Stammesgeschichte der Krokodile
Die heutigen Krokodile (Crocodylia) sind die modernsten Vertreter einer Entwicklungslinie der Archosaurier, die sich bereits vor etwa 250 Millionen Jahren, in der frühen Trias, von der Linie der Flugsaurier und Dinosaurier getrennt hat.
Begrifflichkeit „Krokodil“ im paläontologischen Kontext
Bereits vor mehr als 200 Millionen Jahren, in der späten Trias, hatte jene Archosaurier-Linie, die zu den heutigen Krokodilen führt, mit den Phytosauriern Vertreter hervorgebracht, die in Aussehen und Lebensweise heutigen Krokodilen relativ ähnlich waren. Daher heißt es in populärwissenschaftlichem Zusammenhang oft, dass Krokodile eine sehr alte Gruppe sind, und bisweilen werden sie als „lebende Fossilien“ bezeichnet.[1] Tatsächlich ist der Begriff „Krokodil“ nicht genau definiert und umfasst urtümlich wirkende, echsenartige, meist eher großwüchsige, semi-aquatische bzw. amphibische Reptilien, die nicht näher mit Brückenechsen, Eidechsen und Schlangen verwandt sind. Daher ist er im paläontologischen Zusammenhang eher weiter gefasst und bezieht auch zahlreiche ausgestorbene Formen ein, wohingegen „Krokodil“ im zoologischen Sinn eher enger gefasst und vorwiegend auf die rezenten Vertreter und deren unmittelbare fossile Verwandtschaft bezogen ist, deren Wurzeln deutlich weniger weit als 200 Millionen Jahre in die Vergangenheit zurückreichen (siehe → Die Crocodylia – die „eigentlichen“ Krokodile).
Einführung
Die Archosaurier-Linie, die zu den heutigen Krokodilen führt, wird Crurotarsi (oder, nach Neudefinition dieses Taxons auch Pseudosuchia)[2][3] genannt und die Linie, die zu den Flugsauriern und Dinosauriern und damit auch zu den heutigen Vögeln führt, heißt Ornithodira (oder Avemetatarsalia). Beide Linien gingen aus einem basalen Vertreter der Archosaurier-Linie (basale Archosauriformes, siehe auch → „Thecodontia“) hervor. Der wichtigste anatomische Unterschied zwischen Crurotarsiern und Ornithodiren besteht im Bau des Fußgelenks. Während es bei den Crurotarsiern zwischen den beiden oberen (proximalen) Fußwurzelknochen – Fersenbein (Calcaneus) und Sprungbein (Astragalus) – ausgebildet ist (metatarsales Gelenk), verläuft die Gelenklinie bei den Ornithodiren entlang des gemeinsamen Kontaktes von Astragalus und Calcaneus mit den unteren (distalen) Fußwurzelknochen (mesotarsales Gelenk). Im Gegensatz zu den basaleren Archosauriformes, die ebenfalls ein mesotarsales Gelenk besitzen, ist der Calcaneus bei den Ornithodiren stark reduziert. Unter anderem deshalb wird es auch als abgeleitetes Mesotarsalgelenk bezeichnet, um es vom „primitiven“ Mesotarsalgelenk der frühen Archosauriformes zu unterscheiden.
| Kronen-Diapsida (Sauria) |
| ||||||||||||||||||||||||||||||
| ••• |
| ||||||||||||||||||
Verwandtschaftsverhältnisse der basalen Archosauromorpha (nach Benton, 2007).[4] Fußnoten: 1) alternativ auch als Archosauria bezeichnet; 2) Vertreter, die früher unter dem Begriff Thecodontier zusammengefasst wurden; 3) Archosaurier-Kronengruppe, alternativ auch als Avesuchia bezeichnet
Die Crurotarsi – frühe Vorfahren


Von der Entwicklungslinie zu den Krokodilen zweigte eine Reihe von Archosauriergruppen ab, die heute ausgestorben sind. Zu diesen „primitiven“ Archosauriern aus der späten Trias, die mit den Crocodylomorpha als Crurotarsi (Pseudosuchia) zusammengefasst werden, zählen unter anderem die Phytosauria, die Aetosauria und die Rauisuchia. Während die Phytosauria im Habitus relativ krokodilähnlich waren, wiesen die Rauisuchier eher Parallelen zu den großen fleischfressenden Dinosauriern (Theropoden) des Jura und der Kreide auf, insbesondere im Schädelbau. In beiden Fällen beruhen diese Ähnlichkeiten auf Konvergenz.[5]
Die Crocodylomorpha und die Crocodyliformes – erste Krokodile

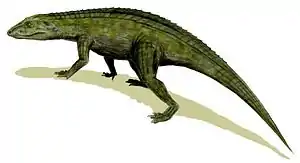
Zu den ersten Vertretern der Crocodylomorpha gehören die Sphenosuchia. Sie tauchten in der oberen Trias auf, also vor etwa 230 Millionen Jahren, und lebten bis zum späten Jura. Sie waren langbeinige Tiere von etwa einem Meter Länge, lebten terrestrisch und waren fakultativ biped, das heißt, sie liefen wenigstens zeitweilig auf zwei Beinen. Von den frühen Crocodylomorpha spalten sich die Crocodyliformes ab. Diese können als „Krokodile im weiteren Sinne“ gesehen werden. Sie enthalten ausschließlich vierbeinig laufende (obligat quadrupede) Vertreter, mit mehr oder weniger krokodiltypischem Aussehen. Die Crocodylia oder „Krokodile im engeren Sinne“ sind jene Untergruppe der Crocodyliformes, die alle rezenten Krokodile sowie deren nächste ausgestorbene Verwandte umfasst (Krokodil-Kronengruppe).
Einer der ältesten bekannten Vertreter der Crocodyliformes war Protosuchus, der sich durch lange Beine auszeichnete und wahrscheinlich ein ziemlich schneller Jäger war. Nach ihm wurde eine Gruppe der frühen Krokodile benannt, die Protosuchia. Ähnlich sah auch der nahe verwandte Orthosuchus aus. Diese Tiere waren noch auf eine Lebensweise auf dem Land eingestellt, während fast alle folgenden Gruppen zu einem amphibischen Leben im Wasser übergingen. Ein zweireihiger Knochenpanzer schützte den Rücken, auch der Bauch war verknöchert. Die Protosuchia waren bis ins frühe Jura auf dem damaligen Superkontinent Pangaea, in dem alle Festlandsmassen vereint waren, weit verbreitet, heutige Funde stammen entsprechend aus Ostasien, Europa, Nord- und Südamerika sowie aus Südafrika.
Die Mesoeucrocodylia


Kurz nach dem Beginn des Jura und dem damit verbundenen Zerbrechen des Urkontinents Pangaea entwickelte sich eine weitere Gruppe der Crocodyliformes, die heute als Mesoeucrocodylia bezeichnet wird. Dieses Taxon ersetzt das früher gebräuchliche paraphyletische Taxon Mesosuchia. Als monophyletisches Taxon enthalten die Mesoeucrocodylia auch die Eusuchia und stellen somit eine geschlossene Gruppe (Taxon) dar, die neben mehreren weiteren Entwicklungslinien auch die heutigen Krokodile (Crocodylia) enthält. Zu den ersten Vertretern der Mesoeucrocodylia gehören die Thalattosuchia, sie traten im frühen Jura vor etwa 190 Millionen Jahren auf und wurden vor allem in Europa gefunden. Diese ältesten Formen waren offensichtlich Meeresbewohner, da man sie in marinen Ablagerungen fand. Eine ihrer Familien waren die Teleosauridae. Diese Tiere zeichnen sich besonders durch die spezialisierten, lang gezogenen Kiefer mit den langen und spitzen Zähnen aus, die zum Fischfang eingesetzt werden konnten. Die Vorderbeine waren verkürzt, konnten jedoch auch an Land eingesetzt werden. Wie die Protosuchia hatten sie außerdem einen Plattenpanzer. Unter ihnen entwickelten sich Formen mit bis zu 10 Metern Länge wie Machimosaurus. Noch stärker an die Lebensweise im Meer angepasst war die spätere zweite Familie, die Metriorhynchidae aus dem Mitteljura. Bei ihnen entwickelten sich die Gliedmaßen zu Flossen um, und der Schwanz wurde zu einer starken Schwanzflosse, die den Tieren einen noch besseren Vortrieb verlieh. Aufgrund der besonderen Anpassungen werden diese Arten gemeinsam mit anderen marinen Krokodiltaxa als Meereskrokodile bezeichnet. Sie starben in der Frühen Kreide aus. Die Thalattosuchia werden alternativ auch als Mitglieder der Neosuchia in die Nähe der Tethysuchia gestellt.
Die Metasuchia





_1.jpg.webp)

| † Notosuchia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ••• |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Notosuchia (nach Pol und Kollegen, 2014).[7]
Im frühen Jura traten die Metasuchia als weitere Gruppe der Mesoeucrocodylia auf. Dieses Taxon wird nicht von allen Paläontologen als notwendig betrachtet und daher manchmal als Synonym der Mesoeucrocodylia gesehen. Die Sichtweise hängt besonders von der jeweils angenommenen Stellung der Thalattosuchia ab. Die große Gruppe der Metasuchia teilte sich früh in die Notosuchia und die Neosuchia auf. Bislang besteht aber große Uneinigkeit, an welchem Punkt genau die Trennung der beiden Gruppen erfolgt ist. Zu den in der frühen Kreide auftauchenden Notosuchia gehören die auf den Südkontinenten lebenden Vertreter der Uruguaysuchidae wie etwa die kleinen Arten der Gattung Araripesuchus. Sie waren in Südamerika und Afrika weit verbreitet. Zur gleichen Zeit wie die Uruguaysuchidae lebten die größeren Trematochampsidae, die mehr amphibisch waren und die Flüsse besiedelten. Ihre genaue verwandtschaftliche Einordnung ist noch umstritten. Möglicherweise stehen sie den Peirosauridae nahe. Eine andere Linie der Notosuchia sind die Ziphosuchia aus der späten Kreide. Zu ihr gehören die Libycosuchidae aus Afrika und die landlebenden Notosuchidae aus Südamerika. Weitere Ziphosuchia sind die ebenfalls landlebenden Sebecosuchia der Südkontinente. Zu Beginn des Tertiärs lebten sie als riesige Räuber mit einem speziell zum Zerschneiden von Fleisch konstruierten Gebiss, ähnlich dem fleischfressender Dinosaurier. Wichtige Gruppen stellten dabei die Sebecidae und die Baurusuchidae dar. Die Sebecidae lebten auch in Afrika und Europa, starben hier jedoch im Eozän, der frühesten Epoche des Tertiärs, wieder aus. In Südamerika waren sie dagegen bis zum Ende des Tertiärs die herrschenden Raubtiere. Der Grund war die damalige Isolation des südamerikanischen Kontinents vom Rest der Welt, so dass fleischfressende Säugetiere keine Konkurrenz für sie darstellten.
Die Neosuchia
 Lebendrekonstruktion von Stomatosuchus inermis aus der späten Kreide von Ägypten
Lebendrekonstruktion von Stomatosuchus inermis aus der späten Kreide von Ägypten Skelett von Bernissartia fagesii aus der späten Kreide von Belgien
Skelett von Bernissartia fagesii aus der späten Kreide von Belgien Rekonstruierter Schädel von Sarcosuchus imperator aus der frühen Kreide vom Nord-Niger, Afrika
Rekonstruierter Schädel von Sarcosuchus imperator aus der frühen Kreide vom Nord-Niger, Afrika Lebendrekonstruktion von Elosuchus cherifiensis aus der frühen Kreide von Nordafrika
Lebendrekonstruktion von Elosuchus cherifiensis aus der frühen Kreide von Nordafrika Schädel von Teleorhinus bzw. Terminonaris robusta aus der späten Kreide von Montana, USA
Schädel von Teleorhinus bzw. Terminonaris robusta aus der späten Kreide von Montana, USA Lebendrekonstruktion von Dyrosaurus phosphaticus aus dem Eozän von Nordafrika
Lebendrekonstruktion von Dyrosaurus phosphaticus aus dem Eozän von Nordafrika Lebendrekonstruktion von Goniopholis simus aus der frühen Kreide von Deutschland
Lebendrekonstruktion von Goniopholis simus aus der frühen Kreide von Deutschland
Systematik
| Neosuchia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ••• † Goniopholididae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Neosuchia (nach de Andrade und Kollegen, 2011).[8]
Die ersten Neosuchia lebten im frühen Jura. Etwas später traten die eher kleinen Atoposauridae des oberen Jura und der unteren Kreide auf, die ausschließlich in Laurasia vorkamen, unter ihnen Theriosuchus pusillus aus dem heutigen Großbritannien. Diese halb im Wasser, halb an Land lebenden Tiere gehören zu einer Entwicklungslinie, zu der auch die späteren modernen Krokodile, die Eusuchia, gehören. Das Erkennungsmerkmal sind die Wirbel, die hier erstmals als procoele Wirbelkörper mit einer vorderen Einbuchtung auftraten, während alle davor existierenden Arten amphicoele bzw. bikonkave Wirbelkörper hatten, also Wirbel mit einer vorderen und einer hinteren Vertiefung. Näher verwandt mit den Eusuchia waren die ebenfalls zu den Neosuchia gehörenden Goniopholididae des späten Jura. Diese Fluss- und Seebewohner lebten nach bisherigen Fossilfunden in Nordamerika, Europa und Thailand ausschließlich auf dem damaligen Nordkontinent Laurasia. Es handelte sich dabei um große Krokodile mit stumpfer Schnauze. Von dieser Entwicklungslinie spalteten sich zuvor die Stomatosuchidae ab. Obwohl der Fossilbefund der nachfolgenden Zeit relativ spärlich ist, lässt sich vermuten, dass sich die Neosuchia recht schnell ausbreiteten. So gab es einen Fund aus der späten Kreide im heutigen Ägypten, der zur Gruppe der Stomatosuchidae gehört. Diese Art wird als Stomatosuchus bezeichnet und hatte eine fast entenschnabelähnliche Schnauze. Die Eusuchia stammen aus einer Linie, die sich kurz zuvor abgespalten hat. Ein anderer Zweig der Neosuchia sind die Tethysuchia; zu ihnen gehören die schmalschnauzigen Pholidosauridae, darunter der aus Afrika stammende Sarcosuchus imperator mit einem Schädel von zwei Metern Länge und einer Gesamtlänge von etwa 11 Metern. In der späten Kreide entwickelten sich besonders langschnäuzige Arten wie Terminonaris (Teleorhinus), die wiederum ins Meer gingen. Gleichzeitig lebten an den Küsten der Tethys Afrikas und Südamerikas weitere Tethysuchia, die sich dem Leben und Jagen im Meer angepasst hatten. Diese Dyrosauridae, unter ihnen Dyrosaurus, waren ebenfalls langschnäuzig und so auf den Fischfang spezialisiert. Wie viele ihrer Verwandten überlebten sie das Massenaussterben an der Kreide-Tertiär-Grenze, starben jedoch zum Ende des Eozäns wahrscheinlich aufgrund der Konkurrenz mit meereslebenden Gavialen und frühen Walen aus.
Die Eusuchia – die „modernen“ Krokodile




| Eusuchia |
| |||||||||||||||||||||||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Eusuchia (nach Buscalioni und Kollegen, 2011).[9]
| Eusuchia |
| ||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Eusuchia (nach de Andrade und Kollegen, 2011).[8]
| Eusuchia |
| ||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Eusuchia (nach Holliday & Gardner, 2012).[10]
| Eusuchia |
| ||||||||||||||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Eusuchia (nach Bronzati, Montefeltro & Langer, 2012).[11]
In der frühen Kreidezeit traten die ersten Vertreter der Eusuchia auf. In der späten Kreide stellen diese bereits die dominierende Gruppe der Krokodile dar. Über den Punkt, ab wann man von Eusuchia sprechen kann und wann man noch von Neosuchia spricht, besteht noch Uneinigkeit. Die wichtigste Gruppe der Eusuchia sind die Crocodylia. Auch hier ist die Abgrenzung der Crocodylia zu den restlichen Eusuchia noch nicht vollständig geklärt. Sehr nahe Verwandte der Crocodylia sind die Aegyptosuchidae. Aegisuchus aus dem heutigen Marokko ist einer ihrer Vertreter.
Die Crocodylia – die „eigentlichen“ Krokodile




| Crocodylia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ••• |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Crocodylia (nach Martin, 2007[12]; Martin, 2010[13]; Martin & Gross, 2011[14]).
| Crocodylia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ••• Alligatoridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ••• Crocodylidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verwandtschaftsverhältnisse der Crocodylia (nach Brochu, 2001[15]; Brochu, 2003[16]; Vélez-Juarbe, Brochu & Santos, 2007[17]; Brochu und Kollegen, 2010[18]; Brochu, 2011[19]; Brochu und Kollegen, 2012[20]; Brochu & Storrs, 2012[21]).
Vor 80 Millionen Jahren existierten bereits Vertreter der Eigentlichen Krokodile (Crocodylia). Deinosuchus aus Nordamerika war einer der frühen Vertreter der Alligatoroidea, eines Taxons, das die näheren Verwandten der Alligatoridae mit einschließt; auch Vertreter der Crocodyloidea, eines Taxons, das die näheren Verwandten der Crocodylidae mit einschließt, kamen in dieser Zeit vor allem im heutigen Nordamerika vor. Die direkte Ahnenlinie dieser beiden Taxa, die dann auch zu den Echten Krokodilen (Crocodylidae) und den Alligatoren (Alligatoridae) führt, liegt noch etwas im Dunklen, als einzige fossile Art, die Merkmale beider Gruppen aufweist, gilt Mekosuchus inexpectatus, ein Vertreter der Mekosuchinae. Es handelte sich dabei um eine landlebende Art, die in Neukaledonien bis in die historische Zeit vor etwa 3500 bis 3900 Jahren gelebt hat. Ebenfalls zu den Mekosuchinae gehörte das Landkrokodil Quinkana fortirostrum. Fast während des gesamten Tertiärs waren sowohl Crocodyloidea als auch Alligatoroidea über alle nördlichen Kontinente weit verbreitet. Die Crocodyloidea und die Alligatoroidea werden manchmal auch als Brevirostres zusammengefasst. Auch in Europa gab es Vertreter dieser beiden Gruppen. Eine bekannte Form der Alligatoroidea aus Europa ist Diplocynodon, ein Vertreter der Diplocynodontinae. Ebenfalls in Europa lebten die Pristichampsidae, eine Gruppe von Krokodilen mit hufähnlichen Zehen.
Vertreter der Crocodylinae, zu denen auch die heutigen Gattungen und Arten der Krokodile gehören, traten seit dem frühen Tertiär (Eozän oder Oligozän) erstmals auf. So fand man Fossilien des Nilkrokodils (Crocodylus niloticus) und des westafrikanischen Panzerkrokodil (Mecistops cataphractus) zum Ende des Tertiärs und im Pleistozän, mit Rimasuchus lloidi wurde allerdings ein noch früherer Vertreter der afrikanischen Crocodylinae gefunden. Aus Asien ist außerdem Crocodylus palaeindicus bekannt. Von der Entstehung der weiteren Arten der Gattung in der ozeanischen Inselwelt und Südamerika sowie vom ersten Auftreten des Stumpfkrokodils (Osteolaemus tetraspis) und des Sunda-Gavials (Tomistoma schlegelii) ist nur wenig bekannt.
Auch die Evolution der heutigen Alligatoren und Kaimane (Alligatoridae) ist nur sehr lückenhaft dokumentiert. Als Ahne des Mississippi-Alligators (Alligator mississippiensis) gilt Alligator olseni aus dem Miozän. Erste Nachweise des China-Alligators (Alligator sinensis) stammen aus dem Pleistozän. Ein früher Vertreter der Kaimane (Caimaninae) war Eocaiman cavernensis. Im Miozän lebte Purussaurus neivensis.
Weitgehend ungeklärt ist der Ursprung der Gaviale (Gavialidae). Diese stellen nach Ansicht einiger Forscher vielleicht sogar einen frühen Zweig der Mesoeucrocodylia dar, die bis in die heutige Zeit mit dem Gangesgavial überlebt haben. Weiter verbreitet ist allerdings die Ansicht, dass es sich bei ihnen um eine Schwestergruppe der beiden anderen modernen Krokodiltaxa handelt, die von langschnäuzigen Formen der Küstengewässer Nordafrikas abstammen. Von dort breiteten sich die frühen Gaviale nach Europa, Asien und Amerika aus und erfuhren in Südamerika eine Radiation, bevor sie, bis auf die einzige heute noch lebende Art in Indien, weltweit aus bislang ungeklärten Gründen ausstarben. Wahrscheinlich aufgrund der Klimaverschlechterung und Abkühlung zum Ende des Tertiärs verschwanden auch viele weitere Artengruppen aus den nördlichen Verbreitungsgebieten in Nordamerika, Asien und Europa.
Weblinks
- Crocodylomorpha: Overview (englisch)
Einzelnachweise
- siehe z. B.: Kerstin Schmidt-Denter: Dem Tod von der Schippe gesprungen – Einige „lebende Fossilien“ hatten Glück. scinexx.de, 12. April 2001.
- Phil Senter: Phylogenetic taxonomy and the names of the major archosaurian (Reptilia) clades. In: PaleoBios. Band 25, Nr. 3, 2005, S. 1–7 (Abstract).
- Sterling J. Nesbitt, Stephen L. Brusatte, Julia B. Desojo, Alexandre Liparini, Marco A. G. De França, Jonathan C. Weinbaum, David J. Gower: Rauisuchia. In: Sterling J. Nesbitt, Julia B. Desojo, Randall B. Irmis (Hrsg.): Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin. (= Geological Society, London, Special Publications. Band 379). Geological Society of London, London 2013, S. 241–274, doi:10.1144/SP379.1.
- Michael J. Benton: Paläontologie der Wirbeltiere. 3. Auflage (Übersetzung der 3. englischsprachigen Auflage durch Hans-Ulrich Pfretzschner), Verlag Dr. Friedrich Pfeil, München 2007, ISBN 3-89937-072-4, S. 139.
- Stephen L. Brusatte, Michael J. Benton, Julia B. Desojo, Max C. Langer: The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida). In: Journal of Systematic Palaeontology. Band 8, Nr. 1, S. 3–47, doi:10.1080/14772010903537732.
- Patrick M. O’Connor et al.: The evolution of mammal-like crocodyliforms in the Cretaceous Period of Gondwana. In: Nature. Band 466, 2010, S. 748–751, doi:10.1038/nature09061 (= Erstbeschreibung der Gattung Pakasuchus).
- D. Pol, P. M. Nascimento, A. B. Carvalho, C. Riccomini, R. A. Pires-Domingues, H. Zaher: A New Notosuchian from the Late Cretaceous of Brazil and the Phylogeny of Advanced Notosuchians. In: PLoS ONE. 9, Nr. 4, 2014, S. e93105. doi:10.1371/journal.pone.0093105. PMID 24695105.
- Marco Brandalise de Andrade, Richard Edmonds, Michael J. Benton, Remmert Schouten: A new Berriasian species of Goniopholis (Mesoeucrocodylia, Neosuchia) from England, and a review of the genus. In: Zoological Journal of the Linnean Society. 163, Nr. s1, 2011, S. S66–S108. doi:10.1111/j.1096-3642.2011.00709.x.
- A.D. Buscalioni, P. Piras, R. Vullo, M. Signore, C. Barbera: Early eusuchia crocodylomorpha from the vertebrate-rich Plattenkalk of Pietraroia (Lower Albian, southern Apennines, Italy). (PDF) In: Zoological Journal of the Linnean Society. 163, 2011, S. S199–S227. doi:10.1111/j.1096-3642.2011.00718.x.
- Casey M. Holliday, Nicholas M. Gardner: A New Eusuchian Crocodyliform with Novel Cranial Integument and Its Significance for the Origin and Evolution of Crocodylia. In: PLoS ONE. 7, Nr. 1, 2012, S. e30471. doi:10.1371/journal.pone.0030471. PMID 22303441. PMC 3269432 (freier Volltext).
- M. Bronzati, F. C. Montefeltro, M. C. Langer: A species-level supertree of Crocodyliformes. In: Historical Biology. 2012, S. 1. doi:10.1080/08912963.2012.662680.
- J. E. Martin: New material of the Late Cretaceous globidontan Acynodon iberoccitanus (Crocodylia) from Southern France. In: Journal of Vertebrate Paleontology. 27, Nr. 2, 2007, S. 362–372. doi:10.1671/0272-4634(2007)27[362:NMOTLC]2.0.CO;2.
- J. E. Martin: A new species of Diplocynodon (Crocodylia, Alligatoroidea) from the Late Eocene of the Massif Central, France, and the evolution of the genus in the climatic context of the Late Palaeogene. In: Geological Magazine. 147, 2010, S. 596–610. doi:10.1017/S0016756809990161.
- J. E. Martin, M. Gross: Taxonomic clarification of Diplocynodon Pomel, 1847 (Crocodilia) from the Miocene of Styria, Austria. In: Neues Jahrbuch für Geologie und Paläontologie – Abhandlungen. 261, Nr. 2, 2011, S. 177–193. doi:10.1127/0077-7749/2011/0159.
- C. A. Brochu: Crocodylian snouts in space and time: phylogenetic approaches toward adaptive radiation. In: American Zoologist. 41, Nr. 3, 2001, S. 564–585. doi:10.1668/0003-1569(2001)041[0564:CSISAT]2.0.CO;2.
- C. A. Brochu: Phylogenetic approaches toward crocodylian history. In: Annual Review of Earth and Planetary Sciences. 31, 2003, S. 357–97. doi:10.1146/annurev.earth.31.100901.141308.
- J. Vélez-Juarbe, C. A. Brochu, H. Santos: A gharial from the Oligocene of Puerto Rico: transoceanic dispersal in the history of a non-marine reptile. In: Proceedings of the Royal Society B. 274, Nr. 1615, 2007, S. 1245–1254. doi:10.1098/rspb.2006.0455. PMID 17341454. PMC 2176176 (freier Volltext).
- C. A. Brochu, J. Njau, R. J. Blumenschine, L. D. Densmore: A New Horned Crocodile from the Plio-Pleistocene Hominid Sites at Olduvai Gorge, Tanzania. In: PLoS ONE. 5, Nr. 2, 2010, S. e9333. doi:10.1371/journal.pone.0009333. PMID 20195356. PMC 2827537 (freier Volltext).
- C. A. Brochu: Phylogenetic relationships of Necrosuchus ionensis Simpson, 1937 and the early history of caimanines. In: Zoological Journal of the Linnean Society. 163, 2011, S. S228–S256. doi:10.1111/j.1096-3642.2011.00716.x.
- Christopher A. Brochu, David C. Parris, Barbara Smith Grandstaff, Robert K. Denton, William B. Gallagher: A new species of Borealosuchus (Crocodyliformes, Eusuchia) from the Late Cretaceous-early Paleogene of New Jersey. Band 32, Nr. 1, 2012, ISSN 0272-4634, S. 105–116, doi:10.1080/02724634.2012.633585.
- C. A. Brochu, G. W. Storrs: A giant crocodile from the Plio-Pleistocene of Kenya, the phylogenetic relationships of Neogene African crocodylines, and the antiquity of Crocodylus in Africa. In: Journal of Vertebrate Paleontology. 32, Nr. 3, 2012, S. 587–602. doi:10.1080/02724634.2012.652324.