Wasseraffen-Theorie
Als Wasseraffen-Theorie (auch: Wassertheorie, Wasseraffen-Hypothese) wird eine Reihe von teils spekulativen Hypothesen bezeichnet, nach der die Vorfahren des anatomisch modernen Menschen (Homo sapiens) im Verlauf der Menschwerdung eine teilweise wasserlebende bzw. amphibische Phase durchgemacht haben sollen. In Fachkreisen konnte sich diese Hypothese nicht durchsetzen.[1][2][3]
Einführung
Die Vertreter der Wasseraffen-Theorie sind der Ansicht, dass frühe Vormenschen sich über lange Zeitspannen und ausdauernd an und in Gewässern und an Uferregionen aufhielten. Dies habe zur Folge gehabt, dass sich einige spezielle evolutionäre Anpassungen an diesen Lebensraum entwickelten, die charakteristisch für den anatomisch modernen Menschen sind. Nach dieser „aquatischen Phase“ seien die Hominini wieder zum Leben ausschließlich an Land übergegangen.
Anatomische und verhaltensbiologische Merkmale, die zur Stützung dieser Theorie herangezogen werden, sind insbesondere:
- der aufrechte Gang
- die verlängerten hinteren Gliedmaßen
- die besondere Struktur der Haut und die Einlagerung des Fettgewebes in der Unterhaut im Vergleich zu Menschenaffen
- der im Vergleich zu anderen Landsäugern deutlich höhere Körperfettanteil des Menschen
- die für die Strömungsverhältnisse bei der Atmung ungünstige nach unten offene Nase
- der Tauchreflex
- der Stimmritzenkrampf als Schließreflex des Kehlkopfes
- die Kontrolle der Atmung beim Tauchen als Voraussetzung zur Entwicklung einer Lautsprache
- die für ein ausschließlich an Land lebendes Säugetier regelabweichend großen Füße
Geschichtlicher Überblick
Die erste Wasseraffen-Theorie: Westenhöfers Aquatile Hypothese
Einige der Hauptargumente der Wasseraffen-Theorie wurden bereits ab 1923 durch den Berliner Pathologen Max Westenhöfer (1871–1957) aufgestellt; eine Zusammenfassung seiner Ideen ist in seinem 1942 publizierten Buch Der Eigenweg des Menschen zu lesen. Er glaubte, dass im Verlauf der Stammesgeschichte des Menschen eine aquatische Phase stattgefunden habe, und belegte dies anhand anatomischer Vergleiche. Er schloss eine Abstammung oder nahe Verwandtschaft von Menschen zu Menschenaffen aus und sah stattdessen den Menschen als direkten (weil ebenso wenig spezialisierten und im Feuchten lebenden) Nachkommen von Lurchen und Salamandern.[4]
Hardys „Aquatic Ape Theory“ und die Weiterentwicklung dieses theoretischen Modells
Vermutlich unabhängig von Westenhöfer kamen dem Meeresbiologen Alister Hardy um 1930 die ersten Gedanken zu diesem Konzept. Er las eine Veröffentlichung, in der Besonderheiten des menschlichen Unterhaut-Fettgewebes beschrieben wurden. Dieses ist – anders als bei den meisten Säugetieren – so fest mit der Haut verbunden, dass es mit angehoben wird, wenn man an der Haut zieht. Hardy, ein Meeresbiologe, assoziierte damit die Verhältnisse bei Walen und überlegte, dass der Mensch in seiner Vorgeschichte eine aquatische Phase gehabt haben könne. Im Laufe der Jahre sammelte er weitere Indizien, zögerte aber mit der Veröffentlichung. Erst im März 1960 publizierte Hardy seine Gedanken in dem Artikel Was man more aquatic in the past in der populärwissenschaftlichen Zeitschrift New Scientist.[5] Hardy bezeichnete seine Hypothesen in diesem Artikel ausdrücklich als „Spekulation“; er hat kein Buch über das Thema veröffentlicht.
Seine Hypothesen wurden zehn Jahre später durch die britische Journalistin Elaine Morgan aufgegriffen[6] und populärwissenschaftlich und in Verbindung mit feministischen Ansichten weiterentwickelt. Morgan veröffentlichte mehrere Werke zu dem Thema. Weitere Anhänger der Aquatic Ape Theory (Wasseraffen-Theorie) sind unter anderen der belgische Allgemeinmediziner Marc Verhaegen,[7] die schwedische Biologin und Sporttaucherin Erika Schagatay,[8] der in Australien lebende, aus Großbritannien stammende IT-Experte Algis Kuliukas[9] sowie der Schweizer Sportwissenschaftler Renato Bender und die Schweizer Ärztin Nicole Bender-Oser, die eine gemeinsame Website zur Wasseraffen-Theorie unterhalten.[10]
1987 wurde in Valkenburg (Niederlande) ein Symposium zu dieser Hypothese durchgeführt, bei dem Verfechter und Gegner ihre Ansichten austauschten. Das zweite Symposium fand 1999 unter dem Titel „Water and Human Evolution“ in Gent (Belgien) statt.
Argumente der Befürworter der Wasseraffen-Theorie
Generell unterliegen alle Hypothesen zur Erklärung der spezifischen Merkmale des Menschen im Verlauf der Evolution einem Problem: Die Argumente beziehen sich in erster Linie auf Vergleiche mit rezenten anatomischen Strukturen und mit Knochenfossilien. Für die meisten evolutionsbiologisch relevanten Merkmale wäre jedoch eine Weichteilerhaltung der Fossilien notwendig, um den Beweis zu führen. Zudem ist durch Fossilien belegt, dass es in den Gewässern in der Nähe von Hominini-Fundstellen „gewimmelt hat von hungrigen Krokodilen und aggressiven Flusspferden. Unsere kleinen, wehrlosen Vorfahren hätten bei einer Konfrontation mit solchem Getier keine Chance gehabt.“[11] Auch war es bisher nicht möglich, aus der Wasseraffen-Theorie überprüfbare Vorhersagen abzuleiten, die zu ihrer weiteren Untermauerung hätten beitragen können.
Unterhautfettgewebe
Der Mensch besitzt als einziger Primat und neben den wasserlebenden Walen, Robben und Seekühen als einziges landlebendes Säugetier ein ausgeprägtes, in Ansätzen auch wärmeisolierend wirkendes Unterhautfettgewebe (Speckschicht). Dies gab Hardy den ersten Anstoß zu seiner Hypothese. Unterhautfettgewebe isoliert im Wasser den Körper besser als nasse Haare. Andere Wissenschaftler sehen jedoch diese Fettschicht – insbesondere im Zusammenhang mit der Haarlosigkeit und den Schweißdrüsen – als Teil eines komplizierten thermoregulatorischen Systems an. Das Unterhautfettgewebe wäre demnach als Kompensation der mangelhaften Isolierung durch die fehlende Behaarung einzuordnen. Eine weitere Erklärung für dieses Phänomen besagt, dass die Speckschicht der Nahrungsreserve diene und insbesondere den erhöhten Energiebedarf des Gehirns bei Nahrungsmangel decke.
Besonders ausgeprägt ist der Unterschied im Anteil des Fettgewebes am Körpergewebe von menschlichen Säuglingen (vergleichsweise hoher Fettanteil) im Vergleich zu Menschenaffen-Säuglingen, die eher dünn sind. Nach der Wasseraffen-Hypothese schützte dies Säuglinge vor dem Auskühlen im Wasser, wenn sie mit ihren Müttern in das Wasser gingen. Das im Vergleich der Geschlechter eher dürftige Unterhautfettgewebe der Männer wird bei dieser Hypothese allerdings nicht erklärt. Zudem ist die durchschnittliche Dichte des Körpers von menschlichen Säuglingen deutlich geringer als bei anderen Landsäugern, so dass sie mehr Auftrieb haben. Ein anderer Erklärungsansatz sieht in diesem Merkmal ausschließlich eine Energiereserve des Säuglings.
Auch die beim Menschen im Vergleich zu anderen Primaten ungewöhnliche Fettverteilung wird als Argument für einen regelmäßigen Aufenthalt im Wasser angeführt. Während bei anderen Primaten das Fettgewebe so am Körper verteilt ist, dass Kopf, Hals und Abdomen gut isoliert sind und Wärme hauptsächlich über die (Hinter-)Beine abgegeben wird, wird beim Menschen der Wärmeaustausch über Gesicht, Hals, Schultern und den oberen Thorax durchgeführt. Die untere Körperhälfte dagegen ist durch das Fettgewebe an unterem Abdomen, Gesäß und Beinen gut isoliert. Von Vertretern von Wasser- bzw. Uferhypothesen der menschlichen Evolution wird dieser Umstand als Anpassung an regelmäßiges Waten im Wasser gedeutet.[12]
Unterschiede in der Ausprägung des Gewebes gibt es auch von Mensch zu Mensch. So überlebte der isländische Fischer Guðlaugur Friðþórsson dank eines selten dicken Unterhautfettgewebes etwa sechs Stunden im 5 Grad kalten Atlantik.[13]
Tauchreflex
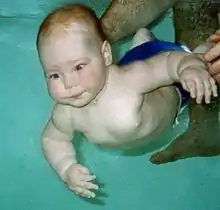
Ein wesentliches Argument für die Hypothese ist das Vorhandensein eines Tauchreflexes beim Menschen: Taucht man einen Säugling unter Wasser, wird er kein Wasser verschlucken. Säuglinge sind etwa bis zum zehnten Lebensmonat fähig, den Atem reflexhaft anzuhalten, und sie erlernen das Schwimmen schnell; dies wird u. a. in Baby-Schwimmkursen genutzt. Der Tauchreflex verlangsamt den Herzschlag beim Tauchen und sorgt dafür, dass das Gehirn vermehrt mit Blut versorgt wird.
Gegen die Deutung des Tauchreflexes im Sinne der Wasseraffen-Theorie wird eingewandt, dass ein solcher Reflex bei vielen landlebenden Säugetieren zu beobachten und er daher wahrscheinlich ein ursprüngliches Merkmal der Säugetiere sei, auch jener, denen keine „aquatische Phase“ zugeschrieben wird.
Schwimmhäute
Bei den meisten Menschen ist vor allem zwischen den Fingern, teilweise auch zwischen den Zehen eine flache Hautpartie ausgebildet, die als Rudiment von Schwimmhäuten interpretiert werden kann. Schimpansen und andere Menschenaffen sollen dieses Merkmal nicht zeigen. Bei einigen Menschen kommt es zu einer fast vollständigen Ausbildung solcher „Schwimmhäute“ (Atavismus). Diese Hautbespannungen deutet Morgan auch als „Schwimmhäute“, die während der aquatischen Phase erworben wurden und möglicherweise wieder teilweise reduziert wurden. Langdon setzt dem entgegen, dass diese „Schwimmhäute“ bei Feten aller Primaten ganz ausgebildet sind und vor der Geburt reduziert werden.[1] Beim Menschen sei die Reduzierung der „Schwimmhäute“ deswegen unvollständig, weil der Mensch an sich neoten sei, das heißt, dass typische Merkmale jugendlicher Stadien bis ins Erwachsenenalter beibehalten werden. Möglicherweise ist dies allerdings lediglich eine Folge der verkürzten Schwangerschaft und der damit einhergehenden Vorverlegung der Geburt, die trotz der Verdreifachung des späteren Gehirnvolumens und der damit einhergehenden Zunahme des Kopfumfanges eine Passage des Geburtskanals ermöglicht.
Der aufrechte Gang
Der aufrechte Gang und die Umstrukturierung des Beckens werden als weitere Belege für die Wasseraffen-Hypothese dargebracht. Wenn sich Menschenaffen gezwungen sehen, in das Wasser zu gehen, dann richten sie sich in der Regel auf und gehen auf den Hinterbeinen. Wenn der Mensch sich über lange Zeit im und am Wasser aufgehalten habe und sich schwimmend fortbewegte, dann habe er sich dem Wasser stromlinienförmig anpassen müssen. Das erkläre das Kippen des Beckens und die Verlagerung der Beine unter den Körper.
Von Paläoanthropologen werden mehrere Hypothesen zum Entstehen des aufrechten Ganges bei den Vorfahren des anatomisch modernen Menschen erörtert, die ohne Verweis auf eine „aquatische Phase“ auskommen (siehe Hauptartikel).
Der Haarstrich
Die Körperbehaarung des Menschen weist ein stromlinienförmiges Muster auf, welches der Richtung des umgebenden Wassers beim Vorwärtsschwimmen entspricht. Dabei könnte es sich um eine Zwischenphase der Anpassung vor dem Verlust des Felles gehandelt haben.[14] Dieses Phänomen lässt sich aber auch durch schnelleres Abperlen von Regenwasser bei aufrechtem Gang erklären. Möglicherweise aus dem gleichen Grund besitzen auch andere landlebende Säugetiere (wie z. B. Hunde) ein ganz ähnliches Muster in ihrem Haarstrich.
Nacktheit
Die spärliche Behaarung des Menschen wird ebenfalls als Argument für die Wasseraffen-Hypothese angeführt. Verwiesen wird darauf, dass Haarlosigkeit im Wasser einen geringeren Strömungswiderstand als Fell biete. Zudem sei die Funktion des Felles für den Schutz gegen UV-Strahlen bei amphibischer Lebensweise weniger wichtig, außer am Kopf. Eine weitere These verweist auf eine Auslese von „stark behaarten“ Wasseraffen durch die Erkältungsproblematik bei nassem Fell. Es sei also nicht so sehr die effiziente Schwimmanpassung erfolgt, sondern die allgemeine Adaption an Flachwassergebiete.[15]
Von Paläoanthropologen werden mehrere Hypothesen zum Verlust der Behaarung bei den Vorfahren des anatomisch modernen Menschen erörtert, die ohne Verweis auf eine „aquatische Phase“ auskommen (siehe Hauptartikel).
Geruchssinn
Ein weiteres Argument ist der reduzierte Geruchssinn des Menschen. Morgan zufolge sei der Geruchssinn verkümmert, da er im Wasser kaum eine Bedeutung habe. Zwar ist die Wahrnehmung von Gerüchen im Wasser sehr gut möglich, jedoch nur mit entsprechend angepassten Riechorganen, wie bei Haien und anderen Fischen. Der Geruchssinn der Primaten ist primär jedoch an der Übertragung von Duftstoffen über die Luft angepasst. Morgan schließt daraus, dass der Geruchssinn sich zurückgebildet habe, weil er im Wasser nicht mehr nützlich war. Als Gegenargument wird genannt, dass der hypothetisch aquatische Affe nahezu die ganze Zeit im Wasser gelebt haben müsse, damit eine Reduzierung des Geruchssinnes keinen Selektionsnachteil darstelle. Aufgrund der sehr unvollständigen Anpassung des aquatischen Affen an das Wasser ist dies jedoch unwahrscheinlich. Morgan berücksichtige zudem nicht, dass sämtliche Trockennasenaffen, eine Unterordnung der Primaten, zu der auch der Mensch zählt, einen schlecht entwickelten Geruchssinn besitzen, während stark ans Wasser gebundene Säuger wie Otter, Biber, Wasserspitzmäuse oder selbst Robben einen recht guten Geruchssinn besitzen.
Nährstoffbedürfnisse
Ebenfalls als Indiz für eine wassergebundene Lebensweise deuten die Vertreter der Wasseraffentheorie die Nährstoffbedürfnisse des Menschen. Einige für die Entwicklung des Gehirns wichtige Nährstoffe wie DHA, EPA, Jod, und Taurin seien nur im Wasser in ausreichender Menge verfügbar.[16]
In einer 2019 veröffentlichten Studie wurde festgestellt, dass Bonobos im kongolesischen Salonga-Nationalpark ihren Jodbedarf durch den Verzehr von Wasserpflanzen decken. Einige der untersuchten Wasserpflanzen haben Jodgehalte, die beinahe so hoch wie in Meeresalgen seien. Terrestrische Pflanzen im selben Lebensraum wiesen dagegen keinen hohen Jodgehalt auf. Die Autoren der Studie sehen darin eine mögliche Erklärung, wie Menschen abseits von Küsten ihren Jodbedarf gedeckt haben können.[17][18]
Dagegen wird eingewendet, dass auch der Verzehr von Fleisch die genannten Nährstoffe in ausreichender Menge liefern könne.
Tränenflüssigkeit und Schweiß
Als Beleg für eine marine Episode unserer Vorfahren wird jedoch die Salinität der menschlichen Tränenflüssigkeit gewertet. Mit einem durchschnittlichen Salzgehalt von 3,5 % liegt diese im Bereich des Meerwassers. Diese Konzentration ist typisch für Meeres-Säuger. Terrestrische Säugetiere haben dagegen i. d. R. deutlich weniger Salze in ihrer Tränenflüssigkeit (Ausnahme: der Elefant, für den allerdings eine Abstammung von marinen Vorfahren vorgeschlagen wird).
Als Hinweis auf marine Vorfahren wird auch der – im Vergleich zu anderen Säugern – „verschwenderisch“ hohe Salzgehalt von menschlichem Schweiß interpretiert. Er könnte zum Beispiel zur Ausscheidung überflüssiger, durch die Nahrung aufgenommener Salze gedient haben.
Fossilfunde
Als weiteres Indiz wird auf Befunde aus vielen frühen Hominiden-Fundstätten verwiesen, durch die belegt ist, dass die erhalten gebliebenen Knochen in aquatische Sedimente eingebettet wurden (Flüsse, Seen). Dies gilt für
- Orrorin tugenensis (wahrscheinlich kein direkter Vorfahr des Menschen, vor 5,6–6,3 Millionen Jahren)
- Ardipithecus ramidus, Äthiopien, vor 4,4 Millionen Jahren
- Australopithecus anamensis, Turkanasee, vor 3,6 Millionen Jahren
- Australopithecus afarensis, vor 3,2–4,1 Millionen Jahren
- Kenyanthropus platyops, vor 3,3–3,5 Millionen Jahren
Lediglich ein Fundort (Laetoli) weist keine aquatisch entstandenen Sedimente, sondern vulkanische Tuffe auf.
Spätere Funde von Australopithecus africanus (vor 2 Millionen Jahren) liegen dagegen häufig nicht in aquatischem Kontext.
Bei der Interpretation dieser Befunde muss allerdings berücksichtigt werden, dass in Sedimenten von Flüssen und Seen eine deutlich höhere Wahrscheinlichkeit der Fossilisation von organischer Substanz (wie z. B. Knochen) besteht als wenn die Knochen an der Erdoberfläche – zumal in der Nähe von Wasserstellen – liegen bleiben und dort von Herdentieren zertreten werden. Die Entdeckung von Hominiden-Fossilien in aquatischen Ablagerungen allein ist daher keine Stütze für die Wasseraffen-Theorie.
Bei fossilen Neandertalern hat man auffällig häufig Gehörgangsexostosen gefunden. Bei etwa der Hälfte der Funde fossiler Neandertaler ließen sich entsprechende Veränderungen des Gehörgangs nachweisen. Die Häufigkeit dieses, im Englischen auch als Surfer’s ear bezeichneten Krankheitsbildes, deutet auf einen regelmäßigen Aufenthalt in (kühlem) Wasser hin.[19] Ein weiteres Indiz für diese Theorie sind Funde von Werkzeugen aus Muscheln, die möglicherweise nur durch Tauchen gesammelt werden konnten.[20]
Kuliukas Wat-Affen-Modell

Der Londoner Anthropologe Algis Kuliukas unterzog in seiner Master-Arbeit 2001 die Wasseraffen-Hypothese einer Untersuchung vor dem Hintergrund aktueller Forschungsergebnisse zu Hominiden-Fossilien sowie der Verhaltensforschung. Dabei modifizierte und präzisierte er das Modell zu einem „Wat-Affen-Modell“ (wading ape) zur Erklärung der zweibeinigen Fortbewegung des Menschen. Zugleich versuchte er Kriterien der Überprüfbarkeit anzuführen: Falls das Wat-Affen-Modell zutrifft, so müssten rezente Menschenaffen ebenfalls waten, wenn sie angeregt werden, sich in das Wasser zu begeben. Falls sie sich dagegen vierfüßig in das Wasser begeben, so wäre das Modell widerlegt.
Tatsächlich ist bekannt, dass Tiefland-Gorillas, die in sumpfigem Gelände leben, ein zweibeiniges Waten beim Eintritt in das Wasser zeigen. Aus der Literatur ist außerdem bekannt, dass dieses Verhalten auch bei Orang-Utans, Schimpansen und Bonobos auftritt.
Darauf folgende Untersuchungen an Bonobos in einem Wildpark bei Brüssel zeigten, dass diese (dem Menschen genetisch am nächsten verwandten) Menschenaffen an Land lediglich zu 2 Prozent den aufrechten Gang benutzten. Sobald sie teilweise im Wasser waren, stieg die Rate auf 50 Prozent. Dabei benutzten sie ihre Arme fast immer zum Halten der Balance. Bei Bonobos, die vollständig im Wasser waren, wurde zu 92 Prozent aufrechter Gang beobachtet, und zwar fast immer ohne Unterstützung durch die Arme.
Um die richtige historische Perspektive zu bewahren, ist es wichtig, zu erkennen, dass Kuliukas’ Thesen keine Neuerung in der Wasseraffendiskussion darstellen. Als Beispiel sei folgende Aussage aus einem Artikel über die Entwicklung der Bipedie des Menschen zu erwähnen, die im Jahr 1997 im Anthropologischen Anzeiger veröffentlicht wurde:
„Die Vorteile des aufrechten Gangs lassen sich durch die Vielseitigkeit dieser Fortbewegungsart in einer semi-aquatischen Lebensweise verstehen. Die Bipedie der frühen Hominiden kann nicht als eine ideale Anpassung an eine isolierte Fortbewegungsart bezeichnet werden, denn die menschliche Bipedie deckt ein großes Spektrum von lokomotorischen Beanspruchungen ab wie das Tauchen, Schwimmen, Waten, Klettern auf Bäumen und Felsen und terrestrische Fortbewegung.“[21]
Ferner erscheint es fragwürdig, die Bestätigung oder Widerlegung der Wassertheorie von einem einzigen Verhaltensmerkmal (Waten bei Menschenaffen) abhängig zu machen. Der wichtigste Aspekt dieser Theorie basiert auf der Berücksichtigung verschiedener Merkmale des Menschen, die als Konvergenzen zu dem Merkmalsmosaik anderer Lebewesen gedeutet werden. Und schließlich war es schon vor der Aufstellung Kuliukas’ Thesen bekannt, dass Primaten (darunter auch Menschenaffen) im seichten Wasser häufig waten,[22] so dass die in der Wissenschaft übliche Reihenfolge „Hypothese aufstellen – Hypothese überprüfen“ hier nicht eingehalten wurde. Kuliukas verdient trotzdem Anerkennung in der Wassertheorie-Debatte, unter anderem für seinen Versuch, eine einheitliche Definition der Wassertheorie zu liefern.
Andere „aquatische“ Primaten
Im Zusammenhang mit dem teilweise vehement ausgetragenen Streit um die Wasseraffen-Hypothese stellt sich die Frage, ob die postulierte Anpassung der menschlichen Vorfahren an das Wasser ein einmaliger Vorgang in der Gruppe der Primaten sei.
Tatsächlich gibt es mindestens drei Beispiele für einen unabhängig erfolgten sekundären Übergang von Affen an eine semi-amphibische Lebensweise:
- der Nasenaffe (Nasalis larvatus), der in den Regenwäldern des Tieflands und den Mangrovensümpfen von Borneo lebt. Diese Tiere gehören zu den sogenannten Schlankaffen und zeichnen sich durch gutes Schwimm- und Tauchvermögen[23] und eine große Nase aus. Bei Weibchen ähnelt diese der menschlichen Nase, bei Männchen ist sie extrem groß (Sexualdimorphismus).
- der Javaneraffe (Macaca fascicularis), der in Südostasien von Myanmar über Thailand, Indonesien bis zu den Philippinen vorkommt. Javaneraffen gehören zu den Pavianartigen. Sie leben in Wäldern, bevorzugt in der Nähe von Gewässern, auch an der Meeresküste. Diese Tiere schwimmen und tauchen häufig. Zu ihrer bevorzugten Nahrung gehören Krabben, die sie am Strand ausgraben und waschen. Sie tauchen mit geöffneten Augen. Das Waschen der Nahrung (auch Früchte werden gewaschen) gilt als nicht ererbte, sondern kulturell weitergegebene Verhaltensweise.[24]

- der Japanmakak, (Macaca fuscata), der ebenfalls zu den Pavianartigen gehört. Er lebt auf den japanischen Inseln und badet – insbesondere im Winter – gern in heißen Quellen. Eine Makakenpopulation auf der Insel Kōjima ist seit 1948 unter wissenschaftlicher Beobachtung. Besonders bekannt wurde das Weibchen Imo aus dieser Population, das 1953 das Waschen von Nahrung erfand. Diese Verhaltensweise hat sich seitdem durch Tradierung durchgesetzt. Später setzte sich auch das Verhalten durch, Nahrung im Meer zu waschen und damit zu salzen. Seitdem setzte sich bei dieser Makakengruppe das Leben im Meer erstaunlich schnell durch, obwohl Japanmakaken generell das Meer meiden. Mitglieder dieser Population entwickelten seitdem das Schwimmen, Tauchen und die Nutzung von Seetang als Nahrung. Zudem begannen diese Affen, die Hände zum Tragen des Futters zu benutzen und größere Strecken (bis 30 m) zweibeinig zu laufen. Anfang der 1980er Jahre begannen die Makaken von Kōjima, rohen Fisch zu essen.
Allen drei Beispielen gemeinsam ist, dass diese Arten zwar weiterhin primär Landbewohner bzw. Baumbewohner sind, aber sekundär zusätzlich den Lebensraum Wasser zumindest teilweise erschlossen haben. Grundsätzlich ist also der Bauplan von Primaten auch geeignet, um eine sekundär aquatische Lebensweise zu entwickeln.
Das Beispiel der Japanmakaken von Kōjima zeigt, dass bei der Diskussion von Hominisationsmodellen nicht nur die klassischen Evolutionsmechanismen (Mutation / Selektion) zu berücksichtigen sind. Vielmehr können auch bei Affen durch Entdecken, Lernen und Tradieren neue Verhaltensweisen entstehen und – unabhängig von der Vererbung – über Generationen weitergegeben werden (Meme). Solche neuen Verhaltensweisen können – wie die Japanmakaken zeigen – zu veränderten Lebensbedingungen einschließlich einer Habitatveränderung und einer Änderung der ökologischen Nische führen und damit auf die Evolutionsvorgänge zurückwirken.
Dies kann erstaunlich rasch geschehen, im Fall der Japanmakaken innerhalb von kaum 50 Jahren. Es müssen also nicht zwingend externe ökologische Faktoren (Klimaänderungen, geologische Änderungen etc.) eingewirkt haben, vielmehr könnten neue Nahrungsvorlieben und Verhaltensweisen entstanden sein, die dann die Lebensweise und den Lebensraum der Vorfahren des Menschen änderten. Damit wären sekundär andere Umwelt- und damit Selektionsbedingungen geschaffen, die nun die Durchsetzung genetisch bedingter Anpassungen begünstigen würden.
Literatur
Artikel
- Renato Bender, Marc Verhaegen, Nicole Oser: Der Erwerb menschlicher Bipedie aus der Sicht der Aquatic Ape Theory. In: Anthropologischer Anzeiger. 55 (1), 1997, 1–14, Zusammenfassung
- Renato Bender, Nicole Oser: Gottesanbeterinnen, Maulwürfe und Menschen. Unipress (Berichte über Forschung und Wissenschaft an der Universität Bern, hrsg. v. der Pressstelle) 95, 1997, S. 20–26, Volltext (Memento vom 28. Januar 2006 im Internet Archive)
- Alister Hardy: Was man more aquatic in the past. In: New Scientist. 17. März 1960, S. 642–645, Volltext (PDF)
- John H. Langdon: Umbrella hypotheses and parsimony in human evolution: a critique of the Aquatic Ape Hypothesis. In: Journal of Human Evolution. Band 33, Nr. 4, 1997, S. 479–494, doi:10.1006/jhev.1997.0146
Monographien
- Renato Bender: Die evolutionsbiologische Grundlage des menschlichen Schwimmens, Tauchens und Watens: Konvergenzforschung in den Terrestrisierungshypothesen und in der Aquatic Ape Theory. Diplomarbeit am Institut für Sport und Sportwissenschaft, Universität Bern, 1999.
- Nicole Bender-Oser: Die Aquatile Hypothese zum Ursprung des Menschen: Max Westenhöfers Theorie und ihre Bedeutung für die Anthropologie. Inaugural-Dissertation zur Erlangung der Doktorwürde der Humanmedizin der Medizinischen Fakultät der Universität Bern, 2004. (eine ausführliche Arbeit zur Geschichte der Aquatilen Hypothese)
- Carsten Niemitz: Das Geheimnis des aufrechten Gangs: unsere Evolution verlief anders. Beck, München 2004, ISBN 978-3-406-51606-1.
- Elaine Morgan: Kinder des Ozeans. Der Mensch kam aus dem Meer (Deutsche Ausgabe von The Aquatic Ape: A Theory of Human Evolution. Goldmann Verlag, 1989, ISBN 978-3-442-11435-1)
- Elaine Morgan: Der Mythos vom schwachen Geschlecht: wie die Frauen wurden, wie sie sind. Goldmann, München 1990, ISBN 3-442-11457-8.
- Elaine Morgan: The Aquatic Ape: A Theory of Human Evolution.
- Elaine Morgan: The Scars of Evolution: What Our Bodies Tell us about Human Evolution.
- Elaine Morgan: The Aquatic Ape Hypothesis: The Most Credible Theory of Human Evolution.
- Max Westenhöfer: Der Eigenweg des Menschen: dargestellt auf Grund von vergleichend morphologischen Untersuchungen über die Artenbildung und Menschwerdung. Mannststaedt, Berlin 1942
Weblinks
- Elaine Morgan fasst ihre Sicht der Theorie und die Situation des wissenschaftlichen Diskurses in einem TED-Talk zusammen, Video: Juli 2009, 17:17 min
- Michael Bujatti-Narbeshuber: Vortrag: Litoraltheorie der Hominisation. (PDF; 547 kB) 2. Symposium für Neurobiologie in Salzburg 1991. In: Agemus. Band 26, 1991.
- aquaticape.org Umfangreiche Darstellung der Theorie (englisch)
- Corina Gericke: Affenkultur (Beobachtungen an Japanmakaken)
Einzelnachweise
- John H. Langdon: Umbrella hypotheses and parsimony in human evolution: a critique of the Aquatic Ape Hypothesis. In: Journal of Human Evolution. Band 33, Nr. 4, 1997, S. 479–494, doi:10.1006/jhev.1997.0146
- Stephen Ornes: Whatever Happened To… the Aquatic Ape Hypothesis? discovermagazine.com vom 5. September 2007. Auch hier heißt es: „the aquatic ape hypothesis never got much support from the scientific community.“
- HM Dunsworth: Human Origins 101. Greenwood Press, Westport 2007, Science 101 Series, ISBN 978-0-313-33673-7, S. 121.
- Bender-Oser, 2004, S. 129
- Alister Hardy: Was man more aquatic in the past. In: New Scientist. 17. März 1960, S. 642–645, Volltext (PDF) (Memento vom 19. Oktober 2016 im Internet Archive)
- William F. Williams (Hrsg.): Encyclopedia of Pseudoscience. Taylor & Francis, Hoboken 2013, ISBN 978-1-135-95522-9, S. 169 f.
- Marc Verhaegen und die Wasseraffen-Theorie
- Erika Schagatay, Publikationen
- Algis Kuliukas: Curriculum Vitae 2014.
- aquatic-hypothesis.com
- Nina Jablonski: Why the Aquatic Ape Theory Doesn' Hold Water. In: Scientific American. Sonderheft: The Story of Us. Herbst 2016, S. 55
- Carsten Niemitz: The evolution of the upright posture and gait—a review and a new synthesis. In: Naturwissenschaften. Band 97, Nr. 3, 1. März 2010, ISSN 1432-1904, S. 241–263, doi:10.1007/s00114-009-0637-3, PMID 20127307, PMC 2819487 (freier Volltext).
- Alva Gehrmann: Naturphänomen: Das Geheimnis des Seehund-Mannes. In: FAZ.NET. ISSN 0174-4909 (Online [abgerufen am 28. April 2020]).
- Desmond Morris: Der Mensch, mit dem wir leben. Kapitel: „Verhalten im Wasser“
- P. Waugh: WAT. University of Oxford
- Diet of Homo ancestors. Abgerufen am 28. April 2020.
- Bonobos suchen gezielt nach Pflanzen mit hohem Jodgehalt - Wissenschaft aktuell. Abgerufen am 28. April 2020.
- Gottfried Hohmann, Sylvia Ortmann, Thomas Remer, Barbara Fruth: Fishing for iodine: what aquatic foraging by bonobos tells us about human evolution. In: BMC Zoology. Band 4, Nr. 1, 2. Juli 2019, ISSN 2056-3132, S. 5, doi:10.1186/s40850-019-0043-z.
- Erik Trinkaus, Mathilde Samsel, Sébastien Villotte: External auditory exostoses among western Eurasian late Middle and Late Pleistocene humans. In: PLOS ONE. Band 14, Nr. 8, 14. August 2019, ISSN 1932-6203, S. e0220464, doi:10.1371/journal.pone.0220464, PMID 31412053, PMC 6693685 (freier Volltext).
- Paola Villa, Sylvain Soriano, Luca Pollarolo, Carlo Smriglio, Mario Gaeta: Neandertals on the beach: Use of marine resources at Grotta dei Moscerini (Latium, Italy). In: PLOS ONE. Band 15, Nr. 1, 15. Januar 2020, ISSN 1932-6203, S. e0226690, doi:10.1371/journal.pone.0226690, PMID 31940356, PMC 6961883 (freier Volltext).
- Bender, Verhaegen, Oser, 1997, S. 1. Auch: Bender, 1999, S. 140ff.
- Bibliographie dazu in Bender, 1999, S. 109ff.
- Proboscis Monkey (Nasalis larvatus).
- Macaca fascicularis – long-tailed macaque.