Welwitschie
Die Welwitschie (; Welwitschia mirabilis) ist die einzige Art der Gattung Welwitschia in der Familie der Welwitschiagewächse (Welwitschiaceae), die bereits vor 112 Millionen Jahren auf der Erde wuchsen.[1]
| Welwitschie | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Welwitschie in Namibia | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Familie | ||||||||||||
| Welwitschiaceae | ||||||||||||
| Markgr. | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Welwitschia | ||||||||||||
| Hook. f. | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Welwitschia mirabilis | ||||||||||||
| Hook. f. |
Die Pflanze gehört zur nacktsamigen Ordnung der Gnetales und wächst endemisch in der Wüste Namib im südlichen Afrika. Aufgrund ihres häufigen Vorkommens ist die Welwitschie unter anderem im Wappen Namibias, Wappen der Stadt Swakopmund und Wappen der Region Kunene abgebildet. Obwohl die Pflanze mehrere hundert Jahre alt wird, besitzt sie nur ein einziges Blattpaar.
Namen und botanische Geschichte

Der österreichische Arzt und Botaniker Friedrich Welwitsch entdeckte diese Pflanze im Jahre 1859 in der Nähe von Cabo Negro in Angola (15–16° S). In einem Brief an Sir William Jackson Hooker, den Leiter der Royal Botanic Gardens Kew, London, vom 16. August 1860 berichtete er erstmals über diese Pflanze. 1862 sandte er Joseph Dalton Hooker, ebenfalls in Kew, ein Exemplar, der die Pflanze 1863 wissenschaftlich beschrieb[2] und sie nach dem Entdecker benannte. Dieser hatte jedoch den Namen Tumboa nach der einheimischen Bezeichnung empfohlen. Hooker äußerte sich über die Pflanze folgendermaßen: It is out of the question the most wonderful plant ever brought to this country, and one of the ugliest.[3] („Dies ist ohne Frage die wunderbarste Pflanze, die je in dieses Land gebracht wurde, und eine der hässlichsten.“)
In Angola wird diese Pflanzenart n'tumbo genannt, was so viel wie „Stumpf“ bedeutet. Die Nama nennen sie ǃkharos oder auch khurub, die Damara nyanka. Die Herero nennen sie onyanga, was „Wüstenzwiebel“ bedeutet. Das Mark wurde früher – roh oder in heißer Asche gebacken – gegessen. Auf Afrikaans heißt sie tweeblaarkanniedood, was etwa „Zwei-Blatt-kann-nicht-sterben“ bedeutet.
Beschreibung und Ökologie
.jpg.webp)



Erscheinungsbild
Die ausdauernde Pflanze besitzt einen kurzen, rübenförmigen Stamm, der aus dem Hypokotyl hervorgeht, eine tiefreichende Pfahlwurzel und zwei Laubblätter, die die Keimblätter ersetzen.
Der Stamm ist verholzt und wird oberirdisch meist rund 50 Zentimeter hoch, maximal 1,50 Meter. Er erreicht einen Durchmesser von bis zu einem Meter und weist Jahresringe auf. Einzelexemplare besitzen einen Umfang von bis zu 8,7 Metern. Die Oberseite des Stammes ist eine konkave Scheibe, da der terminale Apex das Wachstum sehr früh einstellt. Nahe der Blattbasis entspringen die Blütenstände. Das Sekundärholz besitzt Tracheen, eigentlich ein typisches Merkmal der Angiospermen.
Junge Pflanzen sind am Naturstandort sehr selten zu finden. Keimlinge können sich nur nach – sehr seltenen – Extremniederschlägen etablieren, wodurch die Altersstruktur stark diskontinuierlich ist. Der Oberboden muss komplett durchfeuchtet sein, damit die Wurzeln der Jungpflanzen in größere Tiefen vordringen können. Die zwei Keimblätter können bis zu 1,5 Jahre photosynthetisch aktiv sein und sterben danach ab. Schon zuvor entwickelt sich das einzige Laubblattpaar.
Radiokohlenstoffdatierung hat für die untersuchten Pflanzen ein Alter von 500 bis 600 Jahren ergeben. Für die größten Exemplare der Art wird durch Extrapolation dieser Ergebnisse ein Alter von bis zu 2000 Jahren geschätzt.[4]
Ihr Wurzelwerk breitet sich unterirdisch über einen Radius von 15 Metern aus. Darüber hinaus hat die Pflanze eine Pfahlwurzel. Dass die Wurzeln den Grundwasserhorizont erreichen, ist wahrscheinlich, aber nicht gesichert, da sich die Wurzeln in einem harten, Calcit-verkitteten Kies verlieren. Die Wurzeln reichen bis in drei Meter Tiefe.
Blatt
Die beiden Laubblätter können über 2,5 Meter lang werden, manche Berichte sprechen von 6,2 Metern. Am Blattende sterben sie ab und verwittern, die ältesten lebenden Teile können jedoch 10 Jahre alt werden. Da das Hypokotyl sich mit zunehmendem Wachstum auffaltet, reißen die Blätter häufig auf und täuschen so mehrere Blätter vor. In der Umgebung des Brandbergs wurden jedoch Individuen gefunden, die tatsächlich zwei Blattpaare besitzen. Das tritt bei rund 5 % der Population auf. Die Blätter wachsen an einem basalen Meristem. Das Blattwachstum beträgt durchschnittlich 0,17 bis 0,83 Millimeter pro Tag. Die Jahreswerte variieren je nach Standort zwischen 40 und 409 Millimeter pro Jahr. Es besteht jedoch kein signifikanter Zusammenhang zwischen Blattwachstum und Niederschlagsmenge. Bedeutender dürfte die Wasserverfügbarkeit in den tieferen Bodenschichten sein. Die Leitbündel der Blätter können anastomosieren oder blind im Mesophyll enden. Das ist einzigartig unter den Gymnospermen.
Generative Merkmale
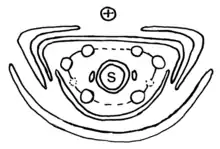
S = rudimentäre Samenanlage
Die Welwitschie ist zweihäusig getrenntgeschlechtig (diözisch), d. h., es gibt weibliche und männliche Pflanzen. Die Blüten befinden sich in zapfenartigen Blütenständen und sitzen in der Achsel von Deckschuppen.
Die Hülle der männlichen Blüten besteht aus zwei kreuzgegenständigen Brakteenpaaren. Die sechs Staubblätter stehen in einem Wirtel und sind an der Basis miteinander verwachsen. Jedes Staubblatt trägt drei miteinander verwachsene Pollensäcke. Die männlichen Blüten enthalten an der Spitze immer eine rudimentäre Samenanlage, die von einem ebenfalls rudimentären Brakteenpaar umgeben ist. Diese Samenanlage produziert Nektar, der zu rund 50 % aus Zucker besteht. Die sterilen weiblichen Samenanlagen und die Nektarproduktion können als evolutionär gescheiterter Versuch zur Bildung einer bisexuellen Blüte betrachtet werden.[5] Die Pollensäcke öffnen sich wie bei den anderen Vertretern der Gnetopsida mit einem Exothecium in Form von oft nur kurzen Schlitzen. Bei der Welwitschie wird der Pollen beim Austrocknen der Pollensäcke nach außen gepresst, was durch die Anordnung von Wandverstärkungen im Exothecium verursacht wird.
Der männliche Gametophyt besteht aus der spermatogenen Zelle und zwei weiteren Zellen. Die spermatogene Zelle teilt sich zu zwei Spermazellen. Die Befruchtung erfolgt über einen Pollenschlauch (Siphonogamie).
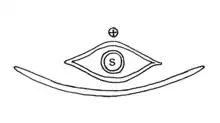
S = Samenanlage

Die weiblichen Blüten sind von zwei miteinander verwachsenen Brakteenpaaren umgeben. Bei der Samenreife wird das innere Brakteenpaar hart, das äußere bildet Flügel. Jede Blüte enthält eine aufrechte Samenanlage. Das Integument ist zu einer langen Mikropyle ausgezogen, an der ein Befruchtungstropfen, der auch als Nektar fungiert, ausgeschieden wird.
Der weibliche Gametophyt entsteht aus freien Kernteilungen aus allen vier aus der Meiose hervorgehenden Kernen und anschließender Zellwandbildung. Er kann bis zu eintausend Zellen umfassen. Es werden keine Archegonien gebildet, die Eizellen sind nicht von den übrigen Zellen des Archegoniums zu unterscheiden. Der Gametophyt wächst dem Pollenschlauch entgegen, indem er schlauchartige Strukturen ausbildet.
Bestäubung und Samenbildung
Die Bestäubung erfolgt durch Insekten, als Kandidaten werden Wanzen und Wespen diskutiert. Die Wanze Probergrothius angolensis[6] ernährt sich vom Nektar, Bestäubung wurde jedoch noch nicht eindeutig nachgewiesen. Die Blüte erfolgt vom Hochsommer bis Herbst, die Samen reifen im Frühjahr und werden durch Zerfall der Zapfen freigesetzt. Die Samen sind rund 3,5 × 2,5 Zentimeter groß und besitzen einen papierartigen Flügel. Die Ausbreitung erfolgt durch den Wind. Die Samen bleiben einige Jahre keimfähig und keimen nur nach stärkeren Regenfällen. Aus den rund 10.000 bis 20.000 Blüten pro Pflanze und Jahr entstehen insgesamt nur rund 20 bis 200 keimfähige Samen.
Vorkommen
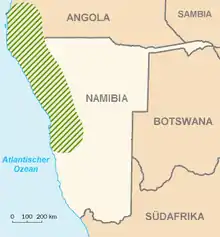
Die Art ist in der Wüste Namib beheimatet; ihr Areal umfasst Teile der Staaten Namibia und Angola[1]. Das Areal beginnt im Norden am Nicolau-Rivier nördlich von Namibe (Angola) und reicht rund 1200 km nach Süden bis zum Kuiseb-Rivier bei Gobabeb (Namibia). Sie ist allerdings nie direkt an der Küste zu finden. Der mittlere Jahresniederschlag an ihren Wuchsorten reicht von 10 mm in Küstennähe bis zu 250 mm in der Mopane-Savanne.
In manchen Gebieten der zentralen Namib ist die Welwitschie die dominante Pflanze. Die höchste Dichte erreicht sie in einer Entfernung von 50 bis 60 km von der Küste. Das Zentrum der Verbreitung ist die „Welwitschia-Vlakte“ (Welwitschia-Fläche) im Dreieck zwischen Khan und Swakop-Rivier. Die Gesamtindividuenzahl in diesem Gebiet wurde auf 5000 bis 6000 Pflanzen geschätzt. Dieses Gebiet ist Teil des Namib-Naukluft-Nationalparks. Die „Welwitschia-Vlakte“ steht seit 2002 auf der Tentativliste der UNESCO als Welterbe.
Systematik und botanische Geschichte
Die Gattung Welwitschia wurde 1863 mit der Erstbeschreibung von Welwitschia mirabilis durch Joseph Dalton Hooker in On Welwitschia, a new genus of Gnetaceae. In: Transactions of the Linnean Society of London, Volume 24, Seite 1–48 aufgestellt. Die Gattung Welwitschia ist monotypisch.
Nach Untersuchungen 2001 durch Leuenberger an kultivierten Exemplaren im Botanischen Gartens Berlin-Dahlem wurde von Welwitschia mirabilis eine zweite Unterart beschrieben. Leuenberger war der Ansicht, dass sie sich nicht nur in der Morphologie, sondern auch in ihrem Areal unterscheiden würden. Die Unterart Welwitschia mirabilis subsp. mirabilis soll nach Leuenberger nur in Angola vorkommen. Für die kultivierten Exemplare gab Leuenberger an: Die männlichen Zapfen sind glatt, bräunlich und ohne deutliche Wachsschicht. Die Blütenstandsachsen sind meist 5 bis 11 Zentimeter lang, die sekundären Achsen bis 2 Zentimeter. Die größten männlichen Zapfen sind 30 bis 45 Millimeter lang. Die Brakteenpaare überlappen sich rund 2 Millimeter. Die Brakteen sind zu mehr als drei Viertel der Länge verwachsen, der Rand der Brakteen ist glatt. – Die Unterart Welwitschia mirabilis subsp. namibiana Leuenberger soll nach Leuenberger nur in Namibia vorkommen. Für die kultivierten Exemplare gab Leuenberger an: Die männlichen Zapfen sind gefurcht, grünlich oder lachsfarben und mit deutlicher Wachsschicht. Die Blütenstandsachsen sind rund 7 bis 15 Zentimeter lang, die sekundären Achsen bis 7 Zentimeter. Die größten männlichen Zapfen sind 20 bis 30 Millimeter lang. Die Brakteenpaare überlappen sich rund 1 Millimeter. Die Brakteen sind zu ein bis zwei Drittel der Länge verwachsen, der Rand der Brakteen ist zerfranst.
All das konnte 2014 durch Jacobson et al. bei Felduntersuchungen nicht bestätigt werden. Es gibt keine Unterarten oder Varietäten.[7]
Fossile Welwitschiaceae
Fossilien von Arten der Ordnung Gnetales sind generell selten, wurden aber seit etwa 2000 etwas häufiger gefunden. Die meisten können entweder der Familie Ephedraceae zugeordnet werden oder sind als Stammgruppen-Vertreter mit unklarer Zugehörigkeit zu einer Familie anzusprechen.[8] Die einzigen überzeugenden Makrofossilien, die als Welwitschiaceae angesprochen werden können, stammen aus der weltberühmten Fossillagerstätte der Santana-Formation (bei Crato in Brasilien). Hier wurden exzellent erhaltene isolierte Blätter als Art Welwitschiophyllum brasiliense beschrieben. Ein Achsenfragment mit ansitzenden Blättern wurde einer neuen Art Welwitschiella austroamericana zugeschrieben. Isolierte zapfenartige Blütenstände wurden als Welwitschiostrobus murili, Funde eines Keimlings mit Keimblatt als Cratonia cotyledon beschrieben. Zusätzlich zu den Makrofossilien wurde auch fossiler Pollen mit Ähnlichkeit zu demjenigen von Welwitschia nachgewiesen, der mehreren Arten zugeordnet worden ist.[9] Die Datierung der Fossilien der Santana-Formation ist nicht ganz unumstritten, es wird ein Alter von etwa 112 bis 114 Millionen Jahre für die Funde abgeschätzt. Bei der Untersuchung der Welwitschiophyllum-Blätter konnte unerwarteterweise festgestellt werden, dass diese noch fossiles Harz enthielten, das chemisch nahe verwandt zu demjenigen aus Welwitschia-Blättern ist.[10] Die Fundorte in Namibia und Brasilien sind heute weit voneinander entfernt. Bei der Ablagerung der Fossilien im Aptium war aber der Südkontinent Gondwana noch nicht getrennt, so dass die Nachbarschaft, auf demselben Kontinent, viel näher war. Fossile Funde von Welwitschia selbst, oder andere Pflanzenfossilien der Familie aus Afrika sind bisher unbekannt.
Physiologie
Wasseraufnahme

Vielfach wird angeführt, die Welwitschie nehme ihren Bedarf an Wasser als Tau über die Blätter oder auch über ein oberflächennahes Feinwurzelsystem auf.
Die Welwitschie besitzt jedoch keine morphologischen Strukturen zur Aufnahme von Wasser über die Blätter. Die Blätter sind typisch xeromorph aufgebaut: Sie besitzen eine dicke Cuticula, die Spaltöffnungen sind eingesenkt, die Spalten sind besonders cutinisiert (Akkrustierung) und damit wasserabweisend. Zudem reicht der Nebel selten zu den Hauptvorkommen der Welwitschie und die Tau-Mengen reichen für die gemessene Pflanzenverdunstung nicht aus. Außerdem erreicht die Pflanze ihr morgendliches Wasserpotential bereits im Laufe der Nacht, lange bevor der Nebel einfällt. All das spricht für den Boden als alleinige Wasserquelle.
Versuche mit radioaktiv markiertem Wasser ergaben zwar, dass die Blätter Wasser aufnehmen können, jedoch nur sehr langsam und durch passive Diffusion, sodass diese keinen nennenswerten Beitrag zur Wasserversorgung leistet.
Allerdings dürfte die Aufnahme von Tau über oberflächennahe Feinwurzeln eine gewisse Rolle in der Wasserversorgung der Pflanze spielen. Schätzungen sprechen von einem Äquivalent von 50 mm Jahresniederschlag.
Inhaltsstoffe
Wie für Xerophyten vielfach typisch, enthalten die Blätter der Welwitschie recht hohe Konzentrationen anorganischer Ionen (in der Vakuole), wobei in den jungen Blattabschnitten Kalium- und Chlorid-Ionen dominieren, in älteren Abschnitten nimmt der Natrium-Anteil deutlich zu, da das phloemmobile Kalium in die jungen Abschnitte verlagert wird, so ändert sich das K/Na-Verhältnis von 5:1 im Meristem zu 1:4 an der Blattspitze. Zum osmotischen Ausgleich wird im Cytosol vor allem Prolin gebildet.
Die organischen Säuren, wie Äpfelsäure, Citronensäure, Isocitronensäure und Chinasäure, erreichen in Summe einen Gehalt von über 100 mmol/kg Trockengewicht.
Das Samenöl der Welwitschie enthält – wie auch das von Gnetum – Cyclopropene. Damit besitzen diese beiden Gattungen eine Sonderstellung gegenüber den anderen Gymnospermen, inklusive Ephedra, die alle Δ5-Fettsäuren enthalten. Mit ihrem Fettsäurespektrum steht Welwitschia den Malvales nahe, die aber zu den Angiospermen gehören.[11]
CAM-Stoffwechsel
.jpg.webp)
Die Diskussion, ob die Welwitschie eine CAM-Pflanze ist, ist nach dreißig Jahren noch nicht endgültig beendet, obwohl sich die Anzeichen dafür mehren.
Die δ-13C-Werte (zur Erklärung der Zahlenwerte siehe Isotopendiskriminierung) liegen in der „Welwitschia-Vlakte“ mit −17,5 ‰ bis −19,5 ‰ zwischen den Werten für C3- und C4-Pflanzen, was für CAM sprechen würde. In der Savanne erreicht Welwitschia jedoch Werte von −23,3 ‰ und entspricht eher einer C3-Pflanze. Auch eine hohe Aktivität der PEP-Carboxylase und Labor-Gaswechselmessungen sprechen für den CAM-Metabolismus. In einer neueren Publikation konnten von Willert et al. (2005) zwar auch im Gelände eine nächtliche CO2-Aufnahme zeigen, die jedoch nur 4 % der CO2-Aufnahme während 24 Stunden entsprach. Die höchsten Raten lagen bei 0,2 µmol/(m2·s). Diese nächtliche CO2-Aufnahme trat im Dezember und Januar auf, also im Sommer der Südhalbkugel. Diese Werte sprechen zumindest für ein sogenanntes CAM-cycling, d. h. die Refixierung des Atmungs-CO2. Die Blätter enthalten hohe Konzentrationen von Malat und Citrat. Bedingt durch die extrem hohe Variabilität im Blatt konnten von Willert et al. jedoch keine diurnale Änderung der Säurekonzentration zeigen. Das wäre jedoch der deutlichste Hinweis für den CAM-Stoffwechsel gewesen.
Gegen einen CAM-Stoffwechsel sprechen der xeromorphe Bau und der geringe Wassergehalt der Blätter, der ansonsten für CAM-Pflanzen typisch ist. Ebenso dagegen spricht, wie oben erwähnt, der fehlende Nachweis einer diurnalen Säureänderung.
Photorespiration, Photoinhibition
Die Welwitschie hat an ihrem natürlichen Standort auch eine hohe Photorespiration, die nahe bei 50 % der Gaswechselaktivität liegt. Die hohe Strahlung am Standort führt auch zu einer starken Photoinhibition, besonders in den Nachmittagsstunden. Das ist vor allem in den älteren Blattabschnitten ausgeprägt und führt oft so weit, dass bereits ab Mittag der CO2-Gaswechsel negativ wird.
Herbivorie
Die Welwitschie dient vielfach als Futterpflanze für Pflanzenfresser, u. a. für Oryx-Antilopen, Zebras und Nashörner. Oryx-Antilopen reißen die Blätter vollständig aus der Hypokotylgrube heraus, wobei sie allerdings das Meristem nicht zerstören; so kann die Pflanze innerhalb einiger Jahre wieder nachwachsen. Auch Sandstürme können die Blätter stark schädigen.
Schutz
Die Welwitschie ist zwar nicht bedroht, aufgrund ihrer Bekanntheit jedoch gesetzlich geschützt. Sie ist auch im Anhang II des Washingtoner Artenschutzabkommens (CITES) aufgeführt; lediglich Samen und Sämlinge dürfen gehandelt werden.
Welwitschien in Europa
Welwitschien gedeihen auch in Europa, sie sind allerdings wenig frosttolerant (bis −6 °C). Sie sind in vielen botanischen Gärten zu sehen, werden aber auch privat als Zierpflanzen gehalten. Die Anzucht erfolgt aus Samen, die in Spezialhandlungen erhältlich sind. Jungpflanzen sind, wie auch im natürlichen Habitat, empfindlich gegen Pilze, deren Sporen besonders an Samen von Wildpflanzen haften. Ältere Pflanzen sind relativ unempfindlich, der Standplatz sollte jedoch ihrem natürlichen Habitat möglichst ähnlich sein, bessere Wasser- und Nährstoffversorgung ausgenommen.
Sonstiges
Die Welwitschie ist auf dem Wappen Namibias abgebildet und die Namibische Rugby-Union-Nationalmannschaft leitet ihren Spitznamen Welwitschias von der Pflanze ab.
Siehe auch
Literatur
- Beat Ernst Leuenberger Welwitschia mirabilis (Welwitschiaceae), male cone characters and a new subspecies. In: Willdenowia, Volume 31, 2001, S. 357–381. ISSN 0511-9618doi:10.3372/wi.31.31206.
- Chris H. Bornman: Welwitschia – paradox of a parched paradise. C. Struik Publishers, Kapstadt-Johannesburg 1978, ISBN 0-86977-097-7.
- Patricia Craven, Christine Marais: Namib Flora. Von Swakopmund zur grossen Welwitschia über Goanikontes. Gamsberg Macmillan, Windhoek 2003, ISBN 0-86848-286-2.
- Joh R. Henschel: Welwitschia's World. Wordweaver Publishing House, Windhoek 2012, ISBN 978-99916-878-6-5.
- Robert J. Rodin: Distribution of Welwitschia mirabilis. In: American Journal of Botany. Columbus Ohio 40. 1953, ISSN 0002-9122, S. 280–285 (Repr. Johnson New York).
- Ernst van Jaarsvald, Uschi Pond: Welwitschia mirabilis. Kronenlose Herrscherin der Namib. Penrock Publications, Kapstadt 2013, ISBN 978-3-941602-78-6.
- Dieter J. von Willert, Nicole Armbrüster, Tobias Drees, Maik Zaborowski: CAM or not CAM – what is the answer? In: Functional Plant Biology. Band 32, 2005, ISSN 1445-4408, S. 389–395.
- Maik Veste: Welwitschia mirabilis. In: H. Walter, S.-W. Breckle: Ökologie der Erde. Band 2: Spezielle Ökologie der Tropischen und Subtropischen Zonen. Fischer, Stuttgart 1983, Elsevier, München 2004 (3. Aufl.), ISBN 3-8274-1540-3, S. 474–480.
- Peter Sitte, Elmar Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger. 35. Auflage. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X.
Weblinks
- Welwitschia im Botanischen Garten Berlin-Dahlem.
- Neue Zürcher Zeitung – Welwitschia mirabilis: mit Tricks uralt werden.
- Ausführliche englischsprachige Informationen inklusive Pflegeanleitung bei PlantzAfrica.
- Welwitschia auf Namibweb.com. (englisch)
- Christopher J. Earle: Welwitschia mirabilis Hooker 1863. In: The Gymnosperm Database. 28. Februar 2019, abgerufen am 9. April 2019 (englisch, die dort verwendeten Quellen sind älter als 2007).
Einzelnachweise
- Welwitschia mirabilis„Lebendes Fossil“ in der Namib-Wüste hat verschiedene Unterarten Universität Hamburg, aufgerufen am 6. Dezember 2021
- Joseph Dalton Hooker: On Welwitschia, a new Genus of Gnetaceae. In: Transactions of the Linnean Society of London. Band 24, London 1863, S. 1–48 (online).
- Conservation of Welwitschia mirabilis (Memento vom 30. November 2010 im Internet Archive), abgerufen 17. Februar 2006.
- Keith Coates Palgrave: Trees of southern Africa. 5. Auflage, Struik Publishers, Cape Town 1988. ISBN 0-86977-081-0. S. 62.
- E. Moyroud, M. Monniaux, E. Thévenon, R. Dumas, C. P. Scutt, M. W. Frohlich, F. Parcy: A link between LEAFY and B-gene homologues in Welwitschia mirabilis sheds light on ancestral mechanisms prefiguring floral development. In: New Phytologist. 2017, doi:10.1111/nph.14483.
- Welwitschia Bug. Abgerufen am 10. April 2020.
- Nicholas Jacobson, Peter Jacobson, Ernst van Jaarsveld, Kathryn Jacobson: Field evidence from Namibia does not support the designation of Angolan and Namibian subspecies of Welwitschia mirabilis Hook. In: Transactions of the Royal Society of South Africa, Volume 69, Issue 3, 2014, S. 179–186. doi:10.1080/0035919X.2014.950187.
- Yong Yang (2010): A Review on Gnetalean Megafossils: Problems and Perspectives. Taiwania 55(4): 346-354.
- David L. Dilcher, Mary E. Bernardes-de-Oleivera, Denise Pons, Terry A. Lott (2005): Welwitschiaceae from the Lower Cretaceous of northeastern Brazil. American Journal of Botany 92(8): 1294–1310. doi:10.3732/ajb.92.8.1294
- Emily A. Roberts, Leyla J. Seyfullah, Robert F. Loveridge, Paul Garside, David M. Martill (2020): Cretaceous gnetalean yields first preserved plant gum. Scientific Reports 10, Article number: 3401. doi:10.1038/s41598-020-60211-2
- K. Aitzetmüller, K. Vosmann: Cyclopropenoic fatty acids in gymnosperms: The seed oil of Welwitschia. In: Journal of the American Oil Chemists’ Society. Volume 75, Issue 12, 1998, S. 1761–1765 (doi:10.1007/s11746-998-0329-8).