Nicotiana attenuata
Nicotiana attenuata ist eine Pflanzenart aus der Gattung Tabak (Nicotiana). Innerhalb der Gattung wird sie in die Sektion Petunioides eingeordnet, die am nächsten verwandte Art ist Nicotiana acuminata. Die bis zu 150 Zentimeter hoch werdende, einjährige Pflanze ist auffällig mit drüsigen Pflanzenhaaren (Trichomen) bedeckt. Die Blüten öffnen sich meist in der Nacht, die Krone ist schmal trichterförmig und weiß gefärbt. Damit ist die Art auf eine Bestäubung durch nachtaktive Schmetterlinge angepasst.
| Nicotiana attenuata | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Nicotiana attenuata | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Nicotiana attenuata | ||||||||||||
| Torr. ex S.Watson |
Das Verbreitungsgebiet der Art liegt im Südwesten der Vereinigten Staaten. Dort ist sie vor allem an Standorten zu finden, an denen kurz zuvor Waldbrände auftraten. Unter anderem aufgrund der Anpassung an diesen speziellen Lebensraum dient die Pflanze als Modellorganismus zur Erforschung der Beziehung zwischen Pflanzen und ihrer Umwelt. Von einigen indianischen Völkern wurden die Pflanzen als Rauchtabak verwendet, archäologische Nachweise über eine solche Verwendung können auf die Zeit um 650 n. Chr. datiert werden.
Beschreibung
Vegetative Merkmale

Nicotiana attenuata ist eine aufrecht wachsende, einfache oder verzweigte, stark riechende,[1] einjährige Pflanze, die 50 bis 150 Zentimeter hoch wird. Ist sie verzweigt, ist der mittlere Stängel meistens am kräftigsten ausgebildet. Die nahe dem Boden befindlichen Laubblätter bilden eine Rosette, sie sind leicht fleischig und werden 5 bis 10 Zentimeter lang. Sie sind elliptisch, eiförmig elliptisch oder elliptisch langgestreckt geformt. Nach vorn sind sie stumpf oder selten auch spitz. Die Blattstiele sind meist etwa halb so lang wie die Blattspreiten. Im Alter von zwei bis sechs Wochen beginnt sich die Sprossachse zu verlängern.[2] Die oberhalb der grundständigen Rosette gebildeten Laubblätter haben zunehmend kürzere Stiele, die Blattspreiten werden dabei immer schmaler, so dass ihre Form lanzettlich bis linealisch-lanzettlich wird, nach vorn werden sie mehr und mehr spitz.[3]
Die Pflanze ist drüsig mit zwei verschiedenen Typen von Trichomen behaart: einem längeren, gestielten Typ, sowie einem etwas kürzeren, ebenfalls gestielten Typ. Auf den Laubblättern tritt der erste Typ gleichmäßig auf beiden Seiten auf, der zweite Typ ist jedoch meist nur entlang der Blattachse zu finden.[4] Im Alter kann die Pflanze etwas verkahlen.[5] Die Trichome besitzen eine geschwollene Basis, die an lebenden Pflanzen wie Tropfen einer Flüssigkeit aussieht, bei Herbarexemplaren zu einer feinen kraterförmigen Erhebung zusammenfällt.[3]
Blütenstände und Blüten

Die Blütenstände sind kurz traubenförmig oder schmal rispenförmig, sie werden von linealischen,[5] mehr oder weniger geraden Tragblättern begleitet, die kürzer als 30 Millimeter[6] sind. Die Blütenstiele sind zunächst 2 bis 4 Millimeter lang, später verlängern sie sich auf 4 bis 7 Millimeter. Die Blüten besitzen aus fünf verwachsenen Kelchblättern bestehende Blütenkelche mit einer Länge von 7 bis 10 Millimetern. Auf der Außenseite befinden sich auffällige Trichome, die ebenfalls eine verdickte Basis aufweisen. Die Kelchröhre ist mit fünf dreieckigen, ungleich gestalteten Zipfeln besetzt, die größeren erreichen eine Länge von etwa 1,5 bis 4,5[1] Millimetern.[3]
Die Krone besteht aus fünf verwachsenen Kronblättern, sie ist schmal trichterförmig und weiß gefärbt.[5] Die Kronröhre erreicht eine Länge von 20 bis 27 (selten bis 32) Millimetern. Sie teilt sich in einen schmalen Bereich an der Basis mit einer Länge von 5 Millimetern und einem Durchmesser von 1,5 bis 2 Millimetern, sowie einen Kronschlund, der nicht ganz den doppelten Durchmesser erreicht. Dieser Kronschlund ist nahezu zylindrisch, nach oben ist er leicht asymmetrisch verschoben. Die Außenseite ist fein flaumhaarig, blass grünlich und leicht pink überhaucht. Der weiß gefärbte Kronsaum misst 4 bis 6 Millimeter im Durchmesser und ist nur mit schwach ausgeprägten Kronlappen besetzt. Diese sind 2 bis 3 Millimeter lang,[5] stumpf oder nur selten spitz. Die oberen drei sind leicht zurückgebogen, die unteren zwei etwas aufsteigend. Die fünf Staubblätter setzen an der Basis der Kronschlunds an, die Staubfäden sind direkt oberhalb dieser Ansatzstelle fein flaumig behaart und leicht geschwungen. Die Länge der Staubblätter ist unterschiedlich: Bei einem Paar reichen die Staubbeutel bis an die Öffnung der Kronröhre, das zweite Paar ist kürzer und schon deutlich von der Kronröhre umschlossen und das fünfte Staubblatt ist nochmals kürzer.[3]
Die Blüten öffnen sich meistens am Abend. Gelegentlich werden aber auch Blüten gebildet, die sich am Morgen öffnen. Diese sind deutlich kleiner, der Durchmesser der Krone beträgt nur etwa ein Drittel der sich abends öffnenden Blüten.[7]
Früchte und Samen
Die Frucht ist eine 8 bis 12 Millimeter lange, eiförmige, nach vorn zugespitzte Kapsel, an deren Spitze vier Kammern gebildet werden[5] und die etwa 16 Tage nach der Befruchtung der Blüte reift.[8] Sie enthält zwischen 10 und 300 Samen,[8] die mehr oder weniger nierenförmig oder gewinkelt nierenförmig sind und bis zu 0,7 Millimeter lang werden. Sie besitzen eine matt grau-braune, geriffelt netzförmige Oberfläche. Das Embryo ist halbkreisförmig.[3]
Verbreitung und Standorte
Die Art kommt im Südwesten Nordamerikas vor. Die nördlichste Verbreitung liegt bei etwa 50° nördlicher Breite in British Columbia (Kanada), im Süden reicht sie bis 29° nördlicher Breite in Mexiko. Im Westen ist sie in Kalifornien und auf der Insel Guadalupe vor Niederkalifornien zu finden, im Osten reicht das Verbreitungsgebiet über Montana, Wyoming, Colorado und New Mexico bis in das westliche Sonora. Innerhalb dieses Gebietes ist Nicotiana attenuata besonders häufig in der Vegetation des Großen Beckens zu finden, was dem Gebiet vom südlichen Oregon und Idaho, östlich der Sierra Nevada bis zum Owens Valley, entlang Nevadas und bis nach New Mexico entspricht.[9] Die Art wächst in Abschwemmungen, an sandigen Hängen, Straßenrändern und Feldern.[5]
Ökologie
Dormanz und Keimung nach Feuern
Die Pflanzen treten meist nur wenige (ein bis drei) Jahre nach Feuern in Wüsten-Beifuß- und Pinyon-Kiefern/Wacholder-Wäldern auf, nur einzelne Populationen überdauern für mehrere Vegetationsperioden in Abschwemmungen und als Unkraut an neu gebauten Straßen. Nach einem Feuer eines bis zu 150 Jahre alten Wacholder-Bestandes 1994 in Utah auf einer Fläche von über 800 Hektar wurden im nachfolgenden Jahr mehr als 10000 Einzelpflanzen von Nicotiana attenuata festgestellt. Da die nächste Population über 8 Kilometer entfernt ist und keine Ausbreitungsmechanismen der Samen bekannt sind, wird vermutet, dass die Pflanzen aus Samen gewachsen sind, die eine über 100-jährige Dormanz (Samenruhe) überdauert haben. Diese Dormanz wird über eine Kombination aus positiven und negativen Signalen gesteuert: Das Keimen wird durch Stoffe unterbunden, die beim Verrotten von – potentiell um Nährstoffe konkurrierenden – Pflanzen entstehen, wobei die eigentliche Keimfähigkeit der Samen nicht beeinträchtigt wird; gefördert wird das Keimen durch Stoffe, die beim Verbrennen von Zellulose entstehen. Ein Vorteil, den die Pflanze dadurch hat, liegt im Verschwinden potentieller Konkurrenten; zudem kann die erhöhte Ammoniakkonzentration, die nach Bränden im Boden vorhanden ist, zur Synthese von Nikotin[10] ausgenutzt werden.[11]
Bei Experimenten, in denen getopfte Nicotiana attenuata-Pflanzen zum einen an kurz zuvor abgebrannte Wacholder-Standorte und zum anderen an nicht abgebrannte Standorte gestellt wurden, konnte nachgewiesen werden, dass die Menge an Blattfläche, die von Herbivoren gefressen wurden, an ersteren Standorten ein bis 20 mal geringer war. Die Anpassung an diese besonderen Standorte ist daher als ein Schutz gegenüber Fraßfeinden anzusehen.[11]
Bestäuber
Zu den Bestäubern von Nicotiana attenuata gehören Schwärmer der Gattung Manduca (vor allem der Tabakschwärmer (Manduca sexta) und Manduca quinquemaculatus), sowie Kolibris, hier vor allem die Art Archilochus alexandri, aber auch die Rotrücken-Zimtelfe (Selasphorus rufus). Beide Gruppen von Bestäubern besuchen die Blüten, um den zuckerhaltigen Nektar zu sammeln. Neben Zucker enthält diese von der Pflanze bereitgestellte Belohnung jedoch eine große Anzahl an flüchtigen organischen Verbindungen, die über Geruch und Geschmack das Verhalten der Bestäuber beeinflussen. Experimente haben gezeigt, dass dabei Benzylaceton der wichtigste Bestandteil des Nektars ist, um die Bestäuber anzulocken, Nikotin jedoch abstoßend wirkt und die Menge des von den Bestäubern aufgenommenen Nektars reduziert. Die Präsenz von Nikotin im Nektar der Pflanze wurde zunächst meist als Nebenwirkung der Anreicherung des Stoffs in den Blättern zur Abwehr von Fraßfeinden angesehen. Jedoch wird dadurch auch die Anzahl der von einem Bestäuber besuchten Blüten erhöht, was wiederum für eine höhere Auskreuzungsrate und damit für eine geringere Inzuchtdepression der Population sorgt.[12][13]
Abwehr von Fraßfeinden

Als natürliche Feinde von Nicotiana attenuata treten vor allem Raupen verschiedener Schmetterlinge auf. Dabei gibt es zum einen auf Tabak-Arten spezialisierte Schmetterlingsarten wie den Tabakschwärmer (Manduca sexta), aber auch nicht näher spezialisierte Arten wie Spodoptera exigua und Spodoptera littoralis. Es werden vor allem die Laubblätter gefressen, in geringerem Umfang auch Blüten und Früchte.[14] Schäden werden jedoch auch von Wirbeltieren verursacht, die meist die Pflanze oberhalb der Rosette abfressen. Beobachtet wurden Erdhörnchen-Arten der Gattungen der Ziesel (Spermophilus) und Antilopenziesel (Ammospermophilus), der Kalifornische Eselhase (Lepus californicus) sowie das Audubon-Baumwollschwanzkaninchen (Sylvilagus audubonii).[2]
Als effektivste Abwehr gegen die meisten unspezialisierten Fraßfeinde gilt das in der Pflanze angereicherte Nikotin, welches als Nervengift wirkt. Wird die Pflanze befallen, steigert sich die Nikotinproduktion auf etwa das Zwei- bis Vierfache (in Einzelfällen auf das mehr als 14-fache),[2] um die Fraßfeinde abzuwehren. Besonders hoch ist die Konzentration an Nikotin in den Trichomen des Kelchs, ein einzelnes dieser Pflanzenhaare kann zwischen 1,3 und 3,6 Mikrogramm des Alkaloids enthalten.[10]
Als Nebeneffekt der Nikotinanreicherung kann die Pflanze nur eine geringere Menge an Samen erzeugen, jedoch sind diese Einbußen für die Pflanze geringer als der Verlust an Samen, wenn keine Reaktion auf den Befall erfolgt.[15] Selbst die Raupen des spezialisierten Tabakschwärmers wachsen langsamer, wenn sie nikotinhaltige Blätter fressen, jedoch dient das aufgenommene Nikotin wiederum als Schutz gegen die Feinde der Raupen. Wird die Nikotinproduktion in den Pflanzen unterdrückt, steigt der Befall mit einer Vielzahl von Insekten, die sonst nur eine weniger bedeutende Rolle als Fraßfeind spielen. Dazu gehören beispielsweise der Eulenfalter Spodoptera exigua, Vertreter der Heuschrecken-Gattung Trimerotropis und der Blattkäfer Epitrix hirtipennis.[16]
Es konnte gezeigt werden, dass die oralen Ausscheidungen der Raupen des Tabakschwärmers die Produktion von flüchtigen Inhaltsstoffen des Blattgrüns verändern. An rein mechanisch verletzten Blättern treten mehr (Z)-Isomere als (E)-Isomere auf, während sich bei Blättern, die von Tabakschwärmerraupen befallen werden, das Verhältnis mehr und mehr ausgleicht. Diese Veränderung dient räuberisch lebenden Arten der Gattung Geocoris aus der Familie der Geocoridae als Signal, dass sich auf den Pflanzen Beute in Form von Eiern und Larven des Tabakschwärmers befinden.[17]
Wechsel der bevorzugten Bestäuber
Die Nachtfalter der Gattung Manduca sind die primären Bestäuber der Art. Da die ausgewachsenen Falter dadurch wichtig für das Fortbestehen der Art sind, die Raupen jedoch gleichzeitig als Fraßfeind die Pflanzen bedrohen, ist in Nicotiana attenuata ein Regulierungsprozess ausgebildet, mit dem ein übermäßiger Befall von Raupen verhindert werden soll. Fressen die Raupen an den Pflanzen, wird als Reaktion auf die oralen Ausscheidungen und Regurgitantien die Produktion des Pflanzenhormons Jasmonsäure ausgelöst.[18] Dieses bewirkt, dass die Pflanze den Blührhythmus umstellt: Öffnen sich die Blüten normalerweise in den Abendstunden (zwischen 18 und 22 Uhr), um in der Nacht von Faltern bestäubt zu werden, werden bei einem hohen Befall von Schmetterlingsraupen mehr und mehr Blüten gebildet, die sich am Morgen (zwischen 6 und 10 Uhr) öffnen. Diese Blüten sind kleiner, ihr Duft enthält weniger Benzylaceton und ihr Nektar geringere Konzentration an Zucker. Diese Anpassung der Blüten hat zur Folge, dass sie nun vor allem von Kolibris bestäubt werden. Es wird vermutet, dass eine komplette Anpassung auf eine Bestäubung durch Kolibris nachteilig wäre, da sie nicht wie Nachtfalter mittels Blütendüften über weite Strecken heran gelockt werden können und da an den Standorten der Art, die meist nach Feuern bewachsen werden, Nistplätze der Vögel oftmals nicht vorhanden sind.[7]
Systematik
Innerhalb der Gattung Nicotiana wird Nicotiana attenuata in die Sektion Petunioides eingeordnet. Alle Arten der Sektion sind einjährige Kräuter mit einer Chromosomenzahl von n = 12 und diploiden Chromosomensatz, die meisten Arten sind im Südwesten der Anden verbreitet. Am nächsten verwandt ist Nicotiana attenuata mit Nicotiana acuminata. Aufgrund der hohen Ähnlichkeit und den im Gegensatz dazu voneinander entfernt liegenden Verbreitungsgebieten wird angenommen, dass Nicotiana attenuata erst relativ spät durch Verbreitung von Samen über hohe Distanzen nach Nordamerika gelangte.[19][20] Weiterhin wird angenommen, dass vor etwa 1 Million Jahren die allopolyploide Sektion Polydicliae aus den Vorfahren von Nicotiana obtusifolia als männliche Elternart und den Vorfahren von Nicotiana attenuata als weibliche Elternart entstanden ist.[21][22] 2010 wurden zwei künstlich erzeugte Hybride beschrieben, die diesen Prozess nachstellen sollen: Mit Nicotiana miersii als zweite Elternart wurde Nicotiana × mierata und mit Nicotiana obtusifolia Nicotiana × obtusiata erzeugt.[23][24]
Verwendung
Die Verwendung von Nicotiana attenuata als Rauchtabak ist von einigen Indianerstämmen im Verbreitungsgebiet dokumentiert, so unter anderem von den Washoe, Cahuilla, Kumeyaay, Zuñi, Ute, Gosiute und den nördlichen Blackfoot. Oftmals wird die Art in diesem Zusammenhang „Kojotentabak“ genannt. Teilweise wurden die Pflanzen in der Nähe der Siedlungsstätten kultiviert.[25] Bei Ausgrabungen historischer Siedlungsstätten aus der Basketmaker-III-Epoche (um 650 n. Chr.) im Norden Arizonas wurden kleine Bündel gefunden, die unter anderem Pflanzenreste enthielten, die als Nicotiana attenuata identifiziert wurden. Diese Pflanzen wurden wahrscheinlich zu medizinischen Zwecken eingesetzt, zudem konnte nachgewiesen werden, dass dieser Tabak in Pfeifen geraucht wurde.[26][27]
In der ethnobotanischen Literatur des 20. Jahrhunderts findet sich eine Vielzahl weiterer Verwendungsnachweise der Art. Dazu gehört der Einsatz gegen Nasenbluten, Erkältungen, Nesselsucht, Masern, Tuberkulose, Schlangenbisse, unterschiedliche Schmerzen wie Kopfschmerzen, Zahnschmerzen oder Rheuma und gegen den Befall durch verschiedene Würmer. Die Art der dokumentierten Anwendungen reicht vom Rauchen der Blätter über das Einreiben mit einem aus den Pflanzen gekochten Sud bis hin zum Auftragen gekauter Blätter auf zu heilende Körperteile. Verwendet beim Haarewaschen soll die Pflanze gegen Schuppen und Haarausfall helfen.[28]
Forschungsgeschichte
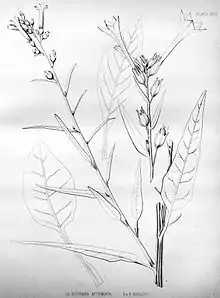
Die nach den Regeln des Internationalen Codes der Nomenklatur für Algen, Pilze und Pflanzen (ICBN) gültige Erstbeschreibung von Nicotiana attenuata wurde 1871 von Sereno Watson im fünften Band der Reihe „United States Geological Exploration of the Fortieth Parallel“ verfasst. Dort gibt er als Autor des Namens John Torrey an, als Quelle des Namens nennt er „in Herb.“ (im Herbarium).[29] Nach Artikel 32.1 ICBN muss ein gültig beschriebenes Taxon eine Beschreibung oder Diagnose enthalten, was bei einem Herbarbeleg nicht gegeben ist. Das sich daraus ergebende Autor-Zitat lautet Torr. ex S.Watson. Dass der Name bereits 1821 von Ernst Gottlieb von Steudel verwendet wurde, ist nach Artikel 34.1 ICBN nicht relevant, da es sich um ein nomen nudum handelt.[30] Als Holotypus gilt ein Herbarbeleg von John Torrey mit der Belegnummer Torrey 354, der sich im Herbarium des New York Botanical Garden befindet. Ein Isotypus gehört zur Sammlung des Asa Gray Herbariums an der Harvard University. Die Belege stammen wahrscheinlich von einem erwachsenen, jedoch noch nicht sehr alten Individuum: es fehlt die bodenständige Laubblattrosette, auch die Blütenstände sind an älteren Pflanzen meist deutlicher ausgeprägt.[3]
Beschreibungen der Art wurden in verschiedenen Florenwerken veröffentlicht, unter anderem in „Synopsis of the Flora of Colorado“ (1874),[31] „Synoptical Flora of North America“ (1878),[32] „A popular California Flora“ (1882),[33] „A Flora of western middle California“ (1901),[34] „Flora of Colorado“ (1905),[35] „Elemental Flora of the Northwest“ (1914),[36] „Flora of southeastern Washington and adjacent Idaho“ (1914),[37] „Flora of New Mexico“ (1914),[38] „Flora of southern British Columbia and Vancouver Island“ (1915),[39] „Flora of the Rocky Mountains and adjacent plains“ (1917),[40] „A Flora of California“ (1943)[41] und „An illustrated flora of the Pacific States“ (1951).[5]
Einen großen Beitrag zur Erforschung der gesamten Gattung Nicotiana lieferten die Arbeiten von Thomas Harper Goodspeed, der die Ergebnisse seiner Arbeit 1954 in der Monographie The Genus Nicotiana zusammenfasste. Dieses Buch und die darin verwendete Systematik und die taxonomischen Abhandlungen galten lange Zeit als Referenz zur Gattung. Die Einteilung in Untergattungen und Sektionen wurde sowohl in der botanischen als auch agrikulturellen Literatur verwendet.[20] Nicotiana attenuata wird darin der Untergattung Petunioides zugeschrieben und innerhalb dieser Untergattung der Sektion Acuminatae.[3] Die Zuordnung zu den Sektionen konnte in den meisten Fällen durch molekularbiologische Untersuchungen bestätigt werden, jedoch ist der von Goodspeed verwendete Sektionsname Acuminatae ungültig, Vorrang hat der ältere Name Petunioides.[20] Diese molekularbiologischen Untersuchungen dienten der Erforschung der phylogenetischen Zusammenhänge sowohl innerhalb der Gattung Nicotiana als auch der gesamten Familie der Nachtschattengewächse. Erstmals wurde Nicotiana attenuata von Richard Olmstead und Jeffery Palmer mit diesen Methoden untersucht, ihre Ergebnisse des Vergleichs der Restriktionsenzyme der DNS der Chloroplasten wurden 1991 veröffentlicht.[42] Weiterhin wurden unter anderem in verschiedenen Untersuchungen die Gene ndhF[43][19] und matK,[44][19] sowie die DNS-Spacer trnL-F[43][19] und trnS-G[19] sequenziert.[45]
Seit Ende der 1980er Jahre wird Nicotiana attenuata verstärkt als Modellorganismus zur Erforschung der Interaktion zwischen Pflanzen und ihrer Umwelt eingesetzt. Vor allem aufgrund ihrer Spezialisierung auf das Leben in einer ökologischen Nische – dem Lebensraum in abgebrannten Busch- und Baumbeständen – ist die Art ein geeignetes Untersuchungsobjekt. Ausschlaggebend für diese Entwicklung sind vor allem die Arbeiten von Ian T. Baldwin, der zunächst an der State University of New York die Forschungen begann. 1995 wurde ihm durch die Max-Planck-Gesellschaft die Gelegenheit geboten, eine Stelle als Gründungsdirektor des Max-Planck-Institut für chemische Ökologie in Jena anzunehmen und dort seine Forschungen weiterzuführen. Das Institut betreibt neben den Einrichtungen und Gewächshäusern in Jena auch eine Versuchsstation in Utah im natürlichen Verbreitungsgebiet der Art, um dort Feldversuche durchzuführen.[46]
Literatur
- Thomas Harper Goodspeed: The Genus Nicotiana: Origins, Relationships and Evolution of its Species in the Light of their Distribution, Morphology and Cytogenics. 1954. (Nachdruck von A.J. Reprints Agency, Neu-Delhi 1982)
Weblinks
- Scan des Holotyps beim New York Botanical Garden
Nachweise
- Washington Natural Heritage Program: Nicotiana attenuata (PDF; 172 kB). In: List of Vascular Plants Tracked by the Washington Natural Heritage Program. Verfasst 1999, abgerufen am 1. September 2012.
- Ian T. Baldwin, Thomas E. Ohneiss: Alkaloidal Responses to Damage in Nicotiana Native to North America. In: Journal of Chemical Ecology. Band 19, Nummer 6, 1993, S. 1143–1153. doi:10.1007/BF00987376
- T. H. Goodspeed, H.-M. Wheeler, P. C. Hutchinson: Nicotiana attenuata, Taxonomic Treatment In: Thomas Harper Goodspeed (Hrsg.): The Genus Nicotiana: Origins, Relationships and Evolution of its Species in the Light of their Distribution, Morphology and Cytogenics. 1954. Nachdruck von A.J. Reprints Agency, Neu-Delhi 1982, S. 427–430.
- Alexander Weinhold u. a.: Phaseoloidin, a Homogentisic Acid Glucoside from Nicotiana Attenuata Trichomes, Contributes to the Plant’s Resistance against Lepidopteran Herbivores. In: Journal of Chemical Ecology. Band 37, Nummer 10, 2011, S. 1091–1098. doi:10.1007/s10886-011-0012-7
- Leroy Abrams: An illustrated flora of the Pacific States. Band 3, Stanford University Press, 1951, S. 684. doi:10.5962/bhl.title.4657
- Michael H. Nee: Nicotiana attenuata. In: Jepson Flora Project, Jepson eFlora. 2012. Abgerufen am 6. September 2012.
- Danny Kessler, Celia Diezel, Ian T. Baldwin: Changing Pollinators as a Means of Escaping Herbivores. In: Current Biology. Band 30, Februar 2010, S. 237–242. doi:10.1016/j.cub.2009.11.071
- Michael Euler, Ian T. Baldwin: The chemistry of defense and apparency in the corollas of Nicotiana attenuata. In: Oecologia. Band 107, Nummer 1, 1996, S. 102–112. doi:10.1007/BF00582240
- Philip V. Wells: An Ecological Investigation of two Desert Tobaccos. In: Ecology. Band 40, Nummer 4, Oktober 1959, S. 626–644. doi:10.2307/1929816
- Ian T. Baldwin: Inducible Nicotine Production in Native Nicotiana as an Example of Adaptive Phenotypic Plasticity. In: Journal of Chemical Ecology. Band 25, Nummer 1, 1999, S. 3–30. doi:10.1007/BF00987376
- Catherine A. Preston, Ian T. Baldwin: Positive and Negative Signals regulate Germination in the post-fire Annual, Nicotiana attenuata. In: Ecology. Band 80, Heft 2, 1999, S. 481–494. doi:10.1890/0012-9658(1999)080[0481:PANSRG]2.0.CO;2
- Danny Kessler, Ian T. Baldwin: Making sense of nectar scents: the effects of nectar secondary metabolites on floral visitors of Nicotiana attenuata. In: The Plant Journal. Band 49, 2006, S. 840–854. doi:10.1111/j.1365-313X.2006.02995.x
- Danny Kessler, Klaus Gase, Ian T. Baldwin: Field Experiments with Transformed Plants Reveal the Sense of Floral Scents. In: Science. Band 321, 29. August 2008, S. 1200–1202. doi:10.1126/science.1160072
- Andrew McCall, Richard Karban: Induced defense in Nicotiana attenuata (Solanaceae) fruit and flowers. In: Oecologia. Band 146, Nummer 4, 2006, S. 566–571. doi:10.1007/s00442-005-0284-0
- Ian T. Baldwin: Jasmonate-induced responses are costly but benefit plants under attack in native populations. In: Proceedings of the National Academy of Science of the USA, Ecology. Band 95, Juli 1998, S. 8113–8118.
- Anke Steppuhn u. a.: Nicotine’s Defensive Function in Nature. In: PLOS Biology. Band 2, Ausgabe 8, August 2004, S. 1074–1080. doi:10.1371/journal.pbio.0020217
- Silke Allmann, Ian T. Baldwin: Insects Betray Themselves in Nature to Predators by Rapid Isomerization of Green Leaf Volatiles. In: Science. Band 329, 27. August 2010, S. 1075–1078. doi:10.1126/science.1191634
- Mario Kallenbach, Fiammetta Alagna, Ian T. Baldwin, Gustavo Bonaventure: Nicotiana attenuata SIPK, WIPK, NPR1, and Fatty Acid-Amino Acid Conjugates Participate in the Induction of Jasmonic Acid Biosynthesis by Affecting Early Enzymatic Steps in the Pathway. In: Plant Physiology. Band 152, Nummer 1, Januar 2010, S. 96–106. doi:10.1104/pp.109.149013
- James J. Clarkson u. a.: Phylogenetic relationships in Nicotiana (Solanaceae) inferred from multiple plastid DNA regions. In: Molecular Phylogenetics and Evolution. Band 33, 2004, S. 75–90. doi:10.1016/j.ympev.2004.05.002
- Sandra Knapp, Mark W. Chase, James J. Clarkson: Nomenclatural changes and a new sectional classification in Nicotiana (Solanaceae). In: Taxon. Band 53, Nummer 1, Februar 2004, S. 73–82.
- K. Yoong Lim u. a.: Sequence of events leading to near-complete genome turnover in allopolyploid Nicotiana within five million years. In: New Phytologist. Band 175, Ausgabe 4, 2007. doi:10.1111/j.1469-8137.2007.02121.x
- Ales Kovarik u. a.: Evolution of rDNA in Nicotiana Allopolyploids: A Potential Link between rDNA Homogenization and Epigenetics. In: Annals of Botany. Band 101, Ausgabe 6, 2008, S. 815–823. doi:10.1093/aob/mcn019
- S. Anssour u. a.: Phenotypic, genetic and genomic consequences of natural and synthetic polyploidization of Nicotiana attenuata and Nicotiana obtusifolia. In: Annals of Botany. Band 103, 2009, S. 1207–1217. doi:10.1093/aob/mcp058
- Tamara Krügel: Nicotiana ×mierata und N. ×obtusiata – zwei neue synthetische Hybriden. In: Haussknechtia. Mitteilungen der Thüringischen Botanischen Gesellschaft Jena. Heft 12, 2010, S. 23–26.
- William Albert Setchell: Aboriginal Tobaccos. In: The American Anthropologist. Band 23, Nummer 4, 1921, S. 397–414. doi:10.5962/bhl.title.19301
- Forrest L. Gager Jr., Virginia Johnson, Joseph C. Holmes: Ancient Tobacco Smokers. In: Science. Band 132, Nummer 3433, Oktober 1960, S. 1021. doi:10.1126/science.132.3433.1021
- Robert F. Raffauf: Persistence of Alkaloids in Plant Tissue. In: Science. Band 131, Nummer 3406, April 1960, S. 1047. doi:10.1126/science.131.3406.1047
- Nicotiana attenuata. In: Native American Ethnobotany – A Database of Foods, Drugs, Dyes and Fibers of Native American Peoples, Derived from Plants. University of Michigan-Dearborn. Online, abgerufen am 29. September 2012
- Sereno Watson: United States Geological Exploration of the Fortieth Parallel, Volume 5: Botany, 1871, S. 276. doi:10.5962/bhl.title.414
- Germplasm Resources Information Network (GRIN): Nicotiana attenuata. Abgerufen am 1. September 2012.
- Thomas C. Porter, John M. Coulter: Synopsis of the Flora of Colorado, United States Government Printing Office, Washington 1874, S. 110–111. doi:10.5962/bhl.title.46971
- Asa Gray: Synoptical Flora of North America. Band 2, Teil 1, Ivison, Blakeman, Tayler and Company, New York, Mai 1878. doi:10.5962/bhl.title.10847
- Volney Rattan: A popular California Flora. A.L. Bancroft and Company, San Francisco, 1882, S. 88a. doi:10.5962/bhl.title.7729
- Willis Linn Jepson: A Flora of western middle California. Encina Publishing Company, Berkeley 1901, S. 391. doi:10.5962/bhl.title.59931
- Per Axel Rydberg: Flora of Colorado. The Agricultural Experiment Station of the Colorado Agricultural College, 1905, S. 304. doi:10.5962/bhl.title.28827
- Theodore C. Frye, George B. Rigg: Elemental Flora of the Northwest. American Book Company, New York, Cincinnati, Chicago 1914, S. 198. doi:10.5962/bhl.title.24439
- Charles V. Piper, R. Kent Beattie: Flora of southeastern Washington and adjacent Idaho. New Era Printing Company, Lancaster 1914, S. 219. doi:10.5962/bhl.title.2502
- E. O. Wooton, Paul C. Standley: Flora of New Mexico. In: Contributions from the United States National Herbarium. Band 19, 1915. doi:10.5962/bhl.title.21099
- Joseph Kaye Henry: Flora of southern British Columbia and Vancouver Island. W.J. Gage & Co., Toronto 1915, S. 261. doi:10.5962/bhl.title.37900
- Per Axel Rydberg: Flora of the Rocky Mountains and adjacent plains. New York 1917, S. 760. doi:10.5962/bhl.title.28832
- Willis Linn Jepson: A Flora of California. Band 3, Curtis & Welch, Cunningham 1943, S. 453. doi:10.5962/bhl.title.7604
- Richard Olmstead, Jeffery D. Palmer: Chloroplast DNA and systematics of the Solanaceae. In: J. G. Hawkes, R. N. Lester, M. Nee, and N. Estrada (Hrsg.): Solanaceae 3: Taxonomy – Chemistry – Evolution. Royal Botanic Gardens, Richmond, U.K., 1991, S. 161–168.
- Vicente F. Garcia, Richard G. Olmstead: Phylogenetics of Tribe Anthocercideae (Solanaceae) Based on ndhF and trnL/F Sequence Data. In: Systematic Botany. Band 28, Nummer 3, 2003, S. 609–615. doi:10.1043/02-52.1
- S. Aoki, M. Ito: Molecular Phylogeny of Nicotiana (Solanaceae) Based on the Nucleotide Sequence of the matK Gene. In: Plant Biology. Band 2, Nummer 3, 2000, S. 316–324. doi:10.1055/s-2000-3710
- Richard Olmstead, Lynn Bohs: A Summary of Molecular Systematic Research in Solanaceae: 1982–2006. In: Solanaceae VI: Genomics Meets Biodiversity. Proceedings of the VIth International Solanaceae Conference, 2006. Acta Horticulturae 745, Juni 2007.
- Alison Abbott: Growth Industry. In: Nature. Band 468, 16. Dezember 2010, S. 886–888. doi:10.1038/468886a