Meristem
Meristem oder Bildungsgewebe bezeichnet einen Gewebetyp der Pflanzen, der aus undifferenzierten Zellen besteht und an dem Wachstum durch Zellteilung beteiligt sein kann.
Der Terminus Meristem wurde erstmals von Carl Wilhelm von Nägeli (1817–1891) in seinem Buch Beiträge zur Wissenschaftlichen Botanik in 1858 benutzt. Er leitet sich ab aus den altgriechischen Wörtern merisein (μερίζειν), das „teilen“ bedeutet und die grundlegende empirisch wahrnehmbare Eigenschaft der Meristeme beschreibt, und stemma (στέμμα) „Binde, Gebinde“.
Man findet primäre Meristeme sowohl im oberirdischen Teil der Pflanzen am äußersten Ende der Sprossspitzen (Sprossapikalmeristeme) und am äußersten Ende der Seitensprossen (Subapikalmeristeme) als auch im unterirdischen Teil, an den Wurzelspitzen (Wurzelapikalmeristeme). Die Zellen der Meristeme besitzen in der Regel dünne Zellwände mit wenig Cellulose. Sie sind, im Gegensatz zu ausdifferenzierten Zelltypen, theoretisch unbegrenzt teilungsfähig. Das Wachstum der Pflanzen beruht im Allgemeinen darauf, dass Spross- und Wurzelmeristeme an gegenüberliegenden Polen der Pflanze laufend Zellen durch Zellteilung an den Pflanzenkörper abgeben, die sich zu den unterschiedlichen Organen Spross, Wurzel und Blatt ausbilden.
Vergleich mit tierischen Stammzellen
Meristematische Zellen bei Pflanzen entsprechen den Stammzellen bei tierischen Organismen insofern, als sie mit ihnen die Eigenschaft gemeinsam haben, undifferenziert und der Ursprung aller spezialisierten Zelltypen zu sein. Die strikte Unterscheidung zwischen embryonalen Stammzellen und ausdifferenzierten adulten Zellen, die sich bei tierischen Organismen aufgrund der einzigartigen Plastizität der Stammzellen treffen lässt, ist bei Pflanzen allerdings weniger ausgeprägt. Selbst ausdifferenzierte Zellen sind hier in der Lage, sich unter bestimmten Bedingungen zu reembryonalisieren und neue, ganz andere Organe zu bilden. Anschaulich macht dies beispielsweise die Wurzelbildung bei Stecklingen.
Klassifikation der Meristeme
Primärmeristeme bilden sich aus Apikalmeristemen und sind für das Primärwachstum der Pflanze zuständig:
- Protoderm: bildet das äußere Abschlussgewebe, die Epidermis
- Grundmeristem: bildet das Markgewebe und das sekundäre Korkkambium
- Prokambium: bildet das Leitgewebe
Sekundärmeristeme (auch Lateralmeristeme) dienen dem Dickenwachstum:
- Kambium (als Kambiumring)
- Phellogen (auch Korkkambium): bildet das sekundäre Abschlussgewebe (Periderm) mit seinen drei Gewebeschichten
Tertiärmeristeme (auch Restmeristeme) sind größere meristematische Zellkomplexe in einer Umgebung mit bereits ausdifferenziertem Gewebe:
- interkalare Meristeme (Spezialfall der Restmeristeme vor allem in Sprossachsen): zwischen bereits differenzierten Bereichen liegendes Meristem, das lokales Längenwachstum bewirkt
- Meristemoide: meristematische Zellgruppen oder Einzelzellen, die letztendlich in Dauergewebe umgewandelt werden
Sekundäre Meristeme
Der junge Keimling besitzt zunächst lediglich ein einziges primäres Sprossapikalmeristem, das bei zweikeimblättrigen Pflanzen zwischen den beiden Keimblättern zu finden ist und während der Embryogenese angelegt wird. Dieses Meristem ist durch die Produktion von Spross und Blättern für die Bildung der primären Sprossachse verantwortlich.
Die Etablierung weiterer Sprossachsen findet durch die Anlage von Meristemen in den Achseln von Laubblättern statt, die als Achselmeristeme oder laterale Meristeme bezeichnet werden. Die unterschiedlich ausgeprägte Aktivität der Achselmeristeme ist eine Grundlage für die mannigfaltigen Wuchsformen im Pflanzenreich. Die Aktivität der Achselmeristeme wird häufig durch das primäre Sprossapikalmeristem zunächst unterdrückt, so dass Seitentriebe verzögert auswachsen. Diese Kontrolle durch das Sprossapikalmeristem wird als Apikaldominanz bezeichnet. Die Bildung lateraler Wurzelmeristeme findet in keinem anatomisch definierten Bereich in einem gewissen Abstand vom primären Wurzelmeristem statt. Hier ist eine innere Gewebeschicht, das Perizykel, der Ursprung dieser lateralen Wurzelmeristeme.
Mit dem Begriff laterale Meristeme werden – neben den sekundären Meristemen in den Achseln der Laubblätter und der Wurzelspitzen – auch das Kambium und das Phellogen bezeichnet, beides teilungsaktive Zellschichten, die im äußeren Bereich des Sprosses für das sekundäre Dickenwachstum vieler Pflanzen verantwortlich sind.
Struktur
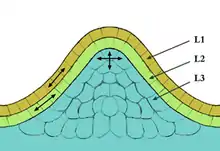
mit den Schichten L1, L2 und L3
Das primäre Sprossapikalmeristem der Bedecktsamer lässt sich in verschiedene Schichten und Zonen aufteilen:
- Die äußere Zellschicht, die Epidermis, wird als L1-Schicht bezeichnet.
- Die darunter liegende Zellschicht ist die L2-Schicht.
- Das weiter innen liegende Gewebe besteht aus mehr als einer Zellschicht und wird L3-Schicht oder auch Corpus genannt.
Die Schichten L1 und L2 bilden zusammen die sogenannte Tunica. Die Zellen teilen sich hier lediglich in antikliner Orientierung, was dazu führt, dass ihre Nachkommen im Allgemeinen in derselben Schicht bleiben. Zellen der L3-Schicht teilen sich in anti- und perikliner Orientierung, so dass in der L3-Schicht keine einzelnen Zellschichten voneinander abgegrenzt werden können.
Eine weitere Form der Einteilung bezieht sich eher auf die Funktion des Meristems. So sind in der zentralen Zone die eigentlichen Stammzellen lokalisiert, die durch Teilung Zellen zur Differenzierung an die periphere Zone des Meristems abgeben. In dieser peripheren Zone beginnen sich die Zellen zu den lateralen Organen, wie beispielsweise den Blättern, zu differenzieren. Unterhalb der zentralen Zone ist die Rippenzone lokalisiert, die hauptsächlich zum Wachstum des Sprosses beiträgt.
Entwicklung
Das Sprossapikalmeristem ist theoretisch in der Lage, seine Teilungsaktivität nahezu unbegrenzt fortzuführen, und dies geschieht auch häufig, wie beispielsweise bei Bäumen. Abhängig von zahlreichen Faktoren wie äußeren Einflüssen, Position an der Pflanze oder Alter der Pflanze terminiert das Sprossmeristem aber ebenso häufig, indem es verschiedene Entwicklungsstufen durchläuft und sich schließlich zu einem Blütenmeristem umwandelt, das mit der Produktion einer Blüte seine Aktivität einstellt. Die letzten lateralen Organe, die innerhalb dieses festgelegten Programms vom Meristem produziert werden, sind Kelchblätter, Kronblätter, Staubblätter und Fruchtblätter (in dieser Reihenfolge). Blüten sind demnach terminierte Sprossapikalmeristeme.
Siehe auch
Literatur
- Strasburger – Lehrbuch der Botanik. Andreas Bresinsky, Christian Körner, Joachim W. Kadereit, Gunther Neuhaus, Uwe Sonnewald, Spektrum Akademischer Verlag, 36. Auflage 2008 ISBN 978-3-8274-1455-7