Carboxysom
Carboxysomen sind bakterielle Mikrokompartimente (englisch bacterial microcompartments, BMC), die aus einer polyedrischen Proteinhülle (englisch shell ‚Schale‘) bestehen und die Enzyme Ribulose-1,5-bisphosphat-carboxylase/-oxygenase (RuBisCO – das vorherrschende Enzym bei der Kohlenstofffixierung und geschwindigkeitsbeschränkende Enzym im Calvin-Zyklus) und Carboanhydrase gefüllt sind.[2]
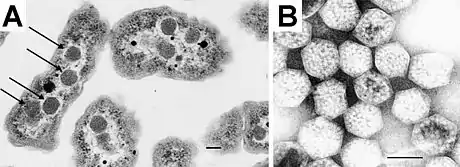
(A) angeordnet in der Zelle und (B) intakt nach der Isolierung.
Balken jeweils 100 nm.[1]
In der RuBisCO-Reaktion ist Sauerstoff ein konkurrierendes Agens zu Kohlendioxid. Daher geht man davon aus, dass sich Carboxysomen als Folge des Anstiegs der Sauerstoffkonzentration in der frühen Erdatmosphäre entwickelt haben.[3] Um die Ineffizienz von RuBisCO zu überwinden, konzentrieren Carboxysomen Kohlendioxid innerhalb ihrer Schale vermöge der Aktivität der dortigen Carboanhydrase. Diese erzeugt Kohlendioxid aus Bikarbonat, das in das Carboxysom diffundiert. Die daraus resultierende Konzentration von Kohlendioxid in der Nähe von RuBisCO verringert den Anteil der Ribulose-1,5-bisphosphat-Oxygenierung und vermeidet dadurch kostspielige photorespiratorische Reaktionen. Die umgebende Schale stellt eine Barriere für den Verlust von Kohlendioxid dar und trägt dazu bei, dessen Konzentration in der Nähe von RuBisCO hoch zu halten bzw. zu erhöhen.[4][5] Carboxysomen sind daher ein wesentlicher Bestandteil des Kohlendioxid-Konzentrationsmechanismus (en. carbon dioxide-concentrating mechanism, CCM).
Carboxysomen sind das am besten untersuchte Beispiel für bakterielle Mikrokompartimente, d h. für Organellen unterschiedlicher Funktion, die sich durch eine Proteinhülle oder -schale auszeichnen.[6][7]
Entdeckung und Forschungsgeschichte
Die Carboxysomen wurden 1956 per Transmissionselektronenmikroskopie (TME) im Cyanobakterium Phormidium uncinatum als Polyederkörperchen entdeckt.[8] Später wurden sie auch in anderen Cyanobakterien[9] sowie in einer Reihe chemotropher (Kohlendioxid fixierender) Bakterien beobachtet, von denen viele Schwefeloxidierer oder Stickstofffixierer sind (z. B. Halothiobacillus, Acidithiobacillus, Nitrobacter und Nitrococcus; alle gehören zu den Proteobakterien).[2][10] Die polyedrischen Körper wurden erstmals 1973 aus Halothiobacillus neapolitanus (damals noch als Thiobacillus neapolitanus bezeichnet) isoliert und es zeigte sich, dass sie RuBisCO in einer m. o. w. starren äußeren Schale enthielten.[11] Die Autoren schlugen vor, sie als Carboxysomen zu bezeichnen, da es sich offenbar um Organellen handelt, die an der Kohlendioxid-Fixierung beteiligt sind.[11]
Die eukaryontischen Glaucophyta besitzen Cyanellen genannten Plastiden, die offenbar aus endosymbiotischen Cyanobakterien entstanden sind und in vieler Beziehung ursprünglicher erscheinen als die Chloroplasten der Grünalgen und Landpflanzen. Auch bei diesen Cyanellan fanden sich carboxysom-artige Körperchen (auch Zentralkörperchen, englisch central bodies, genannt), Analoga der Carboxysomen.[12][13] Ob es sich dabei tatsächlich um Carboxysom-Abkömmlinge handelt, wurde aber auch infrage gestellt.[14] Die ähnlichen Chromatophoren von Paulinella chromatophora zeigen im elektronenmikroskopischen Bild zehn bis zwanzig konzentrisch angeordnete, etwas wellenförmige Thylakoide, mit eingelagerten Phycobilisomen und Carboxysomen.[15]
Aufbau
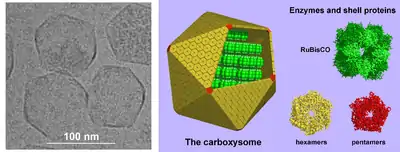
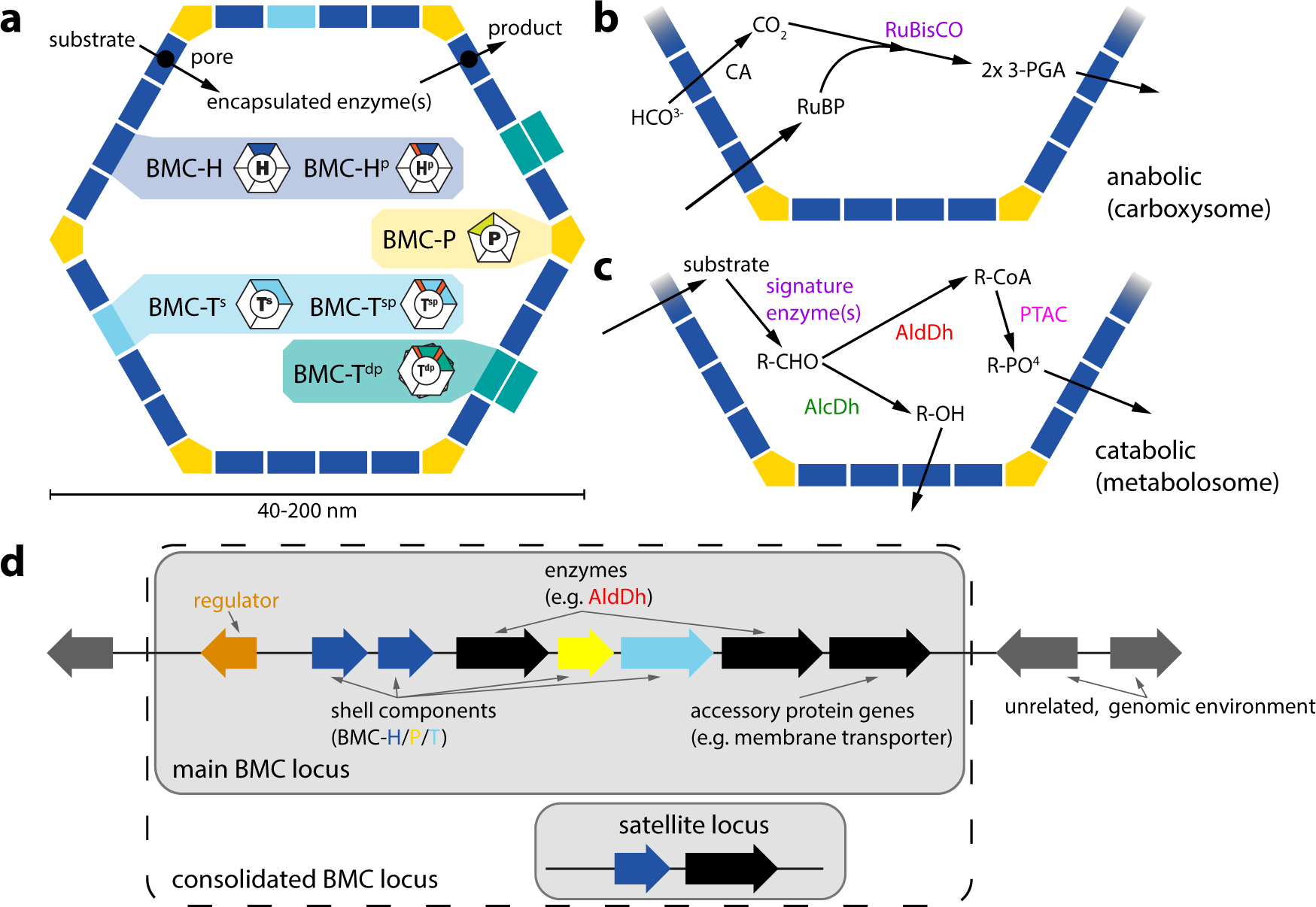
Rechts: Modelle für die Struktur des Carboxysoms. Die Schale besteht aus mehreren hundert hexameren und 12 pentameren Proteinbausteinen. Die 3D-Darstellunen dieser Schalenproteine wurden per Röntgenkristallographie bestimmt. Das wichtigste Enzym im Inneren, RuBisCO, ist vereinfacht in einer regelmäßigen Anordnung dargestellt, obwohl die tatsächliche Organisation dieser Enzyme komplizierter ist. Das andere Schlüsselenzym, die Carboanhydrase, ist in geringeren Mengen vorhanden (nicht dargestellt).
Die Carboxysomen haben eine ikosaedrische oder quasi-ikosaedrische Symmetrie. Elektronen-Kryotoomographie-Studien haben diese Geometrie des Carboxysoms bestätigt[18][19][20] und auch Proteinmoleküle im Inneren (vermutlich RuBisCO) abgebildet, die in einigen konzentrischen Schichten angeordnet sind.[18][20] Die von dieser (annähernd) ikosaedrischen Geometrie abweichende Facettenform einiger Carboxysomen lässt sich auf natürliche Weise mit einer Theorie der elastischen heterogenen dünnen Schalen erklären.[21] Das Carboxysom hat eine äußere Schale, die aus einigen Tausend Proteinuntereinheiten besteht und die ein CO2-produzierendes Enzym (Carboanhydrase) und ein kohlenstoffbindendes Enzym (RuBisCO) einkapselt. Die Struktur der Schalenproteine wurde durch Röntgenkristallographie aufgeklärt. Diese Proteine bilden zyklische Hexamere oder Pseudo-Hexamere und gehören zur BMC-Proteinfamilie (en. Bacterial microcompartment).[17] Meist habe die BMC-H-Hexamere kleine Poren, die offenbar einen Diffusionsweg für kleine Substrate (z. B. Bicarbonat) und erzeugte Produkte (z. B. 3-Phosphoglycerat) in das bzw. aus dem Carboxysom dienen. Positiv geladene Aminosäuren in den Poren tragen vermutlich dazu bei, die Diffusion der negativ geladenen Substrate und Produkte zu fördern.[17] Zu den anderen identifizierten kleineren strukturellen Komponenten der Schale gehören die pentameren Proteine BMC-P, die die Eckpunkte der ikosaedrischen Schale besetzen.[22] Ein dritter Baustein der Carboxysomenschale ist ein Protein, das aus zwei BMC-Domänen in Tandemanordnung besteht (BMC-T). Diese Proteine bilden Trimere, die (im 2er-Tandem) pseudohexamer sind.[23][24] Einige Mitglieder der BMC-T-Proteinfamilie stapeln sich quasi von „Angesicht zu Angesicht“ und bilden auf diese Weise winzige Käfige, die sich bei beiden Typen der Carboxysomen finden lassen.[23][24] Wie die Kristallstrukturenanalyse zeigt, haben diese Proteinkäfige relativ große Poren auf beiden Seiten, die gewissermaßen Tore darstellen (engl. gated pores). Man vermutet, dass das Öffnen und Schließen dieser Poren ähnlich wie bei einer Luftschleuse gesteuert werden könnte. Im Gegensatz zu BMC-H-Proteinen mit konstitutiv offenen Poren dient eine solche Schleuse anscheinend als Weg für größere Substrate (wie Ribulose-1,5-bisphosphat) und Produkte (3-Phosphoglycerat), die die Käfigschale durchqueren müssen.[23][24]
Die Herstellung von leeren Carboxysomenschalen in E. coli ermöglichte die erste Visualisierung dieser Strukturen durch Kryoelektronenmikroskopie (Cryo-EM).[25]
Typen von Carboxysomen
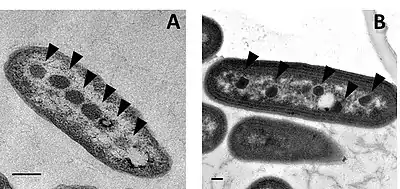
Balken jeweils 200 nm.
Es gibt zwei Arten von Carboxysomen. Obwohl sie sich äußerlich ähneln, unterscheiden sie sich in der Zusammensetzung der Proteine, die sie umschließen, einschließlich der Form des RuBisCO.[26][27][28][29] Darüber hinaus haben Studien grundlegende Unterschiede in ihrer Genorganisation und möglicherweise auch in ihrem Zusammensetzungsweg (en. assembly pathway, siehe Assemblierung) ergeben.
Alpha-Carboxysomen
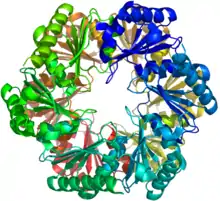
Alpha-Carboxysomen (auch α-Carboxysomen geschrieben) werden auch als cso-Typ von Carboxysomen bezeichnet. Sie enthalten RuBisCO der Form IA und kommen vor in
- Alpha-Cyanobakterien (α-Cyanobacteria, alias Synechococcus/Prochlorococcus/Cyanobium-Klade[13] beispielsweise Cyanobium sp. PCC 7001[30]),[31]
- einigen nitrifizierenden Bakterien
- einigen schwefeloxidierenden Bakterien (wie Halothiobacillus neapolitanus) und
- einigen Purpurbakterien
Alle diese werden – bis auf die α-Cyanobakterien – taxonomisch den Proteobakterien zugeordnet. Das Alpha-Carboxysom war das erste bakterielle Mikrokompartiment, das gereinigt und charakterisiert wurde[32][33] Elektronenmikroskopische Untersuchungen an gereinigten Alpha-Carboxysomen, sowie von Zellschnitten die diese enthalten, ergaben, dass diese typischerweise einen Durchmesser von 100–160 nm haben.[34] Die gemeinsamen Bausteine für die Schale von Alpha-Carboxysomen werden CsoS1A/B/C (BMC-H), CsoS4A/B (BMC-P) und CsoS1D (BMC-T) genannt. CsoS4A/B waren die ersten BMC-P-Proteine, die experimentell als Nebenkomponenten der BMC-Schale nachgewiesen wurden. Es sind nur 12 Pentamere erforderlich, um die Spitzen eines Ikosaeders zu bedecken.[4] CsoS1D ist das erste BMC-T-Protein, dessen Struktur beschrieben wurde; es ist auch das erste Beispiel für die Dimerisierung von zwei BMC-Bausteinen, die sich gegenüberstehen und einen winzigen Käfig mit den Poren an beiden Enden, die (wie vermutet) den Transfer großer Metaboliten durch die Schale erleichtern.[24] Neben der spezifischen Form von RuBisCO zeigen auch andere eingekapselte Proteine wie CsoS2 und CsoSCA Unterschiede zwischen α- und β-Carboxysomen. Das CsoS2-Protein hat einen sehr hohen isoelektrischen Punkt (pI) und eine einzigartige Primärstruktur. Diese scheint dreiteilig zu sein und besteht aus einer N-terminalen, einer mittleren und einer C-terminalen Region.[35] In der N-terminalen und der mittleren Region können sich wiederholende Sequenzmotive identifiziert werden.[35] Man hat vorgeschlagen, dass es sich hier um ein intrinsisch ungeordnetes Protein handelt, das eine wesentliche Rolle beim Aufbau von α-Carboxysomen spielt. CsoSCA ist eine schalenassoziierte β-Carboanhydrase (β-CA, en. beta-carbonic anhydrase).[5][36] Studien an Halothiobacillus neapolitanus haben gezeigt, dass leere Schalen von normaler Form und Zusammensetzung auch in Mutanten aufgebaut werden, denen das carboxymale RuBisCO fehlt. Dies deutet darauf hin, dass die Biogenese der α-Carboxysomenschale und die Sequestrierung (Einlagerung, vgl. Internalisierung) des Enzyms zwei unabhängige, aber funktionell miteinander verbundene Prozesse sind.[37] Es ist die große Untereinheit von RuBisCO, die darüber entscheidet, ob das Enzym in Carboxysomen sequestriert (internalisiert) wird. Interessant ist weiter, dass die Carboxysomen von Halothiobacillus neapolitanus chimäre und heterologe Arten von RuBisCO beherbergen.[37]
Beta-Carboxysomen
Beta-Carboxysomen (auch β-Carboxysomen genannt) kommen in Beta-Cyanobakterien (β-cyanobacteria) wie beispielsweise Synechococcus sp. PCC 7942 vor.[31][38]
Die charakteristischen Proteine des Beta-Carboxysoms sind das RuBisCO mit Form und ein Homolog der γ-Carboanhydrase (γ-CA, en. gamma-carbonic anhydrase). Beta-Carboxysomen sind typischerweise größer als Alpha-Carboxysomen, die beobachteten Durchmesser variieren hier zwischen 200 und 400 nm.[35] Die für die Bildung von Beta-Carboxysomen wesentlichen Strukturproteine werden in dem konservierten Carboxysom-Locus (ccm-Locus) kodiert. Der ccm-Locus umfasst Gene für die Kernproteine CcmM und CcmN sowie die Schalenproteine CcmK (ein BMC-H-Protein), CcmL (ein BMC-P-Protein) und CcmO (ein BMC-T-Protein).[7]
Ein CcmM-Protein in voller Länge besteht aus einer γ--Carboanhydrase-Domäne und (an seinem C-Terminus) drei bis fünf RuBisCO small subunit-like domains (SSLDs).[39] Das ccmM-Gen enthält eine interne Translationsstelle (en. internal translation site), die eine kurze Form von CcmM (nur aus SSLDs bestehend) produziert. Sowohl die lange als auch die kurze Form von CcmM sind für den Aufbau des β-Carboxysoms erforderlich.[40] Das Protein CcmN enthält mehrere Hexapeptid-Wiederholungsdomänen an seinem N-Terminus und ein kurzes α-helicales Verkapselungspeptid am C-Terminus.[41]
Andere strukturelle Komponenten von Beta-Carboxysomen werden außerhalb des ccm-Locus kodiert. CcmP ist ein BMC-T-Protein, das bei Organismen, die β-Carboxysomen bilden, absolut konserviert ist. Zwei CcmP-Pseudohexamere stapeln sich zu einem Nanokompartiment – ein Beispiel für ein schleusenbildendes Protein (en. air-lock forming protein).[23] Die β-Carboxysomen einiger Cyanobakterienstämme enthalten auch eine β-Carboanhydrase, die ebenfalls nicht im ccm-Locus kodiert wird.[42]
Die Schalenproteine der β-Carboxysomen sind im Vergleich zu ihren Gegenstücken in den α-Carboxysomen relativ vielfältig.[38] Da die sie enthaltenden Cyanobakterien sich in ökophysiologisch dynamischen (wechselhaften) Umgebungen aufhalten, vermutet man, dass dies die variablen Anforderungen an die Permeabilität (Durchlässigkeit für Substanzen) der β-Carboxysomen widerspiegelt.[43]
Der Zusammenbau (Assemblierung) eines β-Carboxysom erfolgt von innen nach außen wie folgt: Zunächst bildet sich ein Kern (en. carboxysome core) aus Enzymen, der anschließend von der Proteinschale eingekapselt wird.[44] Der Aufbau des Carboxysoms erfolgt durch eine Reihe von Protein-Protein-Wechselwirkungen: Das Enzym RuBisCO und die beiden Isoformen (Vollform und Kurzform) des CcmM-Proteins interagieren über die SSLDs. In Stämmen, die CcaA enthalten, wird die β-Carboanhydrase durch Wechselwirkung mit dem N-Terminus des Vollform-CcmM in den Carboxysom-Kern gebracht.[45][46] Sobald das Procarboxysom (d. h. der Carboxysom-Kern als Carboxysom-Vorläufer) gebildet ist, interagiert der N-Terminus des Adapterproteins CcmN mit dem N-Terminus von CcmM, während der C-Terminus von CcmN die Schalenproteine CcmK (BMC-H) und CcmO (BMC-T) rekrutiert, wozu ein 15–20 Aminosäuren langes Peptid verwendet wird.[41] Dieses Verkapselungspeptid bildet eine amphipathische α-Helix (siehe α-Helix §Geometrie der Helix und Helix-Helix-Wechselwirkungen), die mit den Schalenkomponenten interagiert. Seine Rolle ist von wesentlicher Bedeutung, da ohne dieses Peptid keine Carboxysomen gebildet werden können.[41][29] Der letzte Schritt ist die Hinzufügung der vom BMC-P-Protein CcmL gebildeten Eckpunkte, die dann den enzymatischen Kern und die Facetten (Seitenflächen des Ikosaeders) wie Kappen zusammenhalten.[44] Die Aufklärung des Zusammensetzungsweges von β-Carboxysomen ermöglichte die Entwicklung eines einzigen synthetischen Proteins, das vier andere Proteine bei der Carboxysomenbildung ersetzt.[47]
Mögliche Anwendungen
Wie auch andere BMCs zieht das Carboxysom die Aufmerksamkeit von Forschern für Anwendungen in der synthetischen Biologie von Pflanzen auf sich.[48][26][49] Durch die Übertragung eines genetischen Moduls aus 10 Genen, das für ein α-Carboxysom kodiert, von Halothiobacillus neapolitanus konnten in E.coli Carboxysom-ähnliche (ikosaedrische und funktionale) Strukturen erzeugt werden.[50] Das Bioengineering von Carboxysom-Schalen hat sich als machbar erwiesen, und es wurde von β-Carboxysomen berichtet, die mit chimären Proteinen oder mit chimären Hüllen konstruiert wurden.[51] Die Einführung von Carboxysomen in pflanzliche Chloroplasten als Teil eines CO2-Konzentrationsmechanismus, wie er in Cyanobakterien vorkommt,[52][53] kann voraussichtlich die Netto-CO2-Fixierung und den Ertrag erheblich verbessern.[54][55] Beispielsweise konnte die Expression von β-carboxysomalen Schalenproteinen und RuBisCO-CcmM-Komplexen der Form IB in Tabak-Chloroplasten erreicht werden, führte aber nicht zu Kompartimenten, die RuBisCO enthalten.[56][57] Jedoch gelang die Konstruktion von minimalen α-Carboxysomen aus dem Cyanobakterium Cyanobium sp. PCC 7001,[30] die Form IA RuBisCO und die CsoS1A- und CsoS2-Proteine enthalten in den Tabak-Chloroplasten.[58] Allerdings wurden bisher (Stand 2017) wurden noch keine identifizierbaren funktionellen Carboxysomen in Pflanzen-Chloroplasten konstruiert. Die Verbesserung der Photosynthese in Pflanzen mit Hilfe dieses Ansatzes hängt letztlich von der Funktion von Transporterproteinen in der inneren Hüllmembran des Chloroplasten ab, die zur Erzeugung einer hohen Bikarbonatkonzentration innerhalb des Chloroplasten beitragen.[59]
Evolutionäre Herkunft
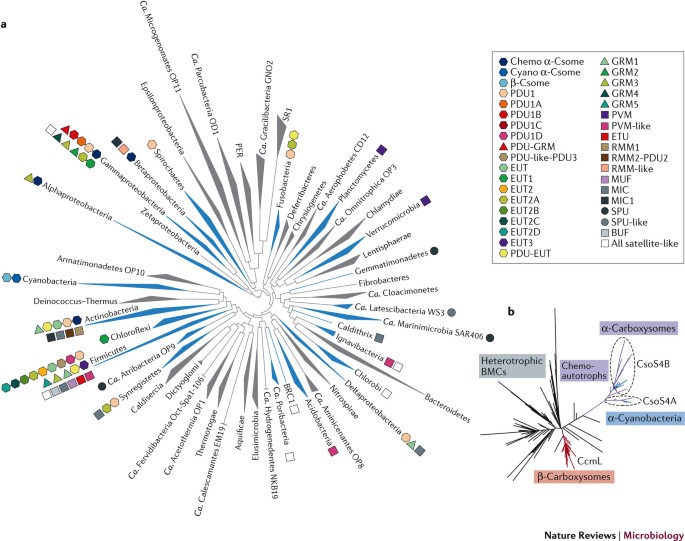
Eine Reihe von Viruskapsiden sind ebenfalls ikosaedrisch und bestehen aus hexameren und pentameren Proteinen, doch gibt es derzeit keine Hinweise auf eine evolutionäre Verwandtschaft zwischen Carboxysomenschalen und Viruskapsiden. Ausgehend von bioinformatischen Untersuchungen der Schalenproteine und des Zusammenbaus der Carboxysomen könnte es sogar sein, dass sich die beiden Typen unabhängig voneinander entwickelt haben.[60][29] Der Vorschlag einer Phylogenie der bakteriellen Mikrokompartiemente (BMCs), der außer den Carboxysomen auch heterotrophe und chemoautotrophe BMCs umfasst findet sich bei Kerfeld et al. (2018).[61]
Weblinks und Literatur
- Mysterious Bacterial Microcompartments Revealed By Biochemists. Auf ScienceNews vom 26. Februar 2008. Quelle: University of California, Los Angeles (UCLA)
- Philip Hunter: Not so simple after all: A renaissance of research into prokaryotic evolution and cell structure. In: EMBO Reports, Band 9, Nr. 3, März 2008, S. 224–226; doi:0.1038/embor.2008.24, PMC 2267389 (freier Volltext), PMID 18311171, Epub 8. Februar 2008.
- Images. Bildergalerie vom Kerfeld Lab, Cheryl A. Kerfeld et al.
- Markus Sutter, Matthew R. Melnicki, Frederik Schulz, Tanja Woyke, Cheryl A. Kerfeld: A catalog of the diversity and ubiquity of bacterial microcompartments. In: Nature Communications, Band 12, Nr. 3809, 21. Juni 2021, doi:10.1038/s41467-021-24126-4. Siehe insbes. Fig. 1.
- Henning Kirst, Cheryl A Kerfeld: Clues to the function of bacterial microcompartments from ancillary genes. In: Biochem Soc Trans., Band 49, Nr. 3, 1. Juli 2021, S. 1085–1098, doi:10.1042/BST20200632. Siehe insbes. BMCs: Carboxysom vs. Metabolosom
{kind=link}
Einzelnachweise
- Y. Tsai, M. R. Sawaya, G. C. Cannon et al.: Structural Analysis of CsoS1A and the Protein Shell of the Halothiobacillus neapolitanus Carboxysome. In: PLOS Biol.. 5, Nr. 6, Juni 2007, S. e144. doi:10.1371/journal.pbio.0050144. PMID 17518518. PMC 1872035 (freier Volltext).
- Todd O. Yeates, Cheryl A. Kerfeld, Sabine Heinhorst, Gordon C. Cannon, Jessup M. Shively: Protein-based organelles in bacteria: carboxysomes and related microcompartments. In: Nature Reviews Microbiology. 6, Nr. 9, 2008, ISSN 1740-1526, S. 681–691. doi:10.1038/nrmicro1913. PMID 18679172.
- M. R. Badger: CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. In: Journal of Experimental Botany. 54, Nr. 383, 2003, ISSN 1460-2431, S. 609–622. doi:10.1093/jxb/erg076. PMID 12554704.
- Fei Cai, Balaraj B. Menon, Gordon C. Cannon, Kenneth J. Curry, Jessup M. Shively, Sabine Heinhorst: The Pentameric Vertex Proteins Are Necessary for the Icosahedral Carboxysome Shell to Function as a CO2 Leakage Barrier. In: PLOS ONE. 4, Nr. 10, 2009, ISSN 1932-6203, S. e7521. bibcode:2009PLoSO...4.7521C. doi:10.1371/journal.pone.0007521. PMID 19844578. PMC 2760150 (freier Volltext).
- Z. Dou, S. Heinhorst, E. B. Williams, C. D. Murin, J. M. Shively, G. C. Cannon: CO2 Fixation Kinetics of Halothiobacillus neapolitanus Mutant Carboxysomes Lacking Carbonic Anhydrase Suggest the Shell Acts as a Diffusional Barrier for CO2. In: Journal of Biological Chemistry. 283, Nr. 16, 2008, ISSN 0021-9258, S. 10377–10384. doi:10.1074/jbc.M709285200. PMID 18258595.
- Cheryl A. Kerfeld, Onur Erbilgin: Bacterial microcompartments and the modular construction of microbial metabolism. In: Trends in Microbiology. 23, Nr. 1, 2015, ISSN 0966-842X, S. 22–34. doi:10.1016/j.tim.2014.10.003. PMID 25455419.
- Seth D. Axen, Onur Erbilgin, Cheryl A. Kerfeld: A Taxonomy of Bacterial Microcompartment Loci Constructed by a Novel Scoring Method. In: PLOS Computational Biology. 10, Nr. 10, 2014, ISSN 1553-7358, S. e1003898. bibcode:2014PLSCB..10E3898A. doi:10.1371/journal.pcbi.1003898. PMID 25340524. PMC 4207490 (freier Volltext).
- G. Drews, W. Niklowitz: Cytology of Cyanophycea. II. Centroplasm and granular inclusions of Phormidium uncinatum. In: Archiv für Mikrobiologie. 24, Nr. 2, 1956, S. 147–162. PMID 13327992.
- E. Gantt, S. F. Conti: Ultrastructure of blue-green algae. In: Journal of Bacteriology. 97, Nr. 3, März 1969, S. 1486–1493. doi:10.1128/JB.97.3.1486-1493.1969. PMID 5776533. PMC 249872 (freier Volltext).
- J. M. Shively: Inclusion Bodies of Prokaryotes. In: Annual Review of Microbiology. 28, Nr. 1, 1974, ISSN 0066-4227, S. 167–188. doi:10.1146/annurev.mi.28.100174.001123. PMID 4372937.
- J. M. Shively, F. Ball, D. H. Brown, R. E. Saunders: Functional Organelles in Prokaryotes: Polyhedral Inclusions (Carboxysomes) of Thiobacillus neapolitanus. In: Science. 182, Nr. 4112, 1973, ISSN 0036-8075, S. 584–586. bibcode:1973Sci...182..584S. doi:10.1126/science.182.4112.584. PMID 4355679.
- S. C. Burey, S. Fathi-Nejad, V. Poroyko, J. M. Steiner, W. Löffelhardt, H. J. Bohner: The central body of the cyanelles of Cyanophora paradoxa: a eukaryotic carboxysome?. Auf: Canadian Journal of Botany, Band 83, Nr. 7, Juli 2005, doi:10.1139/b05-060.
- Linda Oberleitner: Exploring transport processes across the symbiotic interface of amoebal host and early-stage photosynthetic organelle in Paulinella chromatophora. Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Heinrich-Heine-Universität Düsseldorf, Dezember 2020.
- Dana C. Price et al.: Analysis of an improved Cyanophora paradoxa genome assembly. In: DNA Res., Band 26, Nr. 4, August 2019, S. 287–299; doi:10.1093/dnares/dsz009, PMC 6704402 (freier Volltext), PMID 31098614, Epub 14. Mai 2019. Siehe insbes. §3.4.2. Pyrenoid.
- Ludwig Kies: Elektronenmikroskopische Untersuchungen an Paulinella chromatophora Lauterborn, einer Thekamöbe mit blau-grünen Endosymbionten (Cyanellen). In: Protoplasma, Band 80, 1974, S. 69–89; doi:10.1007/bf01666352.
- Protein Databank in Europe: PDBe > 2a1b – Carboxysome shell protein ccmK2
- Cheryl A. Kerfeld: Protein Structures Forming the Shell of Primitive Bacterial Organelles. In: Science. 309, Nr. 5736, 2005, ISSN 0036-8075, S. 936–938. bibcode:2005Sci...309..936K. doi:10.1126/science.1113397. PMID 16081736.
- Cristina V. Iancu, H. Jane Ding, Dylan M. Morris, D. Prabha Dias, Arlene D. Gonzales, Anthony Martino, Grant J. Jensen: The Structure of Isolated Synechococcus Strain WH8102 Carboxysomes as Revealed by Electron Cryotomography. In: Journal of Molecular Biology. 372, Nr. 3, 2007, ISSN 0022-2836, S. 764–773. doi:10.1016/j.jmb.2007.06.059. PMID 17669419. PMC 2453779 (freier Volltext).
- Cristina V. Iancu, Dylan M. Morris, Zhicheng Dou, Sabine Heinhorst, Gordon C. Cannon, Grant J. Jensen: Organization, Structure, and Assembly of α-Carboxysomes Determined by Electron Cryotomography of Intact Cells. In: Journal of Molecular Biology. 396, Nr. 1, 2010, ISSN 0022-2836, S. 105–117. doi:10.1016/j.jmb.2009.11.019. PMID 19925807. PMC 2853366 (freier Volltext).
- Michael F. Schmid, Angel M. Paredesm, Htet A. Khant, Ferda Soyer, Henry C. Aldrich, Wah Chiu, Jessup M. Shively: Structure of Halothiobacillus neapolitanus Carboxysomes by Cryo-electron Tomography. In: Journal of Molecular Biology. 364, Nr. 3, 2006, ISSN 0022-2836, S. 526–535. doi:10.1016/j.jmb.2006.09.024. PMID 17028023. PMC 1839851 (freier Volltext).
- G. Vernizzi, R. Sknepnek, M. Olvera de la Cruz: Platonic and Archimedean geometries in multicomponent elastic membranes. In: Proceedings of the National Academy of Sciences. 108, Nr. 11, 2011, ISSN 0027-8424, S. 4292–4296. doi:10.1073/pnas.1012872108. PMID 21368184. PMC 3060260 (freier Volltext).
- S. Tanaka, Cheryl A. Kerfeld, M. R. Sawaya, F. Cai, S. Heinhorst, G. C. Cannon, T. O. Yeates: Atomic-Level Models of the Bacterial Carboxysome Shell. In: Science. 319, Nr. 5866, 2008, ISSN 0036-8075, S. 1083–1086. bibcode:2008Sci...319.1083T. doi:10.1126/science.1151458. PMID 18292340.
- F. Cai, M. Sutter, J. C. Cameron, D. N. Stanley, J. N. Kinney, Cheryl A. Kerfeld: The Structure of CcmP, a Tandem Bacterial Microcompartment Domain Protein from the β-Carboxysome, Forms a Subcompartment Within a Microcompartment. In: Journal of Biological Chemistry. 288, Nr. 22, Mai 2013, ISSN 0021-9258, S. 16055–16063. doi:10.1074/jbc.M113.456897. PMID 23572529. PMC 3668761 (freier Volltext).
- Michael G. Klein, Peter Zwart, Sarah C. Bagby, Fei Cai, Sallie W. Chisholm, Sabine Heinhorst, Gordon C. Cannon, Cheryl A. Kerfeld: Identification and Structural Analysis of a Novel Carboxysome Shell Protein with Implications for Metabolite Transport. In: Journal of Molecular Biology. 392, Nr. 2, September 2009, ISSN 0022-2836, S. 319–333. doi:10.1016/j.jmb.2009.03.056. PMID 19328811.
- M. Sutter, T. G. Laughlin, K. M. Davies, Cheryl A. Kerfeld: Structure of a synthetic beta-carboxysome shell, T=4. In: dx.doi.org. 25. September 2019. Abgerufen am 19. September 2021.
- J. Zarzycki, Seth D. Axen, James N. Kinney, Cheryl A. Kerfeld: Cyanobacterial-based approaches to improving photosynthesis in plants. In: Journal of Experimental Botany. 64, Nr. 3, 23. Oktober 2012, ISSN 0022-0957, S. 787–798. doi:10.1093/jxb/ers294. PMID 23095996.
- B. D. Rae, B. M. Long, M. R. Badger, G. D. Price: Functions, Compositions, and Evolution of the Two Types of Carboxysomes: Polyhedral Microcompartments That Facilitate CO2 Fixation in Cyanobacteria and Some Proteobacteria. In: Microbiology and Molecular Biology Reviews. 77, Nr. 3, 2013, ISSN 1092-2172, S. 357–379. doi:10.1128/MMBR.00061-12. PMID 24006469. PMC 3811607 (freier Volltext).
- Aiko Turmo, C. Raul Gonzalez-Esquer, Cheryl A. Kerfeld: Carboxysomes: metabolic modules for CO2 fixation. In: FEMS Microbiology Letters. 364, Nr. 18, 14. August 2017, ISSN 1574-6968. doi:10.1093/femsle/fnx176.
- Cheryl A. Kerfeld, Matthew R. Melnicki: Assembly, function and evolution of cyanobacterial carboxysomes. In: Current Opinion in Plant Biology. 31, Juni 2016, ISSN 1369-5266, S. 66–75. doi:10.1016/j.pbi.2016.03.009.
- NCBI: Cyanobium sp. PCC 7001, heterotypic synonyms: Anacystis marina 6, Synechococcus sp. PCC 7001; graphisch: Cyanobium sp. PCC 7001, auf: Lifemap NCBI Version.
- Lynne Whitehead, Benedict M. Long, G. Dean Price, and Murray R. Badger: Comparing the in Vivo Function of α-Carboxysomes and β-Carboxysomes in Two Model Cyanobacteria. In: Plant Physiol., Band 165, Nr. 1, Mai 2014, S. 398–411.; doi:10.1104/pp.114.237941, PMC 4012598 (freier Volltext), PMID 24642960, Epub 18. März 2014.
- J. M. Shively, E. Bock, K. Westphal, G. C. Cannon: Icosahedral inclusions (carboxysomes) of Nitrobacter agilis. In: Journal of Bacteriology. 132, Nr. 2, November 1977, S. 673–675. doi:10.1128/JB.132.2.673-675.1977. PMID 199579. PMC 221910 (freier Volltext).
- G. C. Cannon, J. M. Shively: Characterization of a homogenous preparation of carboxysomes from Thiobacillus neapolitanus. In: Archives of Microbiology. 134, Nr. 1, 1983, ISSN 0302-8933, S. 52–59. doi:10.1007/BF00429407.
- Sabine Heinhorst, Gordon C. Cannon, Jessup M. Shively: Carboxysomes and Their Structural Organization in Prokaryotes. In: Nanomicrobiology. 2014, S. 75–101. doi:10.1007/978-1-4939-1667-2_4. ISBN 978-1-4939-1666-5.
- Fei Cai, Zhicheng Dou, Susan Bernstein, Ryan Leverenz, Eric Williams, Sabine Heinhorst, Jessup Shively, Gordon Cannon, Cheryl A. Kerfeld: Advances in Understanding Carboxysome Assembly in Prochlorococcus and Synechococcus Implicate CsoS2 as a Critical Component. In: Life. 5, Nr. 2, 27. März 2015, ISSN 2075-1729, S. 1141–1171. doi:10.3390/life5021141. PMID 25826651. PMC 4499774 (freier Volltext).
- M. R. Sawaya, G. C. Cannon, S. Heinhorst, S. Tanaka, E. B. Williams, T. O. Yeates, Cheryl A. Kerfeld: The Structure of beta-Carbonic Anhydrase from the Carboxysomal Shell Reveals a Distinct Subclass with One Active Site for the Price of Two. In: Journal of Biological Chemistry. 281, Nr. 11, 2006, ISSN 0021-9258, S. 7546–7555. doi:10.1074/jbc.M510464200. PMID 16407248.
- Balaraj B. Menon, Zhicheng Dou, Sabine Heinhorst, Jessup M. Shively, Gordon C. Cannon: Halothiobacillus neapolitanus Carboxysomes Sequester Heterologous and Chimeric RubisCO Species. In: PLOS ONE. 3, Nr. 10, 2008, ISSN 1932-6203, S. e3570. bibcode:2008PLoSO...3.3570M. doi:10.1371/journal.pone.0003570. PMID 18974784. PMC 2570492 (freier Volltext).
- Manuel Sommer, Fei Cai, Matthew Melnicki, Cheryl A. Kerfeld: β-Carboxysome bioinformatics: identification and evolution of new bacterial microcompartment protein gene classes and core locus constraints. In: J Exp Bot., Band 68, Nr. 14, 22. Juni 2017, S. 3841–3855; doi:10.1093/jxb/erx115., PMC 5853843 (freier Volltext), PMID 28419380, Epub 17. April 2017.
- B. M. Long, M. R. Badger, S. M. Whitney, G. D. Price: Analysis of Carboxysomes from Synechococcus PCC7942 Reveals Multiple Rubisco Complexes with Carboxysomal Proteins CcmM and CcaA. In: Journal of Biological Chemistry. 282, Nr. 40, 2007, ISSN 0021-9258, S. 29323–29335. doi:10.1074/jbc.M703896200. PMID 17675289.
- B. M. Long, L. Tucker, M. R. Badger, G. D. Price: Functional Cyanobacterial beta-Carboxysomes Have an Absolute Requirement for Both Long and Short Forms of the CcmM Protein. In: Plant Physiology. 153, Nr. 1, 2010, ISSN 0032-0889, S. 285–293. doi:10.1104/pp.110.154948. PMID 20304968. PMC 2862411 (freier Volltext).
- J. N. Kinney, A. Salmeen, F. Cai, Cheryl A. Kerfeld: Elucidating Essential Role of Conserved Carboxysomal Protein CcmN Reveals Common Feature of Bacterial Microcompartment Assembly. In: Journal of Biological Chemistry. 287, Nr. 21, 2012, ISSN 0021-9258, S. 17729–17736. doi:10.1074/jbc.M112.355305. PMID 22461622. PMC 3366800 (freier Volltext).
- Gordon C. Cannon, Sabine Heinhorst, Cheryl A. Kerfeld: Carboxysomal carbonic anhydrases: Structure and role in microbial CO2 fixation. In: Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1804, Nr. 2, 2010, ISSN 1570-9639, S. 382–392. doi:10.1016/j.bbapap.2009.09.026. PMID 19818881.
- Manuel Sommer, Markus Sutter, Sayan Gupta, Henning Kirst, Aiko Turmo, Sigal Lechno-Yossef, Rodney L. Burton, Christine Saechao, Nancy B. Sloan, Xiaolin Cheng, Leanne-Jade G. Chan: Heterohexamers Formed by CcmK3 and CcmK4 Increase the Complexity of Beta Carboxysome Shells. In: Plant Physiology. 179, Nr. 1, 2. November 2018, ISSN 0032-0889, S. 156–167. doi:10.1104/pp.18.01190. PMC 6324227 (freier Volltext).
- Jeffrey C. Cameron, Steven C. Wilson, Susan L. Bernstein, Cheryl A. Kerfeld: Biogenesis of a Bacterial Organelle: The Carboxysome Assembly Pathway. In: Cell. 155, Nr. 5, ISSN 0092-8674, S. 1131–1140. doi:10.1016/j.cell.2013.10.044. PMID 24267892.
- Swan S.-W. Cot, Anthony K.-C. So, George S. Espie: A Multiprotein Bicarbonate Dehydration Complex Essential to Carboxysome Function in Cyanobacteria. In: Journal of Bacteriology. 190, Nr. 3, 2007, ISSN 0021-9193, S. 936–945. doi:10.1128/JB.01283-07. PMID 17993516. PMC 2223583 (freier Volltext). Epub 20. Dezember 2020.
- Benedict M. Long, Benjamin D. Rae, Murray R. Badger, G. Dean Price: Over-expression of the β-carboxysomal CcmM protein in Synechococcus PCC7942 reveals a tight co-regulation of carboxysomal carbonic anhydrase (CcaA) and M58 content. In: Photosynthesis Research. 109, Nr. 1–3, 2011, ISSN 0166-8595, S. 33–45. doi:10.1007/s11120-011-9659-8. PMID 21597987.
- C. Raul Gonzalez-Esquer, Tyler B. Shubitowski, Cheryl A. Kerfeld: Streamlined Construction of the Cyanobacterial CO2-Fixing Organelle via Protein Domain Fusions for Use in Plant Synthetic Biology. In: The Plant Cell. 27, Nr. 9, 28. August 2015, ISSN 1040-4651, S. 2637–2644. doi:10.1105/tpc.15.00329. PMC 4815102 (freier Volltext).
- Cheryl A. Kerfeld: Plug‐and‐play for improving primary productivity. In: American Journal of Botany. 102, Nr. 12, Dezember 2015, ISSN 0002-9122, S. 1949–1950. doi:10.3732/ajb.1500409.
- C. Raul Gonzalez‐Esquer, Sarah E. Newnhamn, Cheryl A. Kerfeld: Bacterial microcompartments as metabolic modules for plant synthetic biology. In: The Plant Journalb. 87, Nr. 1, 20. Juni 2016, ISSN 0960-7412, S. 66–75. doi:10.1111/tpj.13166.
- Walter Bonacci, Poh K. Teng, Bruno Afonso, Henrike Niederholtmeyer, Patricia Grob, Pamela A. Silver, David F. Savage: Modularity of a carbon-fixing protein organelle. In: Proceedings of the National Academy of Sciences. 109, Nr. 2, 10. Januar 2012, ISSN 0027-8424, S. 478–483. doi:10.1073/pnas.1108557109. PMID 22184212. PMC 3258634 (freier Volltext).
- Fei Cai, Markus Sutter, Susan L. Bernstein, James N. Kinney, Cheryl A. Kerfeld: Engineering Bacterial Microcompartment Shells: Chimeric Shell Proteins and Chimeric Carboxysome Shells. In: ACS Synthetic Biology. 4, Nr. 4, 2015, ISSN 2161-5063, S. 444–453. doi:10.1021/sb500226j. PMID 25117559.
- GD Price, MR Badger: Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): Functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. In: Journal of Experimental Botany. 59, Nr. 7, 2008, S. 1441–1461. doi:10.1093/jxb/erm112. PMID 17578868.
- GD Price, JJ Pengelly: The cyanobacterial CCM as a source of genes for improving photosynthetic CO2 fixation in crop species. In: Journal of Experimental Botany. 64, Nr. 3, 2013, S. 753–768. doi:10.1093/jxb/ers257. PMID 23028015.
- JM McGrath, SP Long: Can the cyanobacterial carbon-concentrating mechanism increase photosynthesis in crop species? A theoretical analysis.. In: Plant Physiology. 164, Nr. 4, 2014, S. 2247–2261. doi:10.1104/pp.113.232611. PMID 24550242. PMC 3982776 (freier Volltext).
- X Yin, PC Struik: Can increased leaf photosynthesis be converted into higher crop mass production? A simulation study for rice using the crop model GECROS.. In: Journal of Experimental Botany. 68, Nr. 9, 2017, S. 2345–2360. doi:10.1093/jxb/erx085. PMID 28379522. PMC 5447886 (freier Volltext).
- Myat T. Lin, Alessandro Occhialini, P. John Andralojc, Jean Devonshire, Kevin M. Hines, Martin A. J. Parry, Maureen R. Hanson: α-Carboxysomal proteins assemble into highly organized structures in Nicotianachloroplasts. In: The Plant Journal. 79, Nr. 1, 2014, ISSN 0960-7412, S. 1–12. doi:10.1111/tpj.12536. PMID 24810513. PMC 4080790 (freier Volltext).
- Myat T. Lin, Alessandro Occhialini, P. John Andralojc, Martin A. J. Parry, Maureen R. Hanson: A faster Rubisco with potential to increase photosynthesis in crops. In: Nature. 513, Nr. 7519, 2014, ISSN 0028-0836, S. 547–550. bibcode:2014Natur.513..547L. doi:10.1038/nature13776. PMID 25231869. PMC 4176977 (freier Volltext).
- BM Long, WY Hee: Carboxysome encapsulation of the CO2-fixing enzyme Rubisco in tobacco chloroplasts.. In: Nature Communications. 9, Nr. 1, 2018, S. 3570. bibcode:2018NatCo...9.3570L. doi:10.1038/s41467-018-06044-0. PMID 30177711. PMC 6120970 (freier Volltext).
- Benjamin D. Rae, Benedict M. Long, Britta Förster, Nghiem D. Nguyen, Christos N. Velanis, Nicky Atkinson, Wei Yih Hee, Bratati Mukherjee, G. Dean Price, Alistair J. McCormick: Progress and challenges of engineering a biophysical carbon dioxide-concentrating mechanism into higher plants.. In: Journal of Experimental Botany. 68, Nr. 14, Januar 2017, S. 3717#x200B;-3737. doi:10.1093/jxb/erx133. PMID 28444330..
- Matthew R. Melnicki, Markus Sutter, Cheryl A. Kerfeld: Evolutionary relationships among shell proteins of carboxysomes and metabolosomes. In: Current Opinion in Microbiology. 63, Oktober 2021, S. 1–9. doi:10.1016/j.mib.2021.05.011. PMID 34098411. PMC 8525121 (freier Volltext). Siehe insbes. Fig. 1.
- Cheryl A. Kerfeld, Clement Aussignargues, Jan Zarzycki, Fei Cai, Markus Sutter: Bacterial microcompartments. In: Nature Reviews Microbiology, Band 16, Mai 2018, S. 277–290; doi:10.1038/nrmicro.2018.10, Epub 5. März 2018. Siehe insbe. Fig. 2.
{kind=link}
{kind=link}