Natrium-Kalium-Pumpe
Die Natrium-Kalium-ATPase (genauer: Na+/K+-ATPase), auch als Natrium-Kalium-Pumpe oder Natriumpumpe bezeichnet, ist ein in der Zellmembran verankertes Transmembranprotein. Das Enzym katalysiert unter Hydrolyse von ATP (ATPase) den Transport von Natrium-Ionen aus der Zelle und den Transport von Kalium-Ionen in die Zelle gegen die elektrochemischen Gradienten und dient so ihrer Aufrechterhaltung.
| Na+/K+-ATPase | ||
|---|---|---|
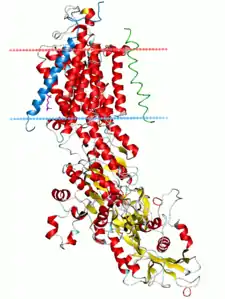 | ||
| Bändermodell der Na+/K+-ATPase nach 3B8E | ||
| Masse/Länge Primärstruktur | 1018+303+66 Aminosäuren | |
| Sekundär- bis Quartärstruktur | α+β+γ, multipass Membranprotein | |
| Kofaktor | Mg2+ | |
| Isoformen | α1/2, β1/2, γ1/2 | |
| Bezeichner | ||
| Gen-Name(n) | ATP1A1, ATP1A2, ATP1A3, ATP1A4 | |
| Transporter-Klassifikation | ||
| TCDB | 3.A.3.1.1 | |
| Bezeichnung | P-ATPase | |
| Enzymklassifikation | ||
| EC, Kategorie | 7.2.2.13, Hydrolase | |
| Reaktionsart | Hydrolyse | |
| Substrat | ATP + H2O + 3 Na+innen + 2 K+außen | |
| Produkte | ADP + Phosphat + 3 Na+außen + 2 K+innen | |
| Vorkommen | ||
| Übergeordnetes Taxon | Lebewesen | |
Geschichte
Die Na+/K+-ATPase wurde 1957 vom dänischen Mediziner Jens Christian Skou entdeckt.[1] 1997 erhielt Skou den Nobelpreis für Chemie „für die Entdeckung des ionentransportierenden Enzyms Natrium-Kalium-ATPase“.[2]
Struktur des Proteins
Nach der TCDB-Klassifikation für Membrantransport-Proteine zählt die Natrium-Kalium-Pumpe zu Familie der P-Typ-ATPasen (Typ 2).[3]
Die Natrium-Kalium-ATPase besteht aus je einer α- und einer β-Protein-Untereinheit, die zusammen ein stabiles und funktionsfähiges Enzym bilden. Die α-Untereinheit besteht aus 10 Transmembrandurchgängen, die durch intra- und extrazelluläre Loops verbunden sind. Sie hydrolysiert während des Reaktionszykluses ATP, wird im Bereich des großen cytoplasmatischen M4-M5-Loops phosphoryliert und transportiert die Natrium- und Kalium-Ionen. Deshalb wird sie auch als die katalytische Einheit bezeichnet. Von der α-Untereinheit sind bisher vier Isoformen bekannt: α1 – α4.[4]
Die β-Untereinheit ist ein Typ-II-Glycoprotein mit einem einzelnen Membrandurchgang. Im funktionsfähigen Enzym liegt sie in der Nähe der Transmembrandurchgänge M7 und M10 der α-Untereinheit und interagiert mit deren extrazellulären M7/M8-Loop M7/M8 sowie mit ihren intrazellulären Regionen. Während der Reifung des Proteins erfüllt die β-Untereinheit die Funktion eines molekularen Chaperons, indem sie die korrekte Faltung und Membranintegration der α-Untereinheit unterstützt und sie vor Degradation schützt. Außerdem hat sie Einfluss auf die Affinität, mit der die α-Untereinheit Kalium- und Natriumionen bindet. Von der β-Untereinheit sind bisher vier Isoformen bekannt: β1, β2, β3 und βm.[5]
Ein drittes Molekül, das sich mit der α- und der β-Untereinheit assoziiert und früher als γ-Untereinheit bezeichnet wurde, gehört zur Protein-Familie FXYD und moduliert die Affinität der Natriumpumpe für Natrium- und Kaliumionen sowie ATP. Es spielt außerdem eine Rolle bei der Stabilisation des Enzyms.[6] Für die Ausbildung des funktionsfähigen Enzyms ist die Anwesenheit eine γ-Untereinheit nicht notwendig.[7]
Die Natrium-Kalium-ATPase kommt in mehreren Kombinationen von Isoformen der α- und β-Untereinheiten vor, die Unterschiede in Verteilung, Affinität zu Herzglykosiden und Funktion aufweisen. Der α1-Isoenzymtyp findet sich in allen Zellen des Menschen, α2- und α3-Typen in den Nervenzellen und Herzmuskelzellen (Myokard). Seltene Mutationen in den ATP1A2- und ATP1A3-Genen können zu erblicher Migräne, alternierender Hemiplegie und Dystonie führen.[8][9][10]
Funktion
Der gegenläufige Transport (Antiport) von 3 Na+ gegen 2 K+ über die Zellmembran erfolgt gegen die jeweiligen Konzentrationsgefälle und in der Summe gegen das elektrische Ruhemembranpotential (elektrogen); er ist damit gleich zweifach von extern zugeführter Energie abhängig: energieabhängiger / aktiver Transport. Jene wird in diesem Fall als chemische Bindungsenergie durch die Hydrolyse von ATP zur Verfügung gestellt.
Das Phänomen des ATP-getriebenen Transports ist für den Na+/K+-Transport durch die Plasmamembran am besten untersucht. Beide Kationen sind in Zellen ungleich verteilt:
- Die Na+-Konzentration im Inneren der Zelle ist gering (7–11 mmol/l);
- die K+-Konzentration im Inneren ist hoch (120–150 mmol/l).
Dieser lebenswichtige Konzentrationsgradient wird einerseits durch Kalium-Kanäle bewirkt (siehe Blutzucker-Sensorsystem), andererseits durch die elektrogene Natrium-Kalium-ATPase.
Mechanismus

ATP-Hydrolyse und Na+/K+-Transport sind strikt gekoppelt (elektrogenes Prinzip):
- Je Molekül ATP werden drei Na+-Ionen nach außen und zwei K+-Ionen nach innen befördert. In der Bilanz wird dem intrazellulären Raum also ein positiver Ladungsträger entzogen. Dieser Mechanismus ist die treibende Kraft für den Erhalt des insbesondere für Nerven- und Muskelzellen funktional wichtigen elektrischen Ruhemembranpotentials. Es ist jedoch nicht die Ladungsbilanz der Pumpe allein, die zur Erzeugung des Ruhemembranpotentials führt. Die hohe Permeabilität der Zellmembran für K+-Ionen, die durch Kalium-Kanäle gegeben ist und die geringe Permeabilität für Na+-Ionen während des Ruhemembranpotentials sind ebenfalls für dessen Erhalt erforderlich.
- für den Pumpmechanismus sind Konformationsänderungen erforderlich, die durch Phosphorylierung eines Aspartat (Asp-) Restes der α-Untereinheit erzeugt werden.
- bei diesem Vorgang werden zunächst drei Na+-Ionen eingeschlossen;
- deren Abgabe nach außen erfolgt im Austausch gegen zwei K+-Ionen, deren Bindung eine Phosphatase aktiviert, die den Asp-Rest wieder dephosphoryliert.
Wirkung der Herzglykoside
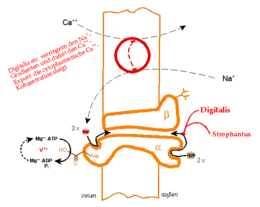
Glykoside der Digitalis-Gruppe (Digoxin, Digitoxin und dessen Aglykon Digitoxigenin) und der Gruppe der Strophanthusgewächse (g-Strophanthin (engl. Synonym: Ouabain) und k-Strophanthin) – letztere jedoch nur in hohen Konzentrationen – blockieren die K+-Konformation der ATPase noch im phosphorylierten Zustand. Damit hemmen sie den Ionentransport.
Indirekt steigt hierdurch die Konzentration an intrazellulärem Ca2+ und damit die Kontraktion des Herzmuskels, denn
- der Transport des Ca2+ hängt nach dem Antiport-Prinzip vom Na+-Konzentrationsgradienten ab (Natrium-Calcium-Austauscher).
- Ist dieser verringert, verbleibt Ca2+ in zunehmendem Maße in der Muskelzelle, wobei sich deren Kontraktion verstärkt.
Da die Herzmuskelzellen eines Menschen mit Herzinsuffizienz zu viel Calcium enthalten (calcium overload, der zur Minderung der Kontraktilität führt), war es bis vor kurzem unverständlich, wieso eine weitere Steigerung des zellulären Calcium-Gehalts zur Steigerung der Kontraktilität führen kann. Eine mögliche erklärende Hypothese: Die α2- und α3-Isoformen der Natrium-Kalium-Pumpen sind zusammen mit den Natrium-Calcium-Austauschern direkt über den Ausläufern des Calcium-Speichers der Zelle (Sarkoplasmatisches Retikulum) lokalisiert. Diese funktionelle Einheit wird Plasmerosom genannt. Hierdurch kann die lokale Natrium- bzw. Calcium-Konzentration durch Hemmung nur relativ weniger Natrium-Kalium-Pumpen durch Herzglykoside gesteigert werden, was das Sarkoplasmatische Retikulum zur Freisetzung von wesentlich größeren Mengen an Calcium an die kontraktilen Proteine (bei zum Beispiel jedem Herzschlag) anregt, ohne dass sich die Gesamt-Konzentration der Zelle an Natrium- und Calcium wesentlich verändert. Diese wird eher durch die α1-Isoform der Natrium-Kalium-Pumpe reguliert. Die Plasmerosome wurden für Nervenzellen und Arterien-Muskelzellen bereits nachgewiesen und sind wahrscheinlich auch in Skelett- und Herzmuskelzellen vorhanden.[11]
g-Strophanthin ist ein endogenes Herzglykosid, das bei Säugetieren von der Nebennierenrinde produziert wird. Die endogene Synthese, eine orale Einnahme oder eine niedrig dosierte, langsame intravenöse Verabreichung von g-Strophantin führen nur zu geringen Plasmakonzentrationen, die zunächst zu einer Stimulation der Natrium-Kalium-Pumpe mit daraus resultierender Reduzierung des zellulären Natrium- und Calcium-Gehalts führen. Dies kann zu einem negativ inotropen Effekt wie bei einem Nitro-Präparat oder auch zu einem positiv inotropen Effekt führen (wahrscheinlich je nach Ausgangslage der Calcium-Konzentration der Herzmuskelzellen).[12][13][14]
Auch k-Strophanthin kann die Na-K-Pumpe stimulieren, jedoch nicht Digoxin.[13] Dies erklärt zum Beispiel die gegensätzliche Wirkung von Strophanthin und Digoxin bei Angina Pectoris, wobei Strophanthin positiv auf EKG und Anfallshäufigkeit wirkt, Digoxin bekanntlich jedoch negativ.[13][15]
Andere Substanzen mit Einfluss auf den Transporter
Eine Hemmung der Natrium-Kalium-ATPase kann außer durch Digitalis ebenfalls durch Ouabain stattfinden.
Umgekehrt erreicht man eine Aktivierung des Transporters durch den Einfluss von Insulin oder Adrenalin. Ersteres wird häufig im klinischen Alltag dazu genutzt, um bei einer Hyperkaliämie durch Gabe von Insulin zeitgleich mit Glucose einen Shift des extrazellulären Kaliums nach intrazellulär über Steigerung der Aktivität der Natrium-Kalium-ATPase zu erreichen. Dieser Effekt hilft bei Hyperkaliämie aber nur kurzfristig, da das Kalium nur nach intrazellulär verlagert wird und nach einer gewissen Zeit wieder ins Blut gelangt.
Siehe auch
Weblinks
Einzelnachweise
- Skou, J.C. (1957): The influence of some cations on an adenosine triphosphatase from peripheral nerves. In: Biochim. Biophys. Acta. Bd. 23, S. 394–401. PMID 13412736
- Informationen der Nobelstiftung zur Preisverleihung 1997 an Jens Christian Skou (englisch)
- Eintrag der Familie der P-type ATPase auf der Homepage der TCDB. Abgerufen am 4. Januar 2014.
- G. Blanco: Na,K-ATPase subunit heterogeneity as a mechanism for tissuespecific ion regulation. In: Seminars in Nephrology. 2005, S. 292–303.
- N. Pestov, N. Ahmad, T. Korneenko, et al.: Evolution of Na,K-ATPase βm-subunit into a coregulator of transcription in placental mammals. In: Proceedings of the National Academy of Sciences. 2007, S. 11215–11220.
- D. Jones, T. Li, E. Arystarkhova et al.: Na,K-ATPase from mice lacking the γ-subunit (FXYD2) exhibits altered Naþ affinity and decreased thermal stability. In: Journal of Biological Chemistry. 2005 S. 19003–19011.
- G. Scheiner-Bobis, R. Farley: Subunit requirements for expression of functional sodium pumps in yeast cells. In: Biochimica et Biophysica Acta. 1994, Aug 3;1193(2), S. 226–234.
- Orphanet: Hemiplegische Migräne
- Orphanet: Alternierende Hemiplegie der Kindheit
- Orphanet: Dystonie-Parkinsonismus
- Blaustein MP, Juhaszova M, Golovina VA, Church PJ, Stanley EF: Na/Ca exchanger and PMCA localization in neurons and astrocytes: functional implications. In: Ann. N. Y. Acad. Sci.. 976, November 2002, S. 356–66. PMID 12502582.
- Gao J, Wymore RS, Wang Y, et al.: Isoform-specific stimulation of cardiac Na/K pumps by nanomolar concentrations of glycosides. In: J. Gen. Physiol.. 119, Nr. 4, April 2002, S. 297–312. PMID 11929882. PMC 2238186 (freier Volltext).
- Saunders R, Scheiner-Bobis G: Ouabain stimulates endothelin release and expression in human endothelial cells without inhibiting the sodium pump. In: Eur. J. Biochem.. 271, Nr. 5, März 2004, S. 1054–62. PMID 15009217.
- Belz GG, Matthews J, Sauer U, Stern H, Schneider B: Pharmacodynamic effects of ouabain following single sublingual and intravenous doses in normal subjects. In: Eur. J. Clin. Pharmacol.. 26, Nr. 3, 1984, S. 287–92. PMID 6428911.
- Kubicek F, Reisner T: Hypoxietoleranz bei koronarer Herzkrankheit unter der Einwirkung von Digoxin, Beta-Methyl-Digoxin und g-Strophanthin. In: Therapie der Gegenwart. 112, Nr. 5, Mai 1973, S. 747–9 passim. PMID 4708582.