Calvin-Zyklus
Der Calvin-Zyklus (auch Calvin-Benson-Zyklus[1], Calvin-Benson-Bassham-Zyklus (CBB) oder Ribulosebisphosphatzyklus) ist eine zyklische Folge von chemischen Umsetzungen, durch die Kohlenstoffdioxid (CO2) zu Glucose reduziert und assimiliert wird. Der Stoffwechselweg findet in C3-Pflanzen und mit zusätzlichen Reaktionen in allen anderen Photosynthese betreibenden (photoautotrophen) Lebewesen statt; es handelt sich um deren Dunkelreaktion. Außerdem dient er vielen chemoautotrophen Lebewesen zur Assimilation von Kohlenstoff aus Kohlenstoffdioxid. In Analogie zum Citratzyklus wird der Calvin-Zyklus auch als reduktiver Pentosephosphat-Zyklus bezeichnet. Der Zyklus wurde von den US-amerikanischen Biochemikern Melvin Calvin, Andrew A. Benson und James Alan Bassham in der Zeit von 1946 bis Mitte der 1950er Jahre entdeckt und manchmal nach Benson und Calvin oder nach allen drei Forschern benannt.
Der Calvin-Zyklus besteht aus mehreren zyklisch angeordneten enzymatischen Teilschritten und läuft bei Pflanzen im Stroma der Chloroplasten ab, bei Bakterien dagegen im Cytoplasma. Die einzelnen Teilschritte lassen sich in drei Phasen einteilen: die Fixierung von CO2, die Reduktion des primären Fixierungsproduktes (3-Phosphoglycerat) und die Regeneration des CO2-Akzeptors (Ribulose-1,5-bisphosphat).
Als Reduktionsmittel für die CO2-Reduktion im Calvin-Zyklus dient NADPH, das dabei zu NADP+ oxidiert wird. Die Reduktion ist endergon, als Energiequelle dient ATP, das Energie abgibt, indem es in ADP und Phosphat gespalten wird.
Bei photoautotrophen Lebewesen werden NADPH und ATP durch die sogenannte Lichtreaktion der Photosynthese gebildet und für den Calvin-Zyklus zur Verfügung gestellt. Bei chemoautotrophen Lebewesen werden NADPH und ATP durch die exergonen chemischen Umsetzungen ihres Energiestoffwechsels gebildet.
Die Einzelschritte des Zyklus
CO2-Fixierung
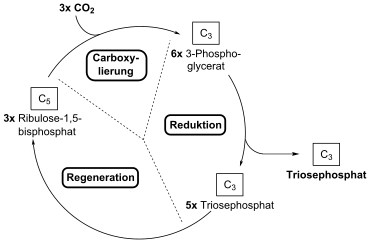
Im ersten Schritt des Calvin-Zyklus wird CO2 durch das Schlüsselenzym RuBisCO an Ribulose-1,5-bisphosphat (RuBP2) als Akzeptormolekül addiert; die hochgradig instabile Zwischenstufe (3-keto-2-carboxyarabinitol 1,5-bisphosphate[2]) zerfällt spontan in zwei Moleküle 3-Phosphoglycerat (3-PG), das erste fassbare Zwischenprodukt bei C3-Pflanzen.
Das primäre Fixierungsprodukt 3-Phosphoglycerat ist nicht nur ein wichtiges Zwischenprodukt im Calvin-Zyklus, sondern tritt auch an anderen wichtigen Stellen beim Auf- bzw. Abbau der Glucose auf: der Gluconeogenese bzw. Glykolyse im Cytoplasma. Es dient auch als Vorläufer zum Aufbau der Stärkespeicher im Chloroplasten. Bevor 3-PG jedoch in die genannten Reaktionen eintritt, wird es im nächsten Teil des Calvinzyklus im Chloroplasten zu Glycerinaldehyd-3-phosphat (abgekürzt G3P oder GAP) reduziert.
Reduktion des primären Fixierungsproduktes (3-Phosphoglycerat)
Nach Phosphorylierung durch eine Kinase und Reduktion durch eine spezielle Glycerinaldehyd-3-phosphat-Dehydrogenase (GAPDH; als Reduktans NADPH statt NADH) entsteht der Gluconeogenesemetabolit Glycerinaldehyd-3-phosphat (GAP), ein wichtiger Verzweigungspunkt. Da in jedem Umlauf ein Molekül CO2 fixiert wird, steht nach jeweils drei Umläufen in der Bilanz ein Molekül der Triose GAP für Biosynthesen zur Verfügung, und steht mit Dihydroxyacetonphosphat (DHAP) im Gleichgewicht. Beide werden auch als Triosephosphate bezeichnet. Sie sind die ersten als Assimilationsgewinn entstehenden Kohlenhydrate und können entweder
- zur Bildung des als Reservestoff dienenden Polysaccharids Stärke im Stroma der Chloroplasten von Pflanzen dienen oder
- über die Zwischenstufe Dihydroxyacetonphosphat (DHAP) und im Gegentausch zu anorganischem Phosphat (Pi) ins Cytoplasma ausgeschleust werden, wo sie
- in die Glycolyse bzw. Gluconeogenese einfließen oder
- der Synthese cytosolischen von dem Transportzucker Saccharose (Rohrzucker; siehe unten) sowie
- der Synthese des Zellwandmaterials Cellulose dienen können.
Mit Saccharose können über das Phloem auch andere Pflanzenteile mit Zucker versorgt werden. Damit der Zyklus wieder beginnen kann, muss allerdings ein Teil der Triosephosphate zum primären Akzeptor Ribulose-1,5-bisphosphat regeneriert werden. Dazu dient der dritte Teil des Calvinzyklus.
Regeneration von Ribulose-1,5-BP
Im dritten Teil erfolgt der Ringschluss des Calvin-Zyklus über den reduktiven Pentosephosphatweg. Bei der Fixierung von drei CO2 an Ribulose-1,5-bisphosphat (C5) entstehen folgerichtig sechs Triosephosphate (C3). Davon ist aber nur eines „echter“ Assimilationsgewinn, aus den anderen fünf müssen wieder die drei verbrauchten Ribulose-1,5-bisphosphate regeneriert werden.
CO2-Fixierung

Im Detail wird die CO2-Gruppe an das C2-Atom der Enolform des Ribulose-1,5-bisphosphat addiert. Es entsteht eine enzymgebundene, hypothetische 3-oxo-Säure (Arabinit; genau: 2-Carboxy-3-keto-D-arabinol-1,5-bisphosphat) als instabile Zwischenstufe, die spontan (durch Wasser am C3-Atom hydrolysiert) in zwei Moleküle der Triose-Vorstufe 3-Phosphoglycerat (3-PG) zerfällt. Dabei entstehen aus dem vorher genannten Arabinit (C6) erst ein Molekül D-3-Phosphoglycerat (C3) und ein aus drei C-Atomen bestehendes Carbanion (ebenfalls C3), das durch Protonierung ebenfalls in das primäre Fixierungsprodukt Phosphoglycerat überführt wird. Dadurch werden netto pro gebundenem Kohlenstoffdioxid zwei Moleküle an Phosphoglycerat erzeugt, von denen eines der beiden den neu hinzufixierten Kohlenstoff des Kohlenstoffdioxids enthält.
Reduktion des primären Fixierungsproduktes (3-Phosphoglycerat)

Die Schritte auf dem Weg von 3-Phosphoglycerat zum Glycerinaldehyd-3-phosphat gleichen denen der Gluconeogenese und werden durch Isoenzyme im Chloroplasten katalysiert. Die Reaktion findet in zwei Teilschritten statt. Zunächst wird das 3-Phosphoglycerat durch Phosphorylierung zu 1,3-Bisphosphoglycerat aktiviert. Dazu wird von der katalysierenden Kinase Energie in Form von ATP verbraucht. Danach kann 1,3-Bisphosphoglycerat unter Abspaltung des eben eingeführten Phosphatrestes zu Glycerinaldehyd-3-phosphat (GAP) reduziert werden. Das katalysierende Enzym ist die Licht-aktivierte Glycerinaldehyd-3-phosphat-Dehydrogenase. Bei diesem Schritt wird NADPH als Reduktionsmittel benötigt. Das cytoplasmatische Enzym der Gluconeogenese arbeitet dagegen mit NADH als Reduktionsmittel.
Regeneration von Ribulose-1,5-BP
Im reduktiven Pentosephosphatweg werden drei Moleküle GAP und zwei Moleküle DHAP in einer verzweigten Reaktionsfolge über verschiedene C3, C4, C6 und C7 -Zucker Zwischenprodukte schließlich in drei C5-Moleküle umgewandelt. Diese werden in Ribulose-5-phosphat umgewandelt und mit ATP zu Ribulose-1,5-bisphosphat phosphoryliert. Für diese Prozesse sind vor allem Aldolasen, Transketolasen nötig und außerdem Phosphatasen.
Reaktionen des reduktiven Pentosephosphatweges (für 3CO2):
- Aldolase: GAP (C3) + DHAP (C3) → Fructose-1,6-BP (C6)
- Fructose-1,6-bisphosphat-Phosphatase: Fructose-1,6-BP + H2O → Fructose-6-P + Pi
- Transketolase: Fructose-6-P (C6) + GAP (C3) → Erythrose-4-P (C4) + Xylulose-5-P (C5)
- Aldolase: Erythrose-4-P (C4) + DHAP (C3) → Sedoheptulose-1,7-BP (C7)
- Sedoheptulose-1,7-bisphosphat-Phosphatase: Sedoheptulose-1,7-BP + H2O → Sedoheptulose-7-P + Pi
- Transketolase: Sedoheptulose-7-P (C7) + GAP (C3) → Xylulose-5-P (C5) + Ribose-5-P (C5)
- Rib5P-Epimerase: 2 Xylulose-5-P (C5) → 2 Ribulose-5-P (C5)
- Rib5P-Isomerase: Ribose-5-P (C5) → Ribulose-5-P (C5)
- Ribulose-5-phosphat-Kinase: 3 Ribulose-5-P (C5) + 3 ATP → 3 Ribulose-1,5-BP (C5) + 3 ADP
Summengleichung des Calvin-Zyklus


Je drei CO2 müssen insgesamt neun ATP und sechs NADPH aufgewendet werden.
Jeweils sechs Moleküle ATP und sechs NADPH werden zur Reduktion eingesetzt (sechs Moleküle Glycerinsäure-3-phosphat werden zu sechs Glycerinaldehyd-3-phosphat reduziert). Dabei entstehen jeweils sechs ADP, sechs Phosphat und sechs NADP+. Die anderen drei ATP werden bei der Regeneration des Akzeptors verbraucht (drei Moleküle Ribulose-5-P werden zu drei Molekülen Ribulose-1,5-BP phosphoryliert), es entstehen drei ADP.
Insgesamt werden neun Moleküle ATP hydrolysiert, wobei neun Moleküle ADP und acht Moleküle Phosphat in der Bilanz freigesetzt werden. Das verbleibende neunte Phosphat findet sich im Glycerinaldehyd-3-phosphat wieder.
Regulation des Calvin-Zyklus
Für die Aktivierung einiger der an den Reaktionen beteiligten Enzyme wird Licht benötigt. Dazu gehört nicht nur das Enzym RuBisCO, welches die Fixierung selbst katalysiert. Sondern auch Enzyme im reduktiven Teil des Calvinzyklus (Glycerinaldehyd-3-phosphat-Dehydrogenase) sowie im Regenerativen Teil (Fructose-1,6-bisphosphat-Phosphatase, Sedoheptulose-1,6-bisphosphat-Phosphatase, Sedoheptulose-1,7-bisphosphat-Phosphatase und Ribulose-5-phosphat-Kinase). In reiner Dunkelheit sind diese Enzyme inaktiv, da die zur Assimilation benötigte Energie und Reduktionsäquivalente fehlen. Die Aktivierung erfolgt über den Mechanismus des Ferredoxin-Thioredoxin-Systems. Dabei wird Thioredoxin durch Ferredoxin aus der Lichtreaktion der Photosynthese von der Disulfid (S-S)- in die Dithiol (SH)-Form überführt. Thioredoxin reduziert dann seinerseits Disulfidbrücken in den verschiedenen Enzymen, welche dadurch aktiviert werden. Im Dunkeln wird die Dithiolform der Enzyme von molekularem Sauerstoff wieder zur Disulfid-Form oxidiert. Dabei entsteht Wasser.
Kohlenhydratbildung bei Pflanzen
Nach jeweils drei Durchläufen des Calvin-Zyklus kann in der Bilanz ein Molekül Glycerinaldehyd-3-phosphat (GAP) aus dem Calvin-Zyklus für weitere Synthesen abgezweigt werden. Ein zentrales Produkt der Assimilation im Chloroplasten von Pflanzen ist Stärke, die sich in Form von Granula (Stärkekörnern) zunächst im Stroma ablagert. Aus diesem Zwischenspeicher werden bei Bedarf Kohlenhydrate in Form von Triosephosphaten freigesetzt, die dann im Cytoplasma zum Disaccharid Saccharose umgesetzt werden. Saccharose ist die wichtigste Transportform von Kohlenhydraten, die durch die Siebröhren des Phloems in Speicherorgane aus nicht photosynthetisch aktiven Zellen (Wurzeln, Knollen, Mark) gelangt. Dort kann der Zucker weiter verwertet oder gespeichert werden. Zur Verwertung gehören z. B. die Glycolyse nicht-photosynthetischer Gewebe (und photosynthetischer Gewebe bei Dunkelheit) sowie die Synthese von Cellulose, Nukleotiden und anderen zuckerhaltigen Zellkomponenten. Bei der Speicherung bilden sich erneut Stärkekörner (Stärke-Granula) in Formen, die für die Pflanze und das Gewebe charakteristisch sind (kugelig, oval, linsen-, spindel- oder stabförmig).
Photosynthesetypen
Wie unter Photorespiration ausgeführt, ist die RubisCO bei normalem CO2-Partialdruck der Luft ineffizient. C4-Pflanzen und CAM-Pflanzen unterdrücken die Nebenreaktion daher durch Vorfixierung von CO2. Ermöglicht wird das durch eine "ATP-getriebene CO2-Pumpe". Katalysiert durch eine chloroplastische Pyruvat-Phosphat-Dikinase entsteht als primärer CO2-Akzeptor aus Pyruvat (Pyr) Phosphoenolpyruvat (PEP). Dabei wird Energie in Form von ATP investiert. Eine cytosolische PEP-Carboxylase katalysiert die Kondensation von Kohlenstoffdioxid in Form von Hydrogencarbonat (HCO3−) an PEP. Das Produkt ist die C4-Verbindung Oxalacetat (OA).
- in C4-Pflanzen wird OA in Form von L-Malat oder L-Aspartat in einen benachbarten Zelltyp, den Bündelscheidenzellen, transportiert. Dort wird es wieder in OA umgewandelt und diese C4-Verbindung decarboxyliert. Das freiwerdende Kohlenstoffdioxid dient dann als Substrat für RuBisCO und wird wie weiter oben dargelegt fixiert.
- in (obligaten) CAM-Pflanzen wird OA durch eine Malatdehydrogenase in L-Malat reduziert und dann in den Vakuolen derselben Zelle unter Energieverbrauch gespeichert. Diese Prozesse finden in der Nacht statt. Am Tag wird das gespeicherte Malat wieder freigesetzt und analog wie bei C4-Pflanzen decarboxyliert. Die Fixierung des Kohlenstoffdioxids entspricht dann den weiter oben beschriebenen Schritten.
Durch die räumliche (C4-Pflanzen) bzw. zeitliche (CAM-Pflanzen) Trennung von Kohlenstoffdioxidvorfixierung und Verbrauch in der RuBisCO-Reaktion entstehen lokal sehr hohe CO2-Partialdrücke, die einer Photorespiration entgegenwirken.
Pyruvat-Phosphat-Dikinase
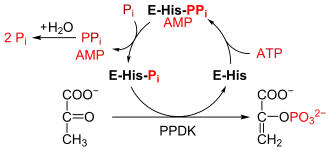
Die Umsetzung von Phosphoenolpyruvat (PEP) in Pyruvat, eine Reaktion der Glycolyse, ist so exergon, dass sie in umgekehrter Richtung nicht direkt (d. h. durch Aufwenden nur eines Moleküls ATP) durchlaufen werden kann. Um Pyruvat zu PEP zu phosphorylieren, verwenden Chloroplasten eine Pyruvat-Phosphat-Dikinase (EC 2.7.9.1). Dieses Enzym hat die ungewöhnliche Eigenschaft, eine Phosphatgruppe durch ATP-Hydrolyse (zu AMP) zu aktivieren. Mechanistisch geschieht dies durch Übertragung eines Pyrophosphatrestes (PPi) auf das Enzym und dessen nachfolgende Phosphorolyse nach in der Abbildung angegebenem Schema.
Literatur
- Hans Walter Heldt, Birgit Piechulla: Pflanzenbiochemie. 4. Auflage. Spektrum Akademischer Verlag, Heidelberg 2008, ISBN 978-3-8274-1961-3.
- Caroline Bowsher, Martin Steer, Alyson Tobin: Plant Biochemistry. Garland Science, New York / Abingdon 2008, ISBN 978-0-8153-4121-5.
Einzelnachweise
- z. B. Thomas D. Sharkey, Discovery of the canonical Calvin–Benson cycle, Photosynthesis Research, Band 140, 2019, S. 235–252
- J. Pierce, T. J. Andrews G. H. Lorimer: Reaction intermediate partitioning by ribulose-bisphosphate carboxylases with differing substrate specificities. Abgerufen am 22. Juni 2017 (englisch).