Blindmaulwurf
Der Blindmaulwurf (Talpa caeca) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae) innerhalb der Ordnung der Insektenfresser (Eulipotyphla). Er kommt im südlichen Europa vor, wo er in zwei räumlich nicht zusammenhängenden (disjunkten) Gebieten auftritt: einerseits im südwestlichen Teil der Alpen und in den Apenninen, andererseits auf der westlichen Balkanhalbinsel. Äußerlich ähnelt der Blindmaulwurf dem Europäischen Maulwurf. Allerdings ist er deutlich kleiner als sein bekannterer Verwandter, zudem bleiben seine Augen als markantes Unterscheidungsmerkmal unter der Haut verborgen. Die Tiere leben vorwiegend in Gebirgsregionen. Dort nutzen sie verschiedene Wälder und Wiesenlandschaften als Habitate, die auf sandigen oder lehmigen Böden wachsen. Sie graben Gänge und Tunnel und ernähren sich zumeist von Regenwürmern. Die Art wurde bereits 1822 wissenschaftlich eingeführt. Teilweise schloss sie im weiteren Verlauf der Forschungsgeschichte auch andere Maulwurfsvertreter des westlichen Eurasiens ein. Ihr Bestand gilt als nicht gefährdet.
| Blindmaulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Blindmaulwurf (Talpa caeca) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Talpa caeca | ||||||||||||
| Savi, 1822 |
Merkmale
Habitus
Der Blindmaulwurf ist ein kleiner Vertreter der Eurasischen Maulwürfe und wird kleiner als der ähnliche Europäische Maulwurf (Talpa europaea) sowie die weiteren zum Teil ebenfalls sympatrisch vorkommenden Arten wie der Römische Maulwurf (Talpa romana) und der Balkan-Maulwurf (Talpa stankovici). Die Kopf-Rumpf-Länge beträgt 10,3 bis 12,4 cm, die Schwanzlänge 2,6 bis 4,3 cm und die Länge des Hinterfußes 1,6 bis 1,8 cm. Die Tiere wiegen 38 bis 67 g. Männliche Individuen sind deutlich größer als weibliche, der Unterschied liegt bei 13 bis 25 %, ist aber abhängig von der jeweiligen Population. Es bestehen außerdem markante geographische Größenabweichungen. Generell ist der Blindmaulwurf auf der Balkanhalbinsel kleiner als in anderen Teilen des Verbreitungsgebietes, außerdem auch da, wo er in direkter Sympatrie mit dem Europäischen und dem Römischen Maulwurf auftritt. Äußerlich ähnelt der Blindmaulwurf den anderen Eurasischen Maulwürfen. Er zeichnet sich durch einen zylindrischen sowie robust gebauten Körper aus, der Kopf sitzt auf einem kurzen Hals und die Vorderfüße sind schaufelartig gestaltet sowie nach außen gedreht. Das Fell weist eine schwarze Farbgebung aus und glänzt seidig. Anders als beim Europäischen Maulwurf überdeckt eine Membran die Augen. Der Nasenspiegel ist klein, rund 3,3 bis 6,6 mm weit und 4,2 bis 8,2 mm breit.[1]
Schädel- und Gebissmerkmale
Die Schädellänge beträgt 27,1 bis 31,7 mm, die Breite am Hirnschädel 13,5 bis 15,3 mm und die Höhe 7,4 bis 9,3 mm. Allgemein ist der Schädel klein und grazil. Das Rostrum zeigt sich länger und schmaler als bei den anderen sympatrischen Maulwurfsarten. Am Eckzahn wird es 3,4 bis 4,1 mm breit, an den Molaren 7,1 bis 9,1 mm. Ersterer Wert entspricht rund 12 bis 14 % der Schädellänge, letzterer rund 26 bis 30 %. Ein weiterer Unterschied betrifft die Lage des Foramen infraorbitale, welches sich beim Blindmaulwurf mit seiner hinteren Kante oberhalb des zweiten und dritten Mahlzahns befindet. Es sind insgesamt 44 Zähne ausgebildet, die Zahnformel lautet: . Der vorderste obere Schneidezahn ist vergleichsweise groß und beansprucht die halbe Länge der Schneidezahnreihe. Der erste Mahlzahn im Oberkiefer besitzt ein zweispitziges Mesostyl, ein kleiner Höcker zwischen den beiden lippenseitigen Haupthöckern (Paraconus und Metaconus). Dies stimmt mit dem Römischen Maulwurf überein, weicht aber vom Europäischen Maulwurf ab. Im Unterschied zum Römischen Maulwurf und entsprechend zum Balkan-Maulwurf ist Oligodontie beim Blindmaulwurf kaum belegt. Die obere Zahnreihe erstreckt sich über 10,3 bis 12,3 mm.[2][1]

Genetische Merkmale
Abweichend von den meisten anderen Vertretern der Eurasischen Maulwürfe in Europa hat der Blindmaulwurf einen diploiden Chromosomensatz von 2n = 36. Dadurch tritt ein zusätzliches Paar an kleinen Chromosomen auf. Darüber hinaus bestehen einzelne Variationen im Karyotyp innerhalb der Populationen. Bei Tieren von der Apenninenhalbinsel und aus den Alpen beträgt die Anzahl der Chromosomenarme 70 beziehungsweise 68 bei den Autosomen (fundamentale Anzahl). Dagegen ist die entsprechende Anzahl der Chromosomen- beziehungsweise Autosomenarme bei Tieren von der Balkanhalbinsel mit 68 und 66 geringer. Das X-Chromosom ist meta- bis submetazentrisch, das Y-Chromosom fleckenförmig.[2][3][1]
Verbreitung und Lebensraum
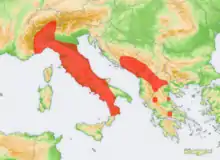
Der Bildmaulwurf kommt in zwei voneinander getrennten Verbreitungsarealen im südlichen und südöstlichen Europa vor. Das eine umfasst den südwestlichen Teil der Alpen mit Teilen der Schweiz und des südöstlichen Frankreichs sowie den Apennin bis zur Südspitze Italiens.[4] Das andere liegt davon isoliert auf der westlichen Balkanhalbinsel und umfasst etwa den Raum zwischen dem Fluss Neretva in Bosnien-Herzegowina südwärts bis Albanien[5][6] und westwärts über Nordmazedonien[7] bis zum Golf von Korinth in Griechenland. Einzelne Vorkommen wurden auch aus der Umgebung von Dubrovnik in Kroatien vermeldet.[8][1]
Die Art bewohnt Laubwälder, Wiesen und Weiden, die Höhenverbreitung reicht vom Meeresspiegelniveau in Dalmatien bis auf rund 2200 m, mitunter auch bis 2500 m. Der bevorzugte Untergrund besteht aus mächtigen sandigen, lehmigen oder sandig-lehmigen Böden, die in manchen Gebieten eine dicke Laubfallschicht von bis zu 20 cm tragen können. Trockene und harte Böden meidet der Blindmaulwurf hingegen. In feuchteren Landschaften ist er in Gebieten mit Schilfrohr und Binsen anzutreffen, waldreichere Habitate sind mit der Silber-Pappel, Weiden, Erlen und der Manna-Esche bewachsen. Gebirgsregionen wie die Massive von Orsomarso und Sila im südlichen italien zeichnen sich durch Kastanien-, Eichen-, Buchen-, Tannen-, Pinien- und Fichten-Pflanzengemeinschaften aus.[4] Teilweise wurden Tiere auch in dichtem Gestrüpp mit Wacholder, Haselnuss und Rubus beobachtet. In den Karstgebieten der Balkanhalbinsel ist das Vorkommen der Art auf Bereiche mit tiefgründigen Böden beschränkt, wie sie mitunter in Dolinen ausgebildet sind.[9] In weiten Bereichen seines Vorkommens lebt der Blindmaulwurf sympatrisch zum Europäischen Maulwurf, zum Römischen Maulwurf und zum Balkan-Maulwurf, allerdings tritt er nur selten an gleichen Fundplätzen auf. Vielmehr okkupiert der Blindmaulwurf bei Anwesenheit anderer Maulwurfarten weitgehend nur die Randgebiete seiner möglichen ökologischen Bandbreite, wie etwa in Italien, wo er die höheren Bergregionen einnimmt und so an kühlklimatische Bedingungen angepasst ist.[10][11] In der Regel ist der Blindmaulwurf seltener als die anderen, größeren Maulwurfarten. Seine Populationen wird aber stark vom Standort bestimmt. In Gebirgslagen sind Nord- und Osthänge häufig dichter und enger besiedelt als Südhänge.[9][1]
Lebensweise
Territorialverhalten
Die Lebensweise des Blindmaulwurfs ist nur wenig erforscht, dürfte aber weitgehend der des Europäischen Maulwurfs ähneln.[12] Die Aktivitätszeiten verteilen sich auf mehrere Phasen am Tag. In den Gebirgen des südlichen Italien wurden verstärkte Aktivitäten zwischen 12:00 und 18:00 Uhr festgestellt, teilweise auch zwischen 06:00 und 12:00 Uhr, während sie in den Stunden vor und nach Mitternacht deutlich zurückgehen. Wahrscheinlich hat die Umgebungstemperatur einen signifikanten Einfluss auf die Aktivitäten des Blindmaulwurfs.[13]
Die Tiere bewegen sich unterirdisch fort und kommen nur selten an die Erdoberfläche. Die von ihnen angelegten Tunnel und Gänge haben einen Durchmesser von 4,0 bis 4,5 cm und verlaufen zumeist in 10 bis 20 cm Tiefe. In lockeren Böden können sie auch 25 bis 35 cm tief liegen. Die Eingänge werden häufig durch Hügel an Auswurfmaterial (Maulwurfshügel) angezeigt. Diese sind kleiner als beim Europäischen Maulwurf: In der Regel häuft der Blindmaulwurf zwischen 1,0 und 3,5 kg an Erde an, durchschnittlich sind es 1,9 kg. Die Hügel sind rund 10 cm hoch bei einem Durchmesser von 15 bis 20 cm. Bezüglich der Sozialstruktur gibt es keine eindeutigen Aussagen. Einige Beobachtungen lassen auf ein einzelgängerisches und territoriales Verhalten schließen. In anderen Fällen wurden bis zu sechs Individuen beiderlei Geschlechts und unterschiedlichen Alters in den Gängen angetroffen. Mitunter wird auch von dem Vorkommen verschiedener Arten in den Gängen berichtet.[9][1]
Ernährung und Fortpflanzung
Die Hauptnahrung des Blindmaulwurfs besteht wie bei anderen Eurasischen Maulwürfen auch aus Regenwürmern. Seine Aktivitäten und sein Vorkommen werden dadurch durch die Häufigkeit der Beutetiere bestimmt.[9] Die Fortpflanzungsphase beginnt möglicherweise im Frühjahr. Ein einzelnes säugendes Weibchen wurde im Mai angetroffen.[1]
Fressfeinde und Parasiten
Zu den wichtigsten Fressfeinden des Blindmaulwurfs gehört die Schleiereule, in deren Gewöllen sich häufig Reste der Tiere finden.[14] Bedeutende Parasiten konnten vor allem mit Fadenwürmern dokumentiert werden. Nachgewiesen sind unter anderem die Gattungen Capillaria und Tricholinstowia. Daneben kommen auch verschiedene Bandwürmer und Saugwürmer vor.[15]
Systematik
Innere Systematik der Eurasischen Maulwürfe nach Demırtaş et al. 2020[16]
|
Der Blindmaulwurf ist eine Art aus der Gattung der Eurasischen Maulwürfe (Talpa). Zu dieser werden rund ein Dutzend weitere Mitglieder gezählt, darunter mit dem Europäischen Maulwurf (Talpa europaea) auch die bekannteste Form. Die Eurasischen Maulwürfe stehen innerhalb der Tribus der Eigentlichen Maulwürfe (Talpini) und der Familie der Maulwürfe (Talpidae). Die Tribus führt die zumeist grabenden Vertreter der Maulwürfe zusammen, während andere Angehörige der Familie demgegenüber nur teilweise unterirdisch leben, sich oberirdisch fortbewegen oder eine semi-aquatische Lebensweise besitzen.[17]
Die wissenschaftliche Erstbeschreibung des Blindmaulwurfs erbrachte Paolo Savi im Jahr 1822. Er trennte damit Tiere von der Apenninenhalbinsel vom Europäischen Maulwurf ab. Als Typuslokalität der Art gilt die Region um Pisa.[18] Ein bedeutendes Unterscheidungsmerkmal des Blindmaulwurfs vom Europäischen Maulwurf stellen unter anderem die bei ersteren mit Haut überdeckten Augen dar, woher auch der Artname rührt (von lateinisch caecus für “blind”). Des Weiteren bestehen Abweichungen in der Anatomie des Kreuzbeins: Beim Blindmaulwurf zeigt dieses einen caecoidalen Aufbau (die Öffnung des Foramens am vierten Kreuzbeinwirbel ist nach hinten gerichtet), beim Europäischen Maulwurf hingegen einen europaeoidalen (die Öffnung des Foramens am vierten Kreuzbeinwirbel ist durch eine Knochenbrücke überdeckt). Später wurden dem Blindmaulwurf alle kleinwüchsigen Maulwürfe des westlichen Eurasiens zugeordnet, die sich vor allem durch ersteres Merkmal auszeichneten, darunter unter anderem der Iberische Maulwurf (Talpa occidentalis), der Kaukasische Maulwurf (Talpa caucasica) und der Levantinische Maulwurf (Talpa levantis). Aufgrund der dadurch entstandenen weiten Verbreitung des Blindmaulwurfs wurde er im Deutschen teilweise unter dem Trivialnamen „Mittelmeermaulwurf“ geführt.[19][20] Während der Kaukasische Maulwurf bereits in den 1920er Jahren als eigenständig anerkannt wurde, lehnte Günter R. Witte eine Abtrennung des Iberischen Maulwurfs im Jahr 1964 ab, um die besonderen Verwandtschaftsverhältnisse zum Blindmaulwurf zu unterstreichen.[20] Jochen Niethammer wiederum stufte im Jahr 1969 den Iberischen Maulwurf als besondere ökologische Variante des Blindmaulwurfs ein. Nur wenige Jahre später gliederten Heinz Felten und Kollegen wiederum den Levantinischen Maulwurf aus biogeographischen Erwägungen aus.[21][22]
Moderne biochemische und genetische Untersuchungen erbrachten dann ab Ende der 1980er Jahre eine komplexere Systematik der westeurasischen Maulwürfe. Bereits 1987 zeigten Analysen an den Allozymen, dass der Iberische Maulwurf sich deutlich vom Blindmaulwurf unterscheidet.[23] Dies wurde dann im Übergang vom 20. zum 21. Jahrhundert durch molekulargenetische Studien bestätigt. Im Ergebnis davon ließ sich innerhalb der Eurasischen Maulwürfe eine westliche Klade um den Europäischen Maulwurf, den Blindmaulwurf und den Levantinischen Maulwurf von einer östlichen Gruppe um den Kaukasischen Maulwurf abtrennen, die sich wenigstens seit dem Übergang vom Miozän zum Pliozän vor rund 6 bis 5 Millionen Jahren eigenständig entwickelt hat. Innerhalb der westlichen Linie besteht eine nähere Verbindung des Blindmaulwurfs zum Römischen Maulwurf (Talpa romana), deren Abspaltung voneinander an das Ende des Pliozäns und den Beginn des Pleistozäns vor gut 2,5 Millionen Jahren datiert. Der Blindmaulwurf selbst zeigt eine stärkere Diversifizierung zwischen der östlichen Population auf der Balkan- und der westlichen auf der Apenninenhalbinsel. Beide trennten sich im Unterpleistozän vor etwa 1,0 Millionen Jahren. Dieser weit zurückreichende Split könnte auch für einen jeweils eigenständigen Artstatus beider Populationen sprechen.[24][25][26][16]
Je nach Autor werden zwischen zwei und sieben Unterarten des Blindmaulwurfs auseinandergehalten. Zu den bekanntesten zählen:[2]
- T. c. augustana Capolongo & Panascì, 1978; südwestliches Alpengebiet, im Aostatal in Italien
- T. c. beaucournui Grulich, 1969; Balkanhalbinsel, vom Pelister in Nordmazedonien
- T. c. caeca Savi, 1822; Nominatform, Apenninenhalbinsel, aus Pisa in der Toskana
- T. c. hercegovinensis (Bolkay, 1925); Balkanhalbinsel, um Stolac in Bosnien-Herzegowina
- T. c. olympica (Chaworth-Musters, 1932); Balkanhalbinsel, vom Olymp in Griechenland
- T. c. steini Grulich, 1971; Balkanhalbinsel, aus dem Lovćen in Montenegro
Boris Kryštufek hingegen betrachtet in dem im Jahr 2018 herausgegebenen achten Band des Standardwerkes Handbook of the Mammals of the World, das sich mit insektenfressenden Säugetieren beschäftigt, den Blindmaulwurf als monotypisch und unterscheidet keine Unterarten. Seiner Meinung nach wurden die aufgestellten Formen bisher nicht hinreichend voneinander abgegrenzt.[1] Genetischen Untersuchungen zufolge bestehen aber teils erhebliche Variationen zwischen einzelnen Populationen in der Toskana und dem Aostatal. Auch mikroregional konnten einzelne Unterschiede herausgearbeitet werden, die zum Teil auf Migrationshindernisse wie Flüsse zurückzuführen sind.[23][27]
Teilweise wurden Fossilfunde kleinwüchsiger Maulwürfe beschrieben, die sich über weite Bereiche Europas verteilen und deren Alter bis in den Ausgang des Pliozäns zurückreicht. Beispiele hierfür sind Podelsice in Polen, Rivoli Veronese beziehungsweise Soave im nördlichen, Isernia la Pineta im zentralen oder Pirro im südlichen Italien,[28] Hundsheim in Österreich und Petersbuch in Deutschland.[29] Die Reste gelten zumeist als der ausgestorbenen Art Talpa minor zugehörig, welche anhand einzelner Fossilien von Hundsheim ihren Namen erhielt.[30] Andere Bezeichnungen wie Talpa gracilis, geprägt an Funden aus Püspökfürdö in Ungarn und wiederum Hundsheim,[31][32] oder Talpa csarnotana, definiert für Reste aus Csarnóta, ebenfalls Ungarn,[33] stellen wahrscheinlich Synonyme dar. Nach Meinung einiger Autoren wie Björn Kurtén kommt Talpa minor aufgrund seiner geringen Größe als Vorgängerform des Blindmaulwurfs in Betracht. Andere Wissenschaftler wiederum sehen unter Berufung auf eine abweichende Gebissausprägung keine direkten Beziehungen zwischen Talpa minor und dem Blindmaulwurf.[34]
Bedrohung und Schutz
Der Blindmaulwurf kann lokal häufig sein, Hinweise auf Bestandsrückgänge gibt es nicht, ebenso wenig sind größere bestandsgefährdende Bedrohungen bekannt. Aufgrund dessen und mit Blick auf die weite Verbreitung stuft die IUCN die Art als “nicht gefährdet” („least concern“) ein. Sie ist in mehreren Naturschutzgebieten präsent.[35]
Literatur
- Stéphane Aulagnier, Patrick Haffner, Anthony J. Mitchell-Jones, François Moutou und Jan Zima: Die Säugetiere Europas, Nordafrikas und Vorderasiens. Der Bestimmungsführer. Haupt, 2009, ISBN 978-3-258-07506-8, S. 64–65
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 611–612) ISBN 978-84-16728-08-4
- Anthony J. Mitchell-Jones, Giovanni Amori, Wieslaw Bogdanowicz, Boris Krystufek, P. J. H. Reijnders, Friederike Spitzenberger, Michael Stubbe, Johan B. M. Thissen, Vladimiŕ Vohralik und Jan Zima: The Atlas of European Mammals. Poyser, London, 1999, ISBN 0-85661-130-1, S. 80–81
Einzelnachweise
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 611–612) ISBN 978-84-16728-08-4
- Boris Kryštufek: The taxonomy of blind moles (Talpa caeca and T. stankovici, Insectívora, Mammalia) from south-eastern Europe. Bonner Zoologische Beiträge 45 (1), 1994, S. 1–16
- E. Gornung, M. Volleth, E. Capanna und R. Castiglia: Comparative cytogenetics of moles (Eulipotyphla, Talpidae): chromosomal differences in Talpa romana and T. europaea. Cytogenetic Genome Research 121, 2008, S. 249–254, doi:10.1159/000138892
- G. Aloise und Mara Cagnin: New southern distribution limit of Talpa caeca Savi, 1922 (Insectivora, Talpidae) in Italy. Mammalian Biology 68, 2003, S. 235–238
- Ferdinand Bego, Boris Kryštufek, Gligor Paspali und Elton Rogozi: Small terrestrial mammals of Albania: Annotated list and distribution. Hystrix Italian Journal of Mammalogy 19 (2), 2008, S. 3–21
- Ferdinand Bego, Enerit Saçdanaku, Michela Pacifici und Carlo Rondinini: Small terrestrial mammals of Albania: distribution and diversity (Mammalia, Eulipotyphla, Rodentia). ZooKeys 742, 2018, S. 127–163
- Boris Kryštufek und Svetozar Petkovski: Annotated checklist of the mammals of the Republic of Macedonia. Bonner zoologische Beiträge 51 (4), 2002, S. 229–254
- Nikola Tvrtković: First documented record of the blind mole, Talpa caeca, in Croatia. Folia Zoologica 48 (2), 1999, S. 157–160
- Ivo Grulich: Die Standortanscprüche von Talpa caeca Savi (Talpidae, Insectivora). Zoologické Listy 19, 1970, S. 199–219 ()
- Anna Loy, Paolo Colangelo, Flavia Annesi und Ernesto Capanna: Origin and evolution of Western European moles (genus Talpa, Insectivora): a multidisciplinary approach. Mammal Study 30, 2005, S. S13–S17
- Anna Loy, Marcelo H. Cassini, Paolo Colangelo und Mirko Febbraro: Distribution, spatial interaction and niche analysis in three species of European moles (genus Talpa, Soricomorpha: Mammalia) in Italy. Biological Journal of the Linnean Society 122 (4), 2017, S. 872–882
- Stéphane Aulagnier, Patrick Haffner, Anthony J. Mitchell-Jones, François Moutou und Jan Zima: Die Säugetiere Europas, Nordafrikas und Vorderasiens. Haupt, 2009, S. 64
- Mirko di Febbraro und anna Loy: A new method based on indirect evidences to infer activity patterns in moles. A test on the blind mole in the Central Apennines. Folia Zoologica 63 (2), 2014, S. 116–121, doi:10.25225/fozo.v63.i2.a9.2014
- Longino Contoli: Sistemi trofici e corologia: Dati su Soricidae, Talpidae ed Arvicolidae d’Italia predati da Tyto alba (Scopoli 1769). Hystrix 1 (2), 1986, S. 97–118
- A. Ribas und J. C. Casanova: Helminth fauna of Talpa spp. in the Palaeoarctic realm. Journal of Helminthologie 80, 2006, S. 1–6, doi:10.1079/JOH2005328
- Sadık Demırtaş, Metin Silsüpür, Jeremy B. Searle, David Bilton und İslam Gündüz: What should we call the Levant mole? Unravelling the systematics and demography of Talpa levantis Thomas, 1906 sensu lato (Mammalia: Talpidae). Mammalian Biology 100, 2020, S. 1–18, doi:10.1007/s42991-020-00010-4
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Paolo Savi: Lettera del Dott. Paolo Savi al Sig. Dott. Carlo Passerini, conservatore dell’I. e R. Museo di Fisica, e Storia Naturale di Firenze. Nuovo Giornale de' Letterati di Pisa 1, 1822, S. 264–265 ()
- Georg H. W. Stein: Schädelallometrien und Systematik bei altweltlichen Maulwürfen (Talpinae). Mitteilungen aus dem Museum für Naturkunde in Berlin 36 (1), 1960, S. 1–48
- Günter R. Witte: Zur Systematik der Insektenfresser des Monte-Gargano-Gebietes (Italien). Bonner Zoologische Beiträge 15 (1/2), 1964, S. 1–35
- Jochen Niethammer: Zur Taxonomie europäischer Zwergmaulwürfe (Talpa “mizura”). Bonner Zoologische Beiträge 20 (4), 1969, S. 360–372
- Heinz Felten, Friederike Spitzenberger und Gerhard Storch: Zur Kleinsäugerfauna West-Anatoliens. Teil II. Senckenbergiana biologica 54 (4/6), 1973, S. 227–290 (S. 230–232)
- Maria Grazia Filippucci, Giuseppe Nascetti, Ernesto Capanna und Luciano Bullini: Allozyme variation and systematics of European moles of the genus Talpa (Mammalia, Insectivora). Journal of Mammalogy 68 (3), 1987, S. 487–499
- P. Colangelo, A. A. Bannikova, B. Kryštufek, V. S. Lebedev, F. Annesi, E. Capanna und A. Loy: Molecular systematics and evolutionary biogeography of the genus Talpa (Soricomorpha: Talpidae). Molecular Phylogenetics and Evolution 55, 2010, S. 372–380
- Jean-Pierre Hugot, Se Hun Gu, Carlos Feliu, Jacint Ventura, Alexis Ribas, Jérôme Dormion, Richard Yanagihara und Violaine Nicolas: Genetic variability of Talpa europaea and Nova hantavirus (NVAV) in France. Bulletin de l’Académie Vétérinaire de France 167 (3), 2014, S. 177–184
- Anna A. Bannikova, Elena D. Zemlemerova, Paolo Colangelo, Mustafa Sözen, M. Sevindik, Artem A. Kidov, Ruslan I. Dzuev, Boris Kryštufek und Vladimir S. Lebedev: An underground burst of diversity – a new look at the phylogeny and taxonomy of the genus Talpa Linnaeus, 1758 (Mammalia: Talpidae) as revealed by nuclear and mitochondrial genes. Zoological Journal of the Linnean Society 175, 2015, S. 930–948
- F. Suchentrunk, R. Maurizio, Anita Haiden und G. B. Hartl: Allozymic variation and diffentiation in moles (Genus Talpa, Insectivora) of the Val Bragaglia (Switzerland) and the Val Chiavenna (Italy). Zeitschrift für Säugetierkunde 60, 1995, S. 321–329
- Tassos Kotsakis, Laura Abbazzi, Chiara Angelone, Patrizia Argenti, Giancarlo Barisone, Flaviano Fanfani, Federica Marcolini und Ferderico Masini: Plio-Pleistocene biogeography of Italian mainland micromammals. Deinsea 10, 2003, S. 313–342
- Wighart von Koenigswald: Mittelpleistozäne Kleinsäugerfauna aus der Spaltenfüllung Petersbuch bei Eichstätt. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 10, 1970, S. 407–432
- Wilhelm Freudenberg: Die Säugetiere des älteren Quartärs von Mitteleuropa mit besonderer Berücksichtigung der Fauna von Hundsheim in Niederösterreich nebst Ausführungen über verwandte Formen aus Mauer, Mosbach, Cromer und anderen Fundorten. Geologische und Paläontologische Abhandlungen N. F. 12, 1914, S. 455–670 (S. 661) ()
- Theodor Kormos: Diagnosen neuer Säugetiere aus der oberpliozänen Fauna des Somlyóberges bei Püspökfürdö. Annales Musei Nationalis Hungarici 27, 1930, S. 237–246
- Theodor Kormos: Revision der Kleinsäuger von Hundsheim. Földtani közlöny 67, 1937, S. 157–171 ()
- Miklós Kretzoi: Insectivoren, Nagetiere und Lagomorphen der jüngstpliozänen Fauna von Csarnóta im Villányer Gebirge (Südungarn). Vertebrata Hungarica 1 (2), 1959, S. 237–246 ()
- J. T. van Cleef-Roders und L. W. van den Hoek Ostende: Dental morphology of Talpa europaea and Talpa occidentalis (Mammalia: Insectivora) with a discussion of fossil Talpa in the Pleistocene of Europe. Zoologische Mededelingen Leiden 75, 2001, S. 52–67
- F. Cassola: Talpa caeca. The IUCN Red List of Threatened Species 2016. e.T41479A2953438 (); zuletzt aufgerufen am 8. September 2020
Weblinks
- Talpa caeca in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: F. Cassola, 2016. Abgerufen am 8. September 2020.