, 2001, S. 2307–2317.
- J. B. Sale: The habitat of the rock hyrax. Journal of the East African Natural History Society 25 (3), 1966, S. 205–214.
- Nadine Gravett, Adhil Bhagwandin, Oleg I. Lyamin, Jerome M. Siegel und Paul R. Manger: Sleep in the Rock Hyrax, Procavia capensis. Brain Behavior and Evolution 79, 2012, S. 155–169.
- J. B. Sale: The Behaviour of the Resting Rock Hyrax in Relation to its Environment. Zoologica Africana 5 (1), 1970, S. 87–99.
- C. R. Taylor und J. B. Sale: Temperature regulation in the hyrax. Comparative Biochemistry and Physiology 31 (6), 1969, S. 903–907.
- George A. Bartholomew und Michael Rainy: Regulation of Body Temperature in the Rock Hyrax, Heterohyrax brucei. Journal of Mammalogy 52 (1), 1971, S. 81–95.
- J. B. Sale: Unusual External Adaptations in the Rock Hyrax. Zoologica Africana 5 (1), 1970, S. 101–113.
- H. N. Hoeck: Differential Feeding Behaviour of the Sympatric Hyrax Procavia johnstoni and Heterohyrax brucei. Oecologia 22, 1975, S. 15–47.
- J. B. Sale: The feeding behaviour of rock hyraxes (genera Procavia and Heterohyrax) in Kenya. African Journal of Ecology 3 (1), 1965, S. 1–18.
- W. B. Neaves: The Annual Testicular Cycle in An Equatorial Colony of Lesser Rock Hyrax, Heterohyrax brucei. Proceedings of the Royal Society of London. Series B, Biological Sciences 206 (1163), 1979, S. 183–189.
- R. B. Heap, S. Gombe und J. B. Sale: Pregnancy in the hyrax and erythrocyte metabolism of pregesterone. Nature 257, 1975, S. 809–811.
- Ronald E. Barry: Synchronous parturition of Procavia capensis and Heterohyrax brucei during drought in Zimbabwe. South African Journal of Wildlife Research 24 (1/2), 1994, S. 1–5.
- T. M. Caro und R. M. Alawi: Comparative aspects of behavioural development in two species of free-living hyrax. Behaviour 95, 1984, S. 87–109.
- Ronald E. Barry und Lisa M. Barry: Species Composition and Age Structure of Remains of Hyraxes (Hyracoidea: Procaviidae) at Nests of Black Eagles. Journal of Mammalogy 77 (3), 1996, S. 702–707.
- Ronald E. Barry und Peter J. Mundy: Seasonal variation in the degree of heterospecific association of two syntopic hyraxes (Heterohyrax brucei and Procavia capensis) exhibiting synchronous parturition. Behavioral Ecology and Sociobiology 52, 2002, S. 177–181.
- N. Chiweshe: Black Eagles and hyraxes – the two flagship species in the conservation of wildlife in the Matobo Hills, Zimbabwe. Ostrich 78 (2), 2007, S. 381–386.
- J. H. Grobler und Vivian J. Wilson: Food of the leopard Panthera pardus (Linn.) in the Rhodes Matopo National Park, Rhodesia, as determined by faecal analysis. Arnoldia 35 (5), 1972, S. 1–9.
- Richard Mark Smith: Movement pattern and feeding behaviour of leopard in the Rhodes Matopo National Park, Rhodesia. Arnoldia 13 (8), 1977, S. 1–16.
- K. Amanda Maswanganye, Michael J. Cunningham, Nigel C. Bennett, Christian T. Chimimba und Paulette Bloomer: Life on the rocks: Multilocus phylogeography of rock hyrax (Procavia capensis) from southern Africa. Molecular Phylogenetics and Evolution 114, 2017, S. 49–62.
- Hans-Georg Scheil und Hendrik N. Hoeck: Elektrophoretische Untersuchungen der Serum-Amylasen (E. C. 3.2.1.1) des Klippschliefers Procavia johnstoni matschie Neumann 1900 und des Buschschliefers Heterohyrax brucei dieseneri (Vic. Njansae) Brauer 1917 – Hyracoidea: Mammalia. Zoologischer Anzeiger 215, 1985, S. 129–132.
- Jeheskel Shoshani, Paulette Bloomer und Erik R. Seiffert: Family Procaviidae Hyraxes. In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 150–151.
- Paulette Bloomer: Genus Heterohyrax Bush Hyraxes. In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 161.
- Henri Heim de Balsac und Max Bégouen: Faits noveaux concernant les Damans del l' Ahaggar. Bulletin du Museum National d'Histoire Naturelle 2, 1932, S. 478–483 (gallica.bnf.fr).
- Robert T. Hatt: An annotated catalogue of the Hyracoidea in the American Museum of Natural History, with a description of a new species from the Lower Congo. American Museum Novitates 594, 1933, S. 1–13.
- Robert T. Hatt: The hyraxes collected by the American Museum Congo expedition. Bulletin of the American Museum of Natural History 72, 1936, S. 117–139.
- Herbert Hahn: Von Baum-, Busch- und Klippschliefern. Wittenberg, 1959, S. 1–88.
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 (departments.bucknell.edu).
- Paulette Bloomer: Extant hyrax diversity is vastly underestimated. Afrotherian Conservation 7, 2009, S. 11–16.
- Paulette Prinsloo und T. J. Robinson: Comparative cytogenetics of the Hyracoidea: chromosomes of two Hyrax species from South Africa. Zeitschrift für Säugetierkunde 56, 1991, S. 331–338.
- D. Tab Rasmussen, Martin Pickford, Pierre Mein, Brigitte Senut und Glenn C. Conroy: Earliest Known Procaviid Hyracoid from the Late Miocene of Namibia. Journal of Mammalogy 77 (3), 1996, S. 745–754.
- John Edward Gray: Revision of the species of Hyrax, founded on the specimens in the British Museum. Annals and Magazine of Natural History 4 (1), 1868, S. 35–52 (biodiversitylibrary.org).
- Henri Marie Ducrotay de Blainville: Ostéographie ou description iconographique comparée du squelette et du système dentaire des mammifères récents et fossiles pour servir de base à la zoologie et à la géologie. Tome 3 : Quaternatés : des damans (Buffon). Paris, 1863, S. 1–45 (gallica.bnf.fr).
- J. Roche: Systematique du genera Procavia et desmans en general. Mammalia 36, 1972, S. 22–49.
- James Bruce: Travels to discover the source of the Nile: in the years 1768, 1769, 1770, 1771, 1772, and 1773. Vol. VI. Dublin, 1790, S. 1–286 (S. 171–180) (biodiversitylibrary.org); deutsch: Reisen zur Entdeckung der Quellen des Nils in den Jahren 1768. 1769. 1770. 1771 und 1772. Band V. Leipzig, 1791, S. 1–384 (S. 145–152) (digitale.bibliothek.uni-halle.de).
- Johann Christian Daniel Schreber: Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. Vierter Teil. Erlangen, 1792, S. 923–926 (biodiversitylibrary.org) und Tafelband 1 Tafel 240 (biodiversitylibrary.org).
- Oldfield Thomas: On the species of Hyracoidea. Proceedings of the Zoological Society of London 1892, S. 50–76 (biodiversitylibrary.org).
- Oldfield Thomas: Further new African mammals. Annals and Magazine of Natural History 8 (5), 1910, S. 191–202 (biodiversitylibrary.org).
- Robert Charles Wroughton: New African mammals of the genera Cricetomys and Procavia. Annals and Magazine of Natural History 8 (5), 1910, S. 106–110(biodiversitylibrary.org).
- A. Brauer: Weitere neue Procavia-Arten aus dem Kgl. Zoologischen Museum in Berlin. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1913, S. 125–141 (biodiversitylibrary.org).
- A. Brauer: Neue Procaviiden. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1917, S. 293–303 (biodiversitylibrary.org).
- G. H. Swynnerton und R. W. Hayman: A Checklist of the Land Mammals of the Tanganyika Territory and the Zanzibar Protectorate. Journal of the East African Natural History Society 20, 1950, S. 274–392 (S. 338).
- J. R. Ellerman und T. C. S. Morrison-Scott: Checklist of Palaearctic and Indian mammals 1758 to 1946. London, 1951, S. 1–810 (S. 335) (biodiversitylibrary.org).
- J. R. Ellerman, T. C. S. Morrison-Scott und R. W. Hayman: Southern African mammals 1758 to 1951: a reclassification. London, 1953, S. 1–363 (S. 157) (archive.org).
- D. Tab Rasmussen und Mercedes Gutiérrez: Hyracoidea. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, London, New York, 2010, S. 123–145.
- T. Butynski, H. Hoeck und Y. A. de Jong: Heterohyrax brucei. The IUCN Red List of Threatened Species 2015. e.T9997A21283287 (); zuletzt abgerufen am 28. Dezember 2017.
Weblinks
- Heterohyrax brucei in der Roten Liste gefährdeter Arten der IUCN 2015. Eingestellt von: T. Butynski, H. Hoeck und Y. A. de Jong, 2014. Abgerufen am 28. Dezember 2017.
- Hyrax vocalizations Rufe der Schliefer, aufgenommen im Rahmen des Eastern Africa Primate Diversity and Conservation Program, zuletzt abgerufen am 15. Dezember 2017