Brückenechse
Die Brückenechse oder Tuatara (Sphenodon punctatus) ist die einzige rezente Art der Familie Sphenodontidae. Diese nur auf neuseeländischen Inseln verbreiteten Tiere gelten als lebende Fossilien, weil sie Überlebende einer relativ diversen Gruppe sind, der Sphenodontia, deren Blütezeit mehr als 150 Millionen Jahre zurückliegt. Die Tiere leben heute nur noch auf einigen kleineren Inseln von Neuseeland.[1]
| Brückenechse | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Brückenechse (Sphenodon punctatus) | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||
| Sphenodon | ||||||||||
| Gray, 1831 | ||||||||||
| Wissenschaftlicher Name der Art | ||||||||||
| Sphenodon punctatus | ||||||||||
| (Gray, 1842) |
Grundlegende morphologische Eigenheiten gegenüber anderen rezenten Vertretern der Reptilien, speziell das Vorhandensein eines unteren Schläfenbogens, welcher als „Brücke“ eine namensgebende Bedeutung hat und sie von den im Habitus recht ähnlichen Schuppenkriechtieren (Squamata) unterscheidet, rechtfertigen nach derzeitiger Ansicht die Einordnung in eine eigene Ordnung (ebenjene Sphenodontia, auch als Rhynchocephalia bezeichnet). Im Unterschied zu vielen anderen wechselwarmen Reptilien sind sie selbst bei niedrigen Temperaturen aktiv und trotz der deutlich geringeren Körperwärme in der Lage, aktiv nach Beutetieren wie Gliederfüßern oder auch Vogeleiern zu suchen. Über die Lebensweise der bedrohten Brückenechsen ist im Gegensatz zu ihrer Morphologie relativ wenig bekannt.
Etymologie
Der Gattungsname Sphenodon ist zusammengesetzt aus den altgriechischen Wörtern σφήν sphén, deutsch ‚Keil‘ und ὀδούς odous, deutsch ‚Zahn‘. Das Spezies-Epitheton punctatus ist lateinisch und heißt ‚gepunktet‘. Die Trivialbezeichnung „Tuatara“ kommt aus der Sprache der Maori, bedeutet ‚stacheliger Rücken‘[2] und bezieht sich auf den stacheligen Rückenkamm der Tiere. Der deutsche Trivialname „Brückenechse“ wurde 1868 von Eduard von Martens eingeführt. Er weist auf die beiden vollständigen „Knochenbrücken“ (Schläfenbögen) im gefensterten hinteren Teil des Schädels dieser Tiere hin[3] (siehe → Skelett).
Morphologie

Brückenechsen werden durchschnittlich 50 bis 75 Zentimeter lang und wiegen etwa ein Kilogramm, Männchen sind dabei etwas größer. Sie sind kräftig gebaute, plumpe Echsen mit einem Dorsalkamm aus verlängerten Hornplättchen. Der Vorderschädel ist leicht schnabelartig verlängert. Sie haben eine gräuliche Grundfarbe.
Haut
Die Haut der Brückenechsen ist der der Schuppenkriechtiere ähnlich. Die Unterhaut besitzt meist horizontal verlaufende Bindegewebsfasern und liegt locker der unter ihr liegenden Muskulatur auf. Die Lederhaut ist sehr dick und besitzt größere Bindegewebsfaserbündel. Unter der Lederhaut liegt der Großteil der großen, verzweigten Chromatophoren. Ihr körniger, schwarzer bis brauner Pigmentinhalt kann durch Kontraktion und Ausdehnung einen physischen Farbwechsel herbeiführen. Wenn dort auch kleiner, lassen sich die Chromatophoren bis in das Stratum corneum verfolgen. Dorsal bildet die Epidermis etliche Granularschuppen, welche an den Zehen ihre maximale Größe erreichen. Den Körperseiten entlang angeordnete Hautfalten tragen zugespitzte Tuberkelschuppen. Die Beschuppung der Bauchseite besteht aus annähernd quadratischen Schildchen. Vom Hinterhaupt verläuft über die Rückenseite des Körpers ein Kamm, der aus Stachelschuppen besteht und bei Männchen höher ist. Brückenechsen häuten sich nur ein- oder zweimal im Jahr.
Skelett
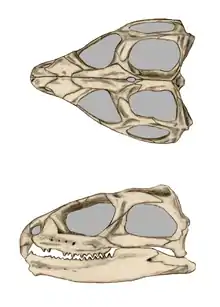
Der diapside Schädel von Brückenechsen unterscheidet sich durch das Vorhandensein zweier voll ausgebildeter Schläfenbögen grundlegend von dem Schädel der übrigen Schuppenechsen. Er verfügt somit auch nicht über das „Schubstangensystem“, das den relativ stark in sich beweglichen (kinetischen) Schädel der übrigen Schuppenechsen auszeichnet. Unter anderem deshalb werden die Brückenechsen in der Systematik auch in eine eigene Untergruppe der Schuppenechsen gestellt, die Sphenodontia. Tatsächlich ist der Schädel der Brückenechsen akinetisch, das heißt, dass Oberkiefer und Gaumendach nicht gegen den Hirnschädel beweglich sind. Allerdings finden sich bei Embryos noch Gelenkungen zwischen den Schädelkomponenten, daher wird der starre Schädel adulter Brückenechsen oft als Anpassung an die grabende Lebensweise und Ernährung gedeutet. Im Zusammenhang mit der Lebensweise stehen auch zwei knöcherne, abwärts gerichtete Fortsätze am vorderen Ende des Prämaxillare (Teil des Oberkiefers), an denen funktionelle „Schneidezähne“ sitzen, was der Schnauzenpartie des Schädels in der Seitenansicht das typische hakenschnabelartige Aussehen verleiht. Die Bezahnung ist akrodont, das heißt, dass die Zähne mit dem oberen Kieferrand verschmolzen sind. Brückenechsen fehlt also jeglicher Zahnwechsel, wodurch aus dem Zustand der Zähne Rückschlüsse auf das Alter gezogen werden können. Bei alten Brückenechsen ist nur noch eine Kauleiste vorhanden. An den äußeren Rändern der Gaumenbeine befindet sich eine Zahnreihe mit elf oder zwölf Zähnen parallel zu den hinteren Zähnen der Maxillaria. Ältere Männchen können zusätzlich ein bis zwei Zähne am Pflugscharbein aufweisen. Am Vorderende des Dentale (zahntragender Teil des Unterkiefers) existiert eine Art Eckzahn, welcher sich stark von der dort vorhandenen spitz-sägeförmigen Zahnreihe unterscheidet. Die Zahnreihe des Unterkiefers greift beim Zubeißen genau in die Lücke zwischen Maxillar- und Gaumenzahnreihe.[3] Da Brückenechsen ihren Unterkiefer nach vorne und zurück schieben können und somit die Beute zerschneiden ("zersägen"), statt sie zu zerreißen, ist es ihnen möglich, harte Chitinpanzer großer Insekten zu knacken und kleine Echsen zu zertrennen.[4]
Anders als bei den Schuppenkriechtieren fehlt bei Brückenechsen mit dem Spleniale das siebte Element des Unterkiefers. Besonders auffällig am Schädel von Brückenechsen ist ein großes Scheitelloch (Foramen parietale) für das Scheitelauge. Das dreieckige, als dünne Knochenplatte ausgebildete Quadratum steht senkrecht zur Schädelmedianebene und wird dorsal durch Flügelbein und Squamosum verstärkt. Ventral bildet das Quadratum zusammen mit dem Quadratojugale eine Gelenkfläche für das gelenkende Element (Articulare) des Unterkiefers.
Das postcraniale Skelett (Skelett ohne Schädel) der Brückenechsen ist durch eine Wirbelsäule mit 27 präcaudalen (vor dem Schwanz liegenden) Wirbeln aus 8 Hals-, 17 Rumpf- und zwei Kreuzwirbeln charakterisiert. Kennzeichnend für den ersten Halswirbel (Atlas) ist ein Rudiment des Proatlas. Wie bei einigen anderen Schuppenkriechtieren kann der Schwanz zum Selbstschutz abgeworfen werden (Autotomie), wobei das Regenerat von einem transparent erscheinenden Knorpelstab gestützt wird. Ab dem vierten Halswirbel haben Brückenechsen kurze Halsrippen. Die Rippen des Rumpfes weisen bei Brückenechsen knorpelartige, flügelähnliche Verbreiterungen auf. Diese überlappen sich und bilden somit eine Art Panzerung für die Leibeshöhle. Die neunte bis zwölfte Rumpfrippe ist unten am Brustbein befestigt.
Nervensystem
Das Gehirn der Brückenechsen ist wesentlich kleiner als das es umgebende Hirnschädelvolumen und deutlich primitiver als das Gehirn anderer Schuppenkriechtiere. Das Endhirn setzt sich in sehr lange Riechbahnen fort, die in dicken Riechkolben enden. Das Mittelhirndach (Tectum mesencephali) liegt sehr viel niedriger als bei Schuppenkriechtieren. Fünfter, siebter, achter, neunter und zehnter Hirnnerv ähneln in ihrer Lage stark denen von Amphibien und Fischen.
Ein besonderes Sinnesorgan, das Scheitelauge, teilen sie nur mit wenigen anderen Reptilien sowie Neunaugen. Es besitzt eine linsenähnliche Epithelschicht, welcher sich ein basaler, netzhautartiger Teil anschließt. Ein dünner Nervus parietalis leitet die Sinnesreize zum Zwischenhirn. Mit diesem Sinnesorgan können sie nur Helligkeit wahrnehmen, das aber wahrscheinlich feiner als „normale“ Augen. Außerdem könnte das Scheitelauge zur Regelung des Wärmehaushaltes dienen.
Die großen Augen von Brückenechsen sitzen in einer dementsprechenden Augenhöhle. Anders als bei Schuppenkriechtieren hat der Musculus retractor bulbi zwei Ansätze am Augapfel. Es fehlt eine Tränendrüse zur Befeuchtung der äußeren Augen, allerdings haben Brückenechsen hinter dem Augapfel eine funktionale Harder-Drüse, welche ein öliges Sekret produziert und so die Nickhaut gleitfähig hält. Überschüssige Feuchtigkeit wird mittels Tränenkanälchen und Tränen-Nasen-Gang abgeleitet, die in die Nasenhöhle führen. Es sind beide Augenlider vorhanden, aber das untere ist deutlich größer und stärker entwickelt. Die transparente Nickhaut ist mit einer Nickhautsehne mit dem sogenannten Musculus bursalis verbunden, welcher die Nickhaut bewegt. Die meist wegen Lichteinfalls schlitzförmige Pupille ist kein Indiz für reine Nachtaktivität, sondern für die hauptsächlich nächtliche Jagdaktivität. Die Netzhaut enthält nicht Stäbchen und Zapfen, sondern zwei verschiedene Zapfentypen, wodurch die Tiere zwar tags und nachts sehr gut sehen können, aber das Farbsehen nicht oder nur rudimentär vorhanden ist. Hinter der Netzhaut findet sich eine Tapetum lucidum genannte Zellschicht, welche durch die Reflexion des durch die Retina dringenden Lichtes als Restlichtverstärker funktioniert.
Die Nase von Brückenechsen weist hinter dem Nasenvorhof (Vestibulum nasi) noch eine zweite größere Erweiterung auf.
Die Geschmacksknospen der Brückenechsen liegen vor allem auf der Gaumenschleimhaut. Die nicht gespaltene Zunge wird, anders als bei etlichen anderen Reptilien, nicht zum Züngeln und somit zur chemosensorischen Orientierung benutzt (siehe auch Jacobson-Organ).
Obwohl Brückenechsen keine äußere Ohröffnung und kein oberflächliches Trommelfell haben, hören sie gut. Das geräumige Mittelohr steht über die Eustachi-Röhre mit der Rachenhöhle in Verbindung. Der restliche Aufbau ist im Wesentlichen mit dem der Schuppenkriechtiere identisch.
Kreislaufsystem und Stoffwechselsystem
Das aus Sinus venosus, einer Herzkammer (Ventriculus cordis) und zwei Herzvorhöfen (Atria) bestehende Herz von Brückenechsen ist durch seine weit vorgeschobene Lage auf Höhe des Schultergürtels charakterisiert. In den Sinus venosus münden drei Hohlvenen ein. Die Vorhöfe wirken von oben gesehen als einheitlicher Sack, sie sind aber innen durch ein gut ausgebildetes Vorhofseptum vollständig getrennt. Die von den Vorhöfen durch eine deutliche Herzkranzfurche (Sulcus coronarius) abgesetzte Herzkammer ist dickwandig und muskulös; im Inneren weist sie kaum Septen auf, eine Plesiomorphie. Weitere Besonderheiten sind ein gut entwickelter Ductus caroticus und Ductus arteriosus auf jeder Seite sowie der Ursprung der rechten und linken Halsschlagader aus einem rechten Aortenbogen. Auch der Ursprung der Arteria laryngea aus dem Pulmonarbogen ist wie die vorher genannten besonderen Gefäßsysteme eine Plesiomorphie.
Der Atmungsapparat der Brückenechsen ist ursprünglich. Der Kehlkopf wird von einem unpaaren Ringknorpel und einem paarigen Stellknorpel gebildet, die Luftröhre wird von knorpeligen Spangen gestützt. Die Luftröhre führt über einen kurzen Bronchus beiderseits in die beiden Lungen, die lediglich dünnwandige Säcke darstellen. Die innere Oberfläche der beiden Lungen ist von einem Netz bienenwabenartiger Räume bedeckt, welche nach hinten größer werden. Mit Hilfe der Kehlkopfknorpel können die sonst stummen Tiere beim Ausatmen zur Verteidigung gegen Feinde und zur Verständigung mit Artgenossen Laute erzeugen.
Der einfach gebaute Verdauungsapparat der Brückenechsen besteht unter anderem aus einer weit dehnbaren Speiseröhre, welche in den langen, spindelförmigen Magen führt. Der Dünndarm liegt in zwei bis drei Schlingen im rechten Teil der Bauchhöhle. Der Dickdarm mündet in den Kotraum (Coprodaeum) der Kloake.
Die linke Niere ist bei Brückenechsen fast doppelt so groß wie die rechte. Je ein kurzer Harnleiter mündet in den Harnraum (Urodaeum) der Kloake. Diese Mündungen sind bei Männchen gemeinsam mit den Samenleitern, aber bei Weibchen getrennt vom Eileiter.
Geschlechtsorgane
Speziell die inneren Geschlechtsorgane weisen keine besondere Spezialisierung auf. Zwar ist bei Weibchen die Fähigkeit zur Spermaspeicherung vorhanden, aber kein spezielles Organ hierfür zu finden. Den Männchen fehlt ein Kopulationsapparat, ein unpaarer Penis ist sekundär verloren gegangen, paarige Hemipenes wie bei Schuppenkriechtieren sind auch nicht in Embryonalstadien vorhanden. Bei beiden Geschlechtern befinden sich manchmal als Hemipenis-Homologa gedeutete Analdrüsen, welche wahrscheinlich Talg produzieren.
Lebensweise

Allgemeines
Brückenechsen sind hauptsächlich dämmerungs- oder nachtaktiv, worauf nicht zuletzt ihre großen Augen mit schlitzartigen Pupillen schließen lassen. Sie graben sich oft eigene Wohnhöhlen in humusartigen Boden, wo sie den Großteil des Tages verbringen. Im Gegensatz zu einer verbreiteten Ansicht leben sie nur selten in Wohnhöhlen von Seevögeln, mehr dazu siehe unten.
Tuataras bewegen sich recht langsam fort. Sie laufen mit abstehenden Beinen und lateralen Wellenbewegungen von Schwanz und Rumpf, welche am Boden schleifen. Kurzfristig können sie mit erhobenen Rumpf spurten, halten diesen Laufstil aber meist nur wenige Meter durch.
Im Unterschied zu fast allen anderen Reptilien, die Körpertemperaturen zwischen 25 und 40 °C bevorzugen, leben Brückenechsen unter wesentlich kühleren Verhältnissen. Die Angaben der Vorzugskörpertemperatur weichen in der Literatur deutlich voneinander ab: Robert Mertens konkretisierte die Vorzugstemperatur auf 10,6 °C[5], in „Grzimeks Tierleben“ wird von 12 °C gesprochen[6], andere Quellen sprechen von 17 bis 20 °C.[7] Offenbar werden sie erst bei 7 °C lethargisch.[8] Der niederen Temperatur entspricht ein langsamer Stoffwechsel: Brückenechsen wachsen sehr langsam, sie können ein hohes Alter erreichen (siehe unten).
Ernährung
Brückenechsen ernähren sich primär von Wirbellosen, meist Insekten (hier besonders Käfern und Heuschreckenartigen), Spinnen und Schnecken sowie Regenwürmern. Nach Aussagen von Einheimischen ist die Weta-Art Deinacrida rugosa die bevorzugte Beute der Brückenechsen.[9] Diese verfolgen sie nicht aktiv, sondern warten oft am Eingang zu ihrer Wohnhöhle, bis eine Weta vorbeikommt. Eine seltene Nahrung von Brückenechsen sind Seevögel – mehr dazu siehe unten.

Fortpflanzung

Die Paarung erfolgt während des südlichen Sommers von Januar bis März, wobei Männchen anders als Weibchen jedes Jahr paarungsbereit sind. Die 25 Quadratmeter großen Territorien der Männchen mit einer Wohnhöhle im Zentrum werden nur zur Paarungszeit verteidigt. Dringt ein Männchen in das Revier eines anderen ein, wird es angegriffen. Dabei kann es durchaus zu Verletzungen kommen, wie zahlreiche Narben an älteren Männchen belegen.
Die eigentliche Umwerbung des Weibchens beginnt, wenn eines in das Territorium des Männchens eindringt. Dabei wird das Weibchen vom Männchen steifbeinig umzirkelt. Da männliche Brückenechsen kein verlängertes Genitalorgan haben, pressen sie ihre Kloake an die des Weibchens. Die Kopulation dauert etwa eine Stunde. Nach ungefähr neun Monaten legen die Brückenechsen-Weibchen in eine selbstgegrabene Erdhöhle ein Gelege von bis zu 15 an den Enden stumpfen, pergamentschaligen, drei Zentimeter langen und vier bis sechs Gramm schweren Eiern. Die Bruthöhle befindet sich oft hunderte Meter weit von der Wohnhöhle des Weibchens entfernt; bei der Bruthöhle verbringen sie anschließend Tage oder auch Wochen. Nach dem Auspolstern des Nestes mit Gras und Erde und dem Schließen der Höhle mit Erde zeigen die Weibchen, anders als die vieler anderer Reptilien, eine Art Brutpflege: Sie halten regelmäßig, manchmal jede Nacht, Wache an ihrem Nest, um zu verhindern, dass andere Weibchen ihre Eier in das Nest legen. In Neuseeland schlüpfen die circa zehn Zentimeter langen und fünf Gramm schweren Jungtiere 13 bis 15 Monate nach der Eiablage; diese längste von Kriechtieren bekannte Brutperiode zeigt, dass die Keimlinge im kälteren neuseeländischen Klima eine Winterruheperiode durchmachen. Die Jungtiere sind anders als die Adulti tagaktiv, um nicht von großen Artgenossen gefressen zu werden. Nach einem Jahr bewohnen sie die Wohnkolonien der Adulti und gleichen ihnen in der Lebensweise. Weibchen legen nicht jedes Jahr Eier ab; nicht zuletzt deswegen ist zum Erhalt der Art ein hohes Individualalter nötig. Nach stark umstrittenen Angaben erreichen Brückenechsen ein Alter von bis zu 150 Jahren. Im Jahre 2009 war das älteste in Gefangenschaft gehaltene Exemplar 111 Jahre alt.[10]
Über die Entwicklung der Eier unter menschlicher Aufsicht ist man durch die Schutzaktion der Viktoria-Universität in Wellington gut unterrichtet. Im November gesammelte Eier von der Insel „The Brothers“ werden bei Temperaturen von 18 bis 23 Grad künstlich bebrütet, wodurch die Schlupfrate wesentlich erhöht wird. Da die Jungtiere dort nach nur sechs Monaten im Mai schlüpfen, lässt sich schließen, dass die Nester in freier Natur niedrigeren Temperaturen und größeren Schwankungen ausgesetzt sind.[11]
Offenbar wird die Verteilung der Geschlechter bei Brückenechsen direkt durch die Bruttemperatur beeinflusst. Bei Sphenodon punctatus schlüpften bei konstant 18 °C Bruttemperatur ausschließlich Weibchen, bei 20 °C 91 % Weibchen, bei 22 °C allerdings nur 23 %.[12]

Zusammenleben mit Seevögeln
Ein bemerkenswerter Aspekt der Biologie und Ökologie der Brückenechsen ist ihr Zusammenleben mit Seevögeln. Eine Annahme besagt, dass sie dort in einer fast symbiotischen Beziehung friedlich und höhlenteilend miteinander lebten. Allerdings überwiegt offenbar der Nutzen für das Reptil: Der Kot der Vögel und der Boden bilden die Nahrungsgrundlage für diverse Wirbellose, welche der Brückenechse als Nahrung dienen. Diese These ist in letzter Zeit verstärkt in Zweifel gezogen worden, da viele Indizien auf eine ausschließlich für die Brückenechse nützliche und für den Seevogel schädliche Beziehung hindeuten. In Erdhöhlen, in denen Brückenechsen lebten, wurden wiederholt angegriffene Küken ohne Kopf gefunden. Da in der Region dieser Funde keine Ratten oder ähnliche Tiere leben, ist die Brückenechse das einzige Tier, welches zur Tötung der Küken imstande ist. Brückenechsen dürften auch Gelege zertrampeln. Bei Beobachtungen flohen die kleineren Pinguin-Sturmtaucher (Pelecanoides urinatrix) immer aus ihren Höhlen, wenn Brückenechsen versuchten, in diese einzudringen. Sturmvögel von beträchtlicher Größe wie Puffinus griseus vertreiben Brückenechsen aus der Nähe ihrer Höhlen und sind dabei meist erfolgreich. Wenn Brückenechsen in den Höhlen von Seevögeln lebten, dann war die Höhle fast immer schon vom Vogel verlassen worden.
Vorkommen
Die Lebensräume der Brückenechsen sind durch ein eher raues Klima, starken Grasbewuchs und geringen Baumwuchs charakterisiert.
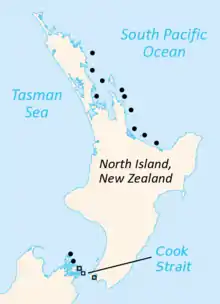
Punkte: Sphenodon punctatus
Quadrate: „Sphenodon guentheri“
Derzeit leben Brückenechsen nur noch auf etwa 30 kleinen neuseeländischen Inseln, die in der Cookstraße sowie zwischen der Bay of Plenty und der Bay of Islands entlang der Nordwestküste der nördlichen Hauptinsel liegen.
Durch den Menschen eingeschleppte Nagetiere sind der Grund für das Verschwinden der Art von den Hauptinseln Neuseelands.[1] Früher lebten die Reptilien auch auf der nördlichen Hauptinsel, ein ehemaliges Vorkommen auf der Südinsel ist umstritten.[3]
Bestandssituation
Die Brückenechsen wurden durch den Menschen stark dezimiert, vor allem durch von ihm eingeführte Ziegen, Katzen, Hunde, Schweine, Ratten und Mäuse sowie durch die Umwandlung ihres natürlichen Lebensraumes in Acker- und Weideland. Die Gattung Sphenodon bewohnt mittlerweile nur noch 0,1 % des ursprünglich von ihr bewohnten Areals. Die Art Sphenodon diversum ist nur aus Knochenfunden bekannt, von der Unterart S. punctatus reischeki wurde seit 1978 kein Exemplar mehr gesehen. Eine weitere Bedrohung für die Art ist Wilderei, da die Brückenechsen aufgrund ihrer Seltenheit und Exotik gefragte Terrarientiere sind.
Seit den neunziger Jahren des 20. Jahrhunderts wurden die Ratten, welche die Hauptbedrohung für die Brückenechse darstellten, auf nahezu sämtlichen von der Art bewohnten Inseln ausgerottet. Ebenso bemüht man sich um Wiederaufforstung und Renaturierung des Lebensraumes sowie um die Wiederansiedlung von Brückenechsen auf einst von ihnen bewohnten Inseln. Auch auf der Nordinsel Neuseelands gibt es mittlerweile wieder Bestände in größeren, umzäunten Gebieten. Diese Maßnahmen haben dazu geführt, dass sich der Bestand auf mittlerweile 55.000 erwachsene Exemplare erholt hat. Obwohl der Lebensraum der Brückenechsen stark fragmentiert ist und die Art auf kontinuierliche Schutzmaßnahmen angewiesen ist, wird sie von der IUCN derzeit als nicht gefährdet (least concern) eingestuft.[13]
Systematik
Interne Systematik
Eine Zeit lang wurde eine Population auf einigen kleinen Inseln in der Cookstraße in die eigene Art Sphenodon guentheri gestellt[14]. Inzwischen ergaben molekularbiologische Untersuchungen, dass die, obschon vorhandenen, genetischen Unterschiede nicht groß genug sind, um die Trennung in zwei Arten zu rechtfertigen; daher wurde Sphenodon guentheri mit Sphenodon punctatus synonymisiert.[15]
Sphenodon diversum wurde 1885 anhand eines einzelnen, unvollständigen und nicht fossilierten Skeletts, das aus einem Steinbruch geborgen worden war, von William Colenso beschrieben.[16] Lebende Exemplare wurden seitdem nicht gefunden.
Stammesgeschichte
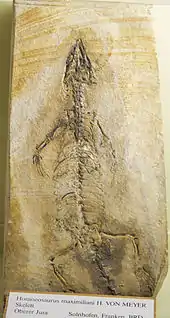
Brückenechsen werden oft als lebende Fossilien bezeichnet, da Vertreter aus der näheren Verwandtschaft der Brückenechsen (Sphenodontia), bereits aus dem Ladinium (obere Mitteltrias) vor circa 240 Millionen Jahren nachgewiesen.[17]
Die Brückenechse Sphenodon, die heute auf einigen neuseeländischen Inseln lebt, unterscheidet sich durch den noch vorhandenen Jochbogen des Schädels, der allen Eidechsen und Schlangen fehlt, von sämtlichen Kleinreptilien. Diese Nachweis ermöglicht die Zuordnung zu einer altertümlichen Gruppe, die in Trias und Jura weltweit anzutreffen war. Ab dem Zeitalter der Oberkreide sind keine Fossilien mehr bekannt.[1]
Sphenodontier waren somit in der Obertrias und im Jura weltweit verbreitet, wurden jedoch ab der frühen Kreide zunehmend von „modernen“ Reptilien, speziell den Schuppenkriechtieren (Squamata), zurückgedrängt und konnten sich nur auf den isolierten neuseeländischen Inseln bis heute halten. Der Grund für die anfängliche Dominanz der Sphenodontier und den plötzlichen Aufschwung der Squamaten ist unklar, weil beide Entwicklungslinien Schwestergruppen und damit gleich alt sind.
Die Sphenodontia lassen sich in zwei Familien unterteilen: Sphenodontidae und Pleurosauridae. Die Sphenodontiden sind terrestrische Formen. Die rezenten Brückenechsen gehören dieser Familie an und der älteste bekannte Sphenodontier in der späten Mitteltrias dürfte ebenfalls dieser Familie angehört haben. Einer der jüngsten Vertreter der Blütezeit der Sphenodontier ist die relativ gut bekannte Gattung Homoeosaurus („Scheinbrückenechsen“) aus dem Oberjura, eine Gruppe, die sich kaum von den heutigen Brückenechsen unterschied. Die Sphenodontiden sind somit seit mindestens 150 Millionen Jahren morphologisch ausgesprochen konservativ. 2020 wurde das Genom erstmals sequenziert, wobei Anzeichen stetiger molekularer Evolution sowie leichter morphologischer Evolution nachgewiesen wurden. Das Genom der Spezies enthält Sequenzen, die bisher nur jeweils in Reptilien oder Säugetieren gefunden wurden, wodurch die phylogenetische Verwandtschaft der Säugetiere und Reptilien bestätigt wurde.[18]
Die Vertreter der Pleurosauriden gingen hingegen zur aquatischen Lebensweise über. Eine dieser Formen ist Pleurosaurus goldfussi. Fossilien dieses bis zu 1,5 Meter langen Reptils, dessen Schwanzwirbelsäule doppelt so lang ist wie der übrige Körper, wurden, wie auch Überreste des Sphenodontiden Homoeosaurus maximiliani, in den oberjurassischen Plattenkalken von Solnhofen (Fränkische Alb)[19] und den Plattenkalken von Canjuers (Département Var, Frankreich)[20] gefunden. Der Fossilbericht der Pleurosauriden ist jedoch im Gegensatz zu dem der Sphenodontiden auf den Jura beschränkt.
Haltung
Da die Tiere strengsten Schutz durch die neuseeländische Regierung genießen, werden derzeit (Stand März 2011) nur etwa 140 Brückenechsen in zehn Institutionen gehalten, davon 43 außerhalb Neuseelands. In Deutschland werden sie nur im Aquarium des Berliner Zoos gezeigt. Dort leben (Stand Mai 2021) sieben Exemplare in einem großen, gekühlten Terrarium.
In Unkenntnis der speziellen Temperaturbedürfnisse von Brückenechsen wurde der Großteil der Tiere früher zu warm gehalten. Eine der ersten in Europa oder auch weltweit gehaltenen Brückenechsen war im zoologischen Institut der Universität Uppsala präsent, wo sie im Herbst 1908 ankam. Sie lebte dort bis Sommer 1931 in einer 75 × 40 × 40 cm großen Holzkiste unter einem Schreibtisch, die gesamte Kiste war nur mit Holzwolle ausgekleidet und enthielt ein Wassergefäß. Die Temperaturen schwankten zwischen 16 und 20 °C. 1931 kam sie in eine 190 × 75 × 30 cm große Holzkiste, welche strukturiert eingerichtet war. Dort zeigte sie größere Aktivitäten. Sie wurde wöchentlich mit ein paar Fleischstreifen oder 15 bis 20 Regenwürmern gefüttert.[21]
Das Tier in der Belletristik
In John Greens Roman Schlaft gut, ihr fiesen Gedanken vermacht ein Mann in seinem Testament sein ganzes Vermögen seiner Tuatara.
Literatur
- Wolfgang Böhme: Sphenodontida, Schnabelköpfe, Brückenechsen. In: Wilfried Westheide und Reinhard Rieger (Hrsg.): Wirbel- oder Schädeltiere. Fischer, Stuttgart 2004, ISBN 3-8274-0900-4 (Spezielle Zoologie. Teil 2), S. 354–357
- K. Klemmer: Die Brückenechse. In: Bernhard Grzimek (Hrsg.): Kriechtiere. Bechtermünz, Augsburg 2000, ISBN 3-8289-1603-1 (Grzimeks Tierleben. Band 6), S. 148–151 (unveränderter Nachdruck der dtv-Ausgabe von 1979/80)
- Manfred Rogner: Brückenechsen, Schnabelkopf-Echsen (Ordnung Rynchocephalia). In: Manfred Rogner: Warane, Skinke und andere Echsen sowie Brückenechsen und Krokodile. Ulmer, Stuttgart 1994, ISBN 3-8001-7253-4 (Echsen. Band 2), S. 220–223.
- Robert L. Carroll: Paläontologie und Evolution der Wirbeltiere. Thieme, Stuttgart 1993, ISBN 3-13-774401-6.
Weblinks
- Sphenodon In: The Reptile Database
- Sphenodon punctatus und Sphenodon guntheri in der Roten Liste gefährdeter Arten der IUCN 2008. Eingestellt von: Australasian Reptile & Amphibian Specialist Group, 1996. Abgerufen am 27. April 2015.
- Brückenechse, auf aquarium-berlin.de
- J. Robert Macey et al.: Evidence of two deeply divergent co-existing mitochondrial genomes in the Tuatara reveals an extremely complex genomic organization, in: Commun Biol Band 4, Nr 116, 29. Januar 2021, doi:10.1038/s42003-020-01639-0. Dazu:
- Devin A. Reese: Lizard-like tuatara carry two distinct mitochondrial genomes, auf: ScienceNews vom 29. Januar 2021
Fußnoten
- Riesensalamander und Brückenechsen Freie Universität Berlin, aufgerufen am 6. Dezember 2021
- Bradford Haami: Ngārara – reptiles. In: Te ara – The encyclopedia of New Zealand. Ministry for Culture & Heritage (New Zealand), 22. September 2012, abgerufen am 6. Oktober 2013 (englisch).
- Nach W. Böhme (2004), S. 354.
- Die kauende und sägende Echse. In: Deutschlandfunk. Abgerufen am 10. Juli 2012.
- Nach einem Bericht von Robert Mertens, zitiert in Grzimek (2000), S. 150
- Nach Grzimek (2000), S. 149
- Nach Rogner (1994), S. 222; sowie W. Böhme (2004), S. 357
- Nach W. Böhme (2004), S. 357
- Nach einem Bericht von Eugen Schuhmacher, zitiert in Grzimek (2000), S. 150–151
- Echse wird mit 111 Jahren Vater. Frankfurter Rundschau, 26. Januar 2009, abgerufen am 6. Oktober 2013.
- Nach Rogner (1994), S. 221
- Nach http://www.aquarium-berlin.de/erleben/tier-highlights/brueckenechse.html
- Sphenodon punctatus in der Roten Liste gefährdeter Arten der IUCN 2019. Eingestellt von: Hitchmough, R., 2019. Abgerufen am 02. April 2020.
- Buller,W.L. 1877. Notes on the tuatara lizard (Sphenodon punctatum), with a description of a supposed new species. Transactions and Proceedings of the New Zealand Institute 9: 317–325
- Hay, Jennifer M.; Stephen D. Sarre, David M. Lambert, Fred W. Allendorf and Charles H. Daugherty. Genetic diversity and taxonomy: a reassessment of species designation in tuatara (Sphenodon: Reptilia). In: Conservation Genetics. 11, Nr. 3 2010: S. 1063–1081
- W. Colenso: Notes on the Bones of a Species of Sphenodon apparently distinct from the Species already known. In: Transactions and Proceedings of the Royal Society of New Zealand. 18, 1885, S. 118–128 (PDF 525 kB)
- Marc E. H. Jones, Cajsa Lisa Anderson, Christy A. Hipsley, Johannes Müller, Susan E. Evans, Rainer R. Schoch: Integration of molecules and new fossils supports a Triassic origin for Lepidosauria (lizards, snakes, and tuatara). BMC Evolutionary Biology. Bd. 13, Nr. 1, 2013, Art.-Nr. 208, doi:10.1186/1471-2148-13-208 (Open Access)
- Ngatiwai Trust Board, Neil J. Gemmell, Kim Rutherford, Stefan Prost, Marc Tollis: The tuatara genome reveals ancient features of amniote evolution. In: Nature. Band 584, Nr. 7821, 20. August 2020, ISSN 0028-0836, S. 403–409, doi:10.1038/s41586-020-2561-9 (nature.com [abgerufen am 2. September 2020]).
- Günter Viohl: Die Solnhofener Plattenkalke, oberer Jura. In: Dieter Meischner (Hrsg.): Europäische Fossillagerstätten. Springer, Berlin·Heidelberg 2000, S. 143–150, doi:10.1007/978-3-642-57198-5_15
- Karin Peyer, Sylvain Charbonnier, Ronan Allain, Émilie Läng, Renaud Vacant: A new look at the Late Jurassic Canjuers conservation Lagerstätte (Tithonian, Var, France). Comptes Rendus Palevol. Bd. 13, Nr. 5, 2014, S. 403–420, doi:10.1016/j.crpv.2014.01.007 (alternativer Volltextzugriff: ResearchGate)
- Nach Cyren (1934), zitiert in Rogner (1994), S. 223